Article Text

Abstract
Background and aims HCV infection is a leading risk factor of hepatocellular carcinoma (HCC). However, even after viral clearance, HCC risk remains elevated. HCV perturbs host cell signalling to maintain infection, and derailed signalling circuitry is a key driver of carcinogenesis. Since protein phosphatases are regulators of signalling events, we aimed to identify phosphatases that respond to HCV infection with relevance for hepatocarcinogenesis.
Methods We assessed mRNA and microRNA (miRNA) expression profiles in primary human hepatocytes, liver biopsies and resections of patients with HCC, and analysed microarray and RNA-seq data from paired liver biopsies of patients with HCC. We revealed changes in transcriptional networks through gene set enrichment analysis and correlated phosphatase expression levels to patient survival and tumour recurrence.
Results We demonstrate that tumour suppressor protein tyrosine phosphatase receptor delta (PTPRD) is impaired by HCV infection in vivo and in HCC lesions of paired liver biopsies independent from tissue inflammation or fibrosis. In liver tissue adjacent to tumour, high PTPRD levels are associated with a dampened transcriptional activity of STAT3, an increase of patient survival from HCC and reduced tumour recurrence after surgical resection. We identified miR-135a-5p as a mechanistic regulator of hepatic PTPRD expression in patients with HCV.
Conclusions We previously demonstrated that STAT3 is required for HCV infection. We conclude that HCV promotes a STAT3 transcriptional programme in the liver of patients by suppressing its regulator PTPRD via upregulation of miR-135a-5p. Our results show the existence of a perturbed PTPRD–STAT3 axis potentially driving malignant progression of HCV-associated liver disease.
- HEPATOCELLULAR CARCINOMA
- HCV
- SIGNALING
This is an Open Access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/
Statistics from Altmetric.com
Significance of this study
What is already known on this subject?
Chronic HCV infection is a leading cause of hepatocellular carcinoma (HCC); HCC risk remains elevated even after viral clearance.
The mechanisms contributing to HCC development are poorly understood.
HCV requires host cell signalling including the STAT3 pathway to maintain infection.
Protein tyrosine phosphatase receptor delta (PTPRD) suppresses tumour by controlling STAT3 activity.
What are the new findings?
PTPRD expression is impaired in hepatocytes of HCV-infected liver tissues and in HCC lesions.
PTPRD expression in liver tissue of patients with HCC is associated with STAT3 transcriptional activity, patient survival and tumour recurrence after surgical resection.
STAT3-mediated transcriptional programme is enriched in HCV-infected livers.
miR-135a-5p is strongly upregulated in livers of patients with HCV and is a regulator of PTPRD mRNA.
How might it impact on clinical practice in the foreseeable future?
HCV infection promotes the activity of its cofactor STAT3 by suppressing its negative regulator PTPRD via miR-135a-5p. Our model suggests the existence of a perturbed PTPRD–STAT3 axis driving malignant progression of liver disease. This finding is of further clinical relevance since it provides a target for urgently needed HCC chemoprevention.
Introduction
More than 150 million people worldwide are infected by HCV,1 which is a leading cause of liver cirrhosis and hepatocellular carcinoma (HCC).2 The arrival of highly effective new therapies consisting of direct-acting antivirals can cure the vast majority of patients,3 but those with advanced liver disease remain at risk for developing HCC even after viral clearance.4 Moreover, the exact mechanisms responsible for this increased susceptibility remain only partly understood. Traditionally, carcinogenesis has been attributed to a multistep accumulation of genetic damage resulting in gain of function by proto-oncogenes and inactivation of tumour suppressor genes.5 Only a minority of these driver genes is frequently involved in cell transformation, while the majority is only occasionally affected.6 In other words, a myriad of combinations in genetic and epigenetic alterations can lead to cancer, and as a consequence, every tumour displays a heterogeneous molecular profile, further clouding understanding of cancer development. In chronic hepatotropic virus infections such as hepatitis C and B, the picture is even more complicated as both viral and non-viral factors are the drivers of hepatocarcinogenesis.7 ,8 Nonetheless, the complexity of malignant transformation can be reduced to a handful of logical underlying principles, of which derailed signalling circuitry is a key component.9 We have previously demonstrated that cellular receptor tyrosine kinase (RTK) signalling is involved in regulating HCV entry into hepatocytes and that the virus binding to the hepatocytes triggers RTK activation.10 ,11 ,12 This indicates that chronic HCV infection modulates host signalling patterns that may contribute to the development of virus-induced liver disease. Since these signalling processes are tightly regulated by protein phosphatases and aberrant phosphatase expression is involved in various syndromes and diseases,13 we screened for disease-relevant phosphatase expression in liver biopsies of chronic patients with HCV.
Materials and methods
Liver tissue, cells and virus
Human needle liver biopsies and liver tissue from patients undergoing surgical resection were collected at the Gastroenterology and Hepatology outpatient clinic of the Basel University Hospital, Switzerland, the Centre Hospitalier Universitaire de Reims, France and the Hôpitaux Universitaires de Strasbourg, France. Protocols for patient tissue collection were reviewed and approved by the ethics committees of the respective university hospitals. Written informed consent was obtained from all patients. Eligible patients were identified by a systematic review of patient charts at the hepatology centres of the university hospitals of Basel, Reims and Strasbourg. Histopathological grading and staging of the HCV liver biopsies, according to the METAVIR classification system, were performed at the pathology institutes of the respective university hospitals. All the patients that donated liver tissue are summarised in online supplementary table S1. Liver biopsy tissues were analysed as described.14 Mouse tissue was obtained from C57BL/6J mice. Tissues were lysed and subjected to quantitative PCR (qPCR) and immunoblot analysis (described below). Primary human hepatocytes (PHH) were isolated and cultured as previously described.11 Huh7.5.1, HEK293T cells and cell culture derived HCV (HCVcc) strains Luc-Jc1 and Jc1E2FLAG have been described.11 ,12 ,15 Jc1E2FLAG was affinity-purified as described.15 Huh7.5.1 were infected with Jc1E2FLAG as described.16
supplementary data
Patient cohorts
Protein tyrosine phosphatase receptor delta (PTPRD) expression was correlated with corresponding clinical data in three independent patient cohorts from Icahn School of Medicine at Mount Sinai, New York, USA (National Centre for Biotechnology Information Gene Expression Omnibus GSE10140) (cohort A),17 from the Hiroshima University Hospital, Hiroshima, Japan (European Genome-phenome Archive, https://ega.crg.eu, accession number, EGAD00001001880) (cohort B)18 and from the University Hospitals of Strasbourg and Reims, France (cohort C).
Antibodies and western blotting
P-STAT1 mAb (58D6) was obtained from Cell Signaling Technology, β-Actin mAb (AC-15) was obtained from Invitrogen and PTPRD pAb (C-18) from Santa Cruz Biotechnology. Western blots were performed using Hybond-P membranes (GE Healthcare), visualised using ECF substrate (GE Healthcare) and quantified with a fluorescence scanner (Typhoon Trio, GE Healthcare).
Analysis of mRNA and miRNA expression
mRNA expression of 84 disease-relevant protein phosphatases was assessed in six liver biopsies from patients with chronic HCV infection and in six non-infected biopsies using qPCR (Human Protein Phosphatases RT² Profiler PCR Array, Qiagen). Total RNA from Huh7.5.1 cells, PHHs and liver tissue was extracted using RNeasy Mini Kit (Qiagen) or Trizol (Life Technologies). Gene expression in the total RNA extracts was assessed using two-step qPCR. The reverse transcription on total RNA extract was made using Maxima reverse transcriptase (Thermo Scientific). qPCR for detecting PTPRD, radical S-adenosyl methionine domain containing 2 (RSAD2), ubiquitin-specific peptidase 18 (USP18) and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was performed with RT² qPCR Primer Assays (Qiagen) using Real-Time PCR ABI Prism 7500 (Thermo Scientific) or Corbett Rotor Gene 6000 (Qiagen). All qPCRs were performed following manufacturers' instructions. Specificities of phosphatase PCR products were validated by melting curve analysis and PCR product sequencing. Differential gene expression of patient biopsies was calculated after 2−ΔCT transformation into individual data points. The transcriptome of Jc1E2FLAG-infected Huh7.5.1 cells was measured by RNA-seq at the Broad Institute as previously described.19 All microRNA (miRNA) indicated in this study are human (hsa-miR) if not indicated differently. miRNA expression was measured using miScript (Qiagen) with forward DNA primers derived from the miRNA sequence implemented in Sanger miRBase database (V.21.0) and referenced with a MIMAT accession number (see online supplementary table S2). miRNA target sites were predicted using miRSystem.20 Synthetic miRNAs (mimics) of miR-135a-5p and the non-targeting negative control miRNA cel-miR-67 (miR-CTRL) were obtained from GE Healthcare.
Luciferase reporter assay
The 3′-untranslated region (3′UTR) of PTPRD was amplified with 5′-TTT CTC GAG CTT TGA CCA CTA TGC AAC GTA G-3′ and 5′-TTT CTC GAG CTG TCC TCG CCG TTT TCT AA-3′, and subcloned in psiCheck-2 (Promega) using XhoI. As specificity control, the seed sequence AGCCAT of two miR-135a-5p target sites on the 3′UTR of PTPRD was replaced by AAAAAA using site-directed mutagenesis. An amount of 150 ng luciferase reporter plasmid was cotransfected with 10 nmol/L miRNA in HEK293T cells using Lipofectamine 2000 (Thermo Fisher Scientific). Activity of Renilla and firefly luciferase was assessed 48 hours post transfection using Dual-Luciferase Reporter Assay (Promega) and a Mithras LB940 plate reader (Berthold Technologies).
FISH analysis
Human needle liver biopsies were collected, immediately embedded in optimal cutting temperature compound and frozen in liquid nitrogen chilled 2-methylbutane. Tissues were then stored at −80°C until use. Sections (10 µm) were cut at cryostat (Leica) and mounted onto Superfrost Plus Gold glass slides (K5800AMNZ72, Thermo Fisher Scientific), fixed overnight in 4% formaldehyde at 4°C and hybridised, as previously described,21 with the following modifications: tissue sections were pretreated by boiling (90°C–95°C) in pretreatment solution (Panomics, Affymetrix) for 1 min, followed by a protease digestion for 10 min at 40°C. Hybridisation was performed using probe sets against patient-specific HCV RNA sequence (type 1) and against human PTPRD mRNA (NM_002839, target region 4754-5835). Preamplification, amplification and detection were performed according to provider's protocol. Images were acquired with a laser scanning confocal microscope (LSM710, Carl Zeiss Microscopy) and Zen2 software, using the same settings for all the tissues analysed. Five random fields were acquired from each section. Image analysis was performed using ImageJ and CellProfiler software, with a customised pipeline. Total number of cells, frequency of HCV-positive and PTPRD-positive cells, and signal intensity were then evaluated and exported for statistical analysis (two-way analysis of variance (ANOVA), p≤0.05).
Bioinformatics of gene expression database
Paired gene expression of 62 formalin-fixed paraffin-embedded tumour and adjacent liver tissues was explored from cohort A17). Downregulation of PTPRD expression in tumour tissue was analysed in silico for 46 patients with chronic HCV. Values of p were calculated with Wilcoxon signed-rank test (two-tailed). Probability of survival and tumour recurrence were evaluated by the log-rank test available on the GenePattern Survival Analysis module (http://www.broad.mit.edu/cancer/software/genepattern).22 Gene Set Enrichment Analysis (GSEA) was performed as described.23 ,24 The normalised enrichment score (NES) is a measure expressing to what extent the members of a gene set shift towards the top or bottom in the gene list ranked on differences in expression between two phenotypes. A false discovery rate (FDR) smaller than 0.25 is generally considered an appropriate cut-off. The GSE10143 database comprised 78 of the 87 response genes designated as the Hallmark_IL6 JAK STAT3 signalling gene set.25
Results
Chronic HCV infection impairs PTPRD expression in vivo
To study the impact of chronic HCV infection on phosphatase expression in patient liver tissue, we extracted RNA from six biopsies of patients with chronic hepatitis C (CHC) and six non-infected biopsies (see online supplementary table S1), and quantified expression of 84 disease-relevant protein phosphatases using qPCR. We identified 24 phosphatases that were significantly (p<0.01, U-test) deregulated in HCV-infected tissue compared with non-infected biopsies (figure 1A). Interestingly, among the phosphatases with most deregulated expression levels in HCV biopsies, we observed an enrichment of candidates with potential relevance for the development of HCC. More particularly, some of these phosphatases act as tumour suppressors in various cancers including PTPRD. PTPRD is frequently inactivated and mutated in human cancers26 ,27 including HCC.28 In order to validate HCV-induced PTPRD downregulation, we measured mRNA expression in a second sample series of 24 liver biopsies of patients with chronic HCV and validated that PTPRD mRNA expression is significantly (p=0.0003, U-test) downregulated compared with 11 additional non-viral control biopsies (figure 1B).

Protein tyrosine phosphatase receptor delta (PTPRD) expression is significantly impaired in livers of patients with chronic hepatitis C (CHC). Expression of 84 protein phosphatases associated with diseases and syndromes was quantified in needle stick biopsies using qPCR and in situ hybridisation of RNA probes. (A) mRNA expression of 11 phosphatases was significantly (p<0.01, U-test) increased (red), and expression of 13 phosphatases was significantly (p<0.01, U-test) decreased (blue). Data are expressed as fold change (fc) phosphatase expression of non-HCV (n=6) and HCV-infected biopsies (n=6) relative to the average expression levels of five housekeeping genes (actin, B2M, GAPDH, HPRT1, RPLP0). Arrow indicates tumour suppressor PTPRD. (B) Expression of PTPRD is significantly impaired in liver biopsies of patients with chronic HCV (p=0.0003, U-test). Validation of HCV-induced downregulation of PTPRD compared with non-infected biopsies (NI) observed in (A) in additional liver biopsies from patients infected with HCV (see online supplementary table S1). Data are expressed as PTPRD mRNA expression relative to GAPDH and visualised as individual data points and median (line). (C and D) Expression of PTPRD is independent from fibrosis and inflammation in liver tissues. PTPRD expression was correlated with METAVIR score of the HCV-infected liver biopsies, but no significant correlation (p>0.5, U-test) between METAVIR grading (C) or staging (D) of the studied biopsies in (A) and (B) (see online supplementary table S1) was observed. Data are expressed as PTPRD mRNA expression relative to GAPDH and visualised as individual data points and median (line). (E) PTPRD expression is impaired in HCV-infected hepatocytes in liver biopsies. Fluorescent in situ hybridisation (FISH) analysis of liver biopsies infected with HCV genotype 3 by simultaneous hybridisation with HCV-specific and PTPRD-specific probe sets. PTPRD labelling was assessed in five random fields from each section. Data are expressed as mean of PTPRD fluorescence intensities in HCV-positive and HCV-negative cells per patient (±SD, p<0.05, two-way analysis of variance).
PTPRD expression in patients with HCV is independent from fibrosis or inflammation
Since biological circuits in liver tissues from patients with CHC reflect a complex interaction of different cell types integrating an inflammatory immune response, we first compared phosphatase expression levels with METAVIR grading and staging data (see online supplementary table S1). No significant relationship between METAVIR score and PTPRD expression was observed in 30 liver biopsies (figure 1C, D), suggesting that PTPRD expression is independent from the degree of inflammation and fibrosis of the studied biopsies and rather susceptible to a more direct virus-induced mechanism within infected hepatocytes.
PTPRD expression is impaired in HCV-infected hepatocytes
To identify the cell type displaying impaired PTPRD expression levels upon HCV infection, we applied an established protocol for multiplex fluorescent in situ hybridisation (FISH) analysis of liver biopsies at single cell resolution.21 This strategy enabled us to distinguish HCV-infected cells from uninfected cells in liver biopsies. Simultaneous hybridisation with specific probe set for PTPRD mRNA demonstrated that PTPRD was significantly and specifically (p<0.05, two-way ANOVA) impaired in infected cells compared with uninfected cells in three biopsies from different patients infected with HCV genotype 3a (see figure 1E, online supplementary table S1 and figure S1). Differential PTPRD expression was detected in cells with hepatocyte morphology. The analysed fields did not show an abnormal level of infiltrating non-parenchymal cells, and the level of PTPRD expression in those cells was marginal compared with hepatocytes (data not shown). This indicates that HCV impairs PTPRD expression directly in hepatocytes. To validate this observation, we measured PTPRD expression levels in isolated PHH infected in vitro with HCV. We observed that PHH robustly express PTPRD both at the mRNA and the protein level, providing a competent model system to study PTPRD. Two days after isolation, PHH were infected for 5 days with HCVcc strain JFH1, after which cells were lysed, and both mRNA and protein were isolated. With this established protocol, up to 10%–30% of hepatocytes can be infected by HCVcc (strain JFH1) as shown29 (see online supplementary figure S2) using a reporter red fluorescence protein–nuclear localisation sequence–interferon-β promoter stimulator lentivirus (RFP-NLS-IPS).30 HCV-infected PHH from different donors showed a significant decrease (p=0.015, U-test) in PTPRD mRNA expression (figure 2A), which also translated into a decreased PTPRD protein level as detected by immunoblot (p=0.016, U-test) (figure 2B, C). To exclude that PTPRD downregulation was caused by an antiviral response mounted by immunocompetent PHH, we treated PHH for 5 days with interferon-α (IFN-α) (figure 2D). No change in PTPRD protein levels was observed by western blot for different concentrations of IFN-α, while STAT1 phosphorylation was properly induced. In parallel, interferon response genes USP18 and RSAD2 displayed an increase in mRNA transcription after IFN-α stimulus, while PTPRD remained at baseline levels (figure 2E). These data confirm that PTPRD downregulation is independent from the innate immune response.

Protein tyrosine phosphatase receptor delta (PTPRD) is significantly impaired by HCV in hepatocytes independent from the innate immune response. (A) PTPRD mRNA expression is significantly impaired in primary human hepatocytes (PHH) after 5 days of infection with HCVcc (strain JFH1). Data are expressed as mean PTPRD expression relative to GAPDH±SEM (p=0.015, U-test; six independent infections of PHH from three donors). (B) PTPRD protein expression is significantly impaired by HCV in PHH. PTPRD and actin expression were assessed by western blotting 5 days after infection of PHH with HCVcc (strain JFH1) compared with non-infected PHH (NI). Band intensities were quantified using Image Quant and are expressed as mean PTPRD expression relative to actin±SEM (p=0.016, U-test; five independent infections of PHH from two different donors). (C) A representative western blot quantified in (B) is shown. (D) PTPRD protein expression is independent from innate immune response. Host antiviral response was simulated by incubation of PHH with interferon-α (IFN-α). Cell culture medium with IFN-α was replaced every day. Cells were lysed after 5 days of incubation, and PTPRD, phospho-STAT1 (p-STAT1) and actin were measured by western blotting. (E) PTPRD is not an IFN-response gene. PHH were incubated for 10 min with 10³ IU/mL IFN-α, and mRNA expression of PTPRD and the known interferon-response genes USP18 and RSAD2 was assessed by RT-qPCR relative to GAPDH 6 and 16 hours post IFN stimulation. CTRL, control.
miR-135a-5p is a mechanistic regulator of PTPRD expression in HCV-infected hepatocytes
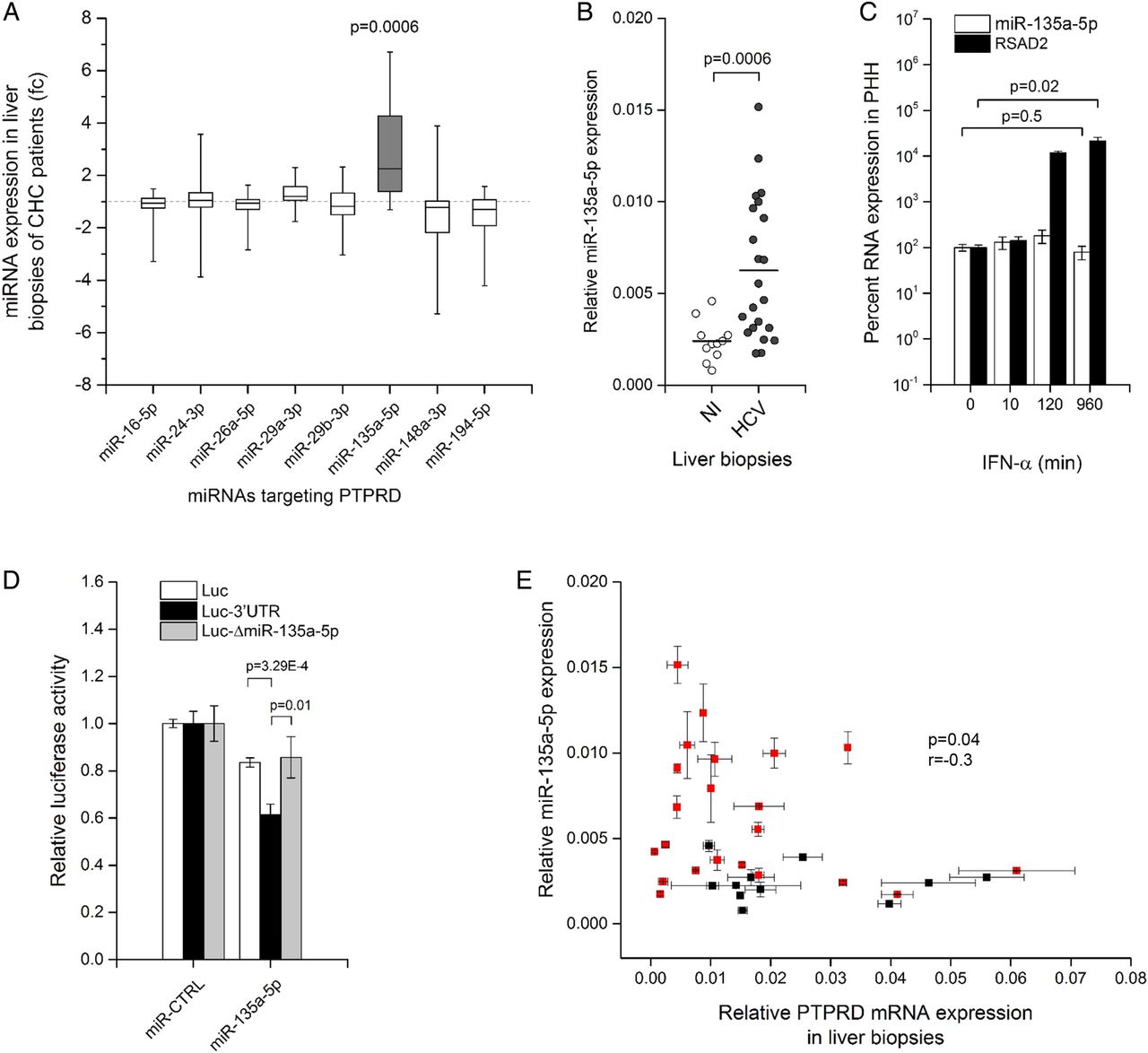
The miRNA machinery is a regulator of gene expression and is exploited by HCV to maintain its replication.31 ,32 We applied multiple bioinformatical algorithms20 scanning for human miRNAs potentially targeting PTPRD mRNA. Combining computational miRNA prediction with observations in HCV-infected Huh7.5.1 cells, we defined a panel of eight miRNAs with two essential characteristics: they potentially target PTPRD and they are upregulated by HCVcc in Huh7.5.1 cells (see online supplementary table S2). To validate the relevance of these eight miRNAs in a real-life setting, we screened for their expression in liver biopsies of patients infected with HCV (figure 3A). This uncovered a striking twofold upregulation of miR-135a-5p in liver specimens of patients with HCV (p=0.0006, U-test) and establishes it as a possible PTPRD regulator (figure 3B). Moreover, similar to PTPRD, miR-135a-5p expression is independent from the host antiviral response, as shown by a kinetics study measuring miR-135a-5p in IFN-α stimulated PHH. miR-135a-5p expression remained level (p=0.5, U-test), while the interferon response gene RSAD2 was significantly (p=0.02, U-test) upregulated (figure 3C). This suggests that miR-135a-5p does not take part in the innate immune response, but is rather relevant to the HCV life-cycle itself. To prove that miR-135a-5p targets PTPRD mRNA, we subcloned the 3′UTR of PTPRD mRNA in the Renilla luciferase expression cassette (Luc-3′UTR) of a bicistronic Renilla/firefly luciferase reporter construct (psiCheck-2, Promega). The 3′UTR of PTPRD harbours two predicted highly conserved miR-135a-5p target sites (see online supplementary table S2). Cotransfection of a miR-135a-5p mimic with the Luc-3′UTR reporter significantly (p=3.29×10−4, U-test) impaired normalised luciferase activity compared with empty vector, while the repression of luciferase expression was lost when the Luc-ΔmiR-135a-5p reporter with mutated miR-135a-5p binding sites was used (figure 3D). This demonstrates that miR-135a-5p is able to silence PTPRD expression, which is also reflected in vivo where a significant (p=0.04, r=−0.03, one-tailed Spearman's correlation test) inverse correlation of PTPRD mRNA and miR-135a-5p levels could be discerned in liver biopsies studied in figure 3B. Remarkably, high levels of miR-135a-5p always corresponded with low PTPRD expression, while low amounts of miR-135a-5p give PTPRD expression more leeway to vary. Interestingly, some of the studied liver biopsies did neither exhibit high PTPRD mRNA nor miR-135a-5p levels (figure 3E), suggesting the presence of an additional more general regulatory mechanism of PTPRD expression that is independent from HCV. Taken together, these data establish miR-135a-5p as a potent HCV-driven regulatory element for PTPRD expression.

HCV-induced expression of miR-135a-5p mediates protein tyrosine phosphatase receptor delta (PTPRD) silencing. (A) miRNA expression screening of potential regulators of PTPRD mRNA in livers of patients with chronic hepatitis C (CHC). Predicted miRNAs potentially regulating the PTPRD mRNA and that are upregulated by HCV in Huh7.5.1 cells (see online supplementary table S2) were screened in liver biopsies using qPCR. Data are expressed as fold change (fc) miRNA expression relative to SNORD61 and visualised as box-and-whisker plot (box=50% of biopsies, line=median, whiskers=minimal and maximal values) centred to the median expression levels of the miRNA in the corresponding non-infected (NI) liver biopsies. Grey bar highlights miR-135a-5p that is upregulated more than twofold in patients with HCV. (B) miR-135a-5p expression is significantly (p=0.0006, U-test) upregulated in livers of patients with HCV (dark grey circles) compared with NI tissues (open circles). (C) miR-135a-5p is not induced by the antiviral response to HCV infection. Isolated primary human hepatocytes (PHH) were treated over 16 hours with 103 IU/mL interferon-α (IFN-α) prior to qPCR analysis of miR-135a-5p and the interferon-stimulated gene RSAD2. While interferon-response gene RSAD2 is significantly (p=0.02, U-test) induced by IFN-α, miR-135a-5p remains level (p=0.5, U-test). (D) miR-135a-5p induces silencing of PTPRD expression. 3′UTR of PTPRD harbouring two miR-135a-5p target sites was subcloned in the Renilla luciferase expression cassette of a bicistronic Renilla/firefly luciferase reporter plasmid (Luc-3′UTR). As a control (Luc-ΔmiR-135a-5p), the seed sequence AGCCAT in Luc-3′UTR was replaced by AAAAAA. Cotransfection of Luc-3′UTR but not Luc-ΔmiR-135a-5p with a miR-135a-5p mimic significantly (p=3.29×10−4, U-test) impairs Renilla luciferase expression in HEK293T cells compared with empty vector (Luc). A minor unspecific effect of miR-135a-5p on empty vector (Luc) was observed. Data are expressed as mean Renilla luciferase activity±SEM normalised to firefly luciferase (five independent experiments in triplicate) relative to cotransfection of the vectors with non-targeting miRNA derived from Caenorhabditis elegans (miR-CTRL). (E) PTPRD mRNA levels significantly and inversely correlate (p=0.04, r=−0.03, one-tailed Spearman's correlation test) with miR-135a-5p levels in liver biopsies. PTPRD mRNA and miR-135a-5p expression in NI and HCV-infected liver biopsies analysed in (B) were compared. All liver biopsies with high miR-135a-5p levels exhibited low PTPRD mRNA expression and vice versa. Liver biopsies from patients infected with HCV are highlighted in red.
STAT3 transcriptional activity is induced in HCV-infected Huh7.5.1 cells in vitro and associated with attenuated PTPRD expression in vivo
PTPRD is a signal transducer and activator of transcription 3 (STAT3) phosphatase,27 and therefore dampened PTPRD expression should promote the interleukin 6-Janus Kinase–STAT3 (IL6-JAK–STAT3) signalling axis. STAT3 signalling is essential for liver regeneration33 and an integral part of the regulation of the host interferon response.10 Moreover, STAT3 is a cofactor for HCV infection, suggesting an accumulation of STAT3 activity in host cells during HCV infection.10 Indeed, a temporal GSEA of HCVcc-infected Huh7.5.1 transcriptomes revealed an accumulation of a STAT3 transcriptional signature (Hallmark_IL6 STAT3 signalling gene set25) over the first 7 days of infection (figure 4A). However, PTPRD is not expressed in hepatoma cells and cannot be rescued in our hands (data not shown), precluding the in vitro study of PTPRD on the induction of the IL6–JAK–STAT3 signalling cascade. Therefore, we analysed a publicly accessible gene expression database of 82 adjacent non-tumour liver tissue specimens of patients with HCC (cohort A).17 Those patients with the lowest 20 percentile of hepatic PTPRD expression in adjacent tissue showed a strong and marked enrichment (NES=1.75, FDR=0.009) of the STAT3 transcriptional programme compared with patients with the highest 20 percentile (see figure 4B, C and online supplementary figure S3), which is testimony of a PTPRD-mediated STAT3 deactivation in vivo. For 62 of these 82 patients, HCV infection status was available. We could thus confirm PTPRD expression downregulation in 46 HCV(+) biopsies compared with 16 HCV(−) biopsies (figure 4D) and show the enhanced STAT3 activity in HCV(+) through GSEA (NES=1.96, FDR=0.001) (see figure 4E and online supplementary figure S4). Taken together, these findings demonstrate that transcriptional activity of the oncogene STAT3 is clearly linked to PTPRD in the livers of patients. Because STAT3 activity is associated with HCCs with poor prognosis,34 ,35 ,36 our findings suggest that HCV infection accumulates STAT3 signalling via suppression of its negative regulator PTPRD and as such may contribute to the exacerbation of chronic liver disease.

Dampened protein tyrosine phosphatase receptor delta (PTPRD) expression in liver biopsies is associated with an accumulated STAT3 transcriptional programme. (A) HCV infection induces STAT3 transcriptional activity. Huh7.5.1 cells were subjected to infection with affinity-purified HCVcc (Jc1E2FLAG) for up to 7 days. Control cells were mock-infected using FLAG peptide. Cell transcriptome was profiled every day by RNA-seq as previously described,19 and the modulation of the Hallmark_IL6 STAT3 signalling gene set was assessed by Gene Set Enrichment Analysis (GSEA) in HCV-infected cells as compared with controls. Significant (p<0.005) normalised enrichment scores (NES) of the gene set are indicated as red (positive enrichment) for each day of infection (grey stairs). (B) Patients were classified according to PTPRD expression in tissue adjacent to hepatocellular carcinoma (HCC) into the highest 20 percentile (red), lowest 20 percentile (blue) and intermediate (grey). GSEA of these 82 adjacent liver biopsies (cohort A)17 revealed (C) a significant enrichment (NES=1.75, false discovery rate (FDR)=0.001) of the Hallmark_IL6 STAT3 signalling gene set (STAT3 signature) in biopsies with 20 percentile lowest PTPRD expression (B, blue). (D) In the adjacent tissues described in (B), PTPRD expression is significantly (p=0.01, U-Test) impaired in the HCV-infected (HCV(+), grey circles, n=46) versus confirmed HCV-negative (HCV(−), empty circles, n=16) biopsies and (E) associated with a significant enrichment (NES=1.96, FDR=0.001) of the STAT3 signature. Gene expression patterns of the leading edge genes of the STAT3 signatures in liver tissues are provided as online supplementary figures S3 and S4.
PTPRD expression is impaired in HCC and associated with survival of patients with HCC and decreased tumour recurrence after surgical resection
We observed that PTPRD mRNA is only marginally expressed in hepatoma cell lines such as Huh7.5.1. Moreover, it does not translate into detectable protein levels as assessed by western blot in a panel of hepatic cell lines tested including HepG2, Huh7, Huh7.5.1, differentiated HepaRG and non-differentiated HepaRG (data not shown), which suggests that impaired PTPRD is a hallmark of cell transformation. To ascertain whether PTPRD is also downregulated in HCC, we assessed PTPRD protein expression in six paired biopsies of patients infected and uninfected with HCC (see online supplementary table S1). In four out of six donors, PTPRD expression was downregulated or even completely absent in tumour lesions compared with paired adjacent tissue samples (figure 5A). To verify whether this downregulation was also specifically present in HCV-infected patients with HCC, we analysed PTPRD expression in paired tumour and adjacent liver tissue specimens.17 In this data set of patients undergoing surgical resection of liver tumours, information for 46 patients with HCV-associated HCCs could be retrieved. In 30 out of 46 paired biopsies was PTPRD expression downregulated in tumour lesions compared with adjacent non-tumoural tissue (figure 5B). This 30/46 ratio echoes the western blot results of paired HCC biopsies (figure 5A), and the observed downregulation in tumour tissue was statistically significant (p=0.01, two-sided Wilcoxon signed-rank test) (figure 5B). Next, we took advantage of the long-term clinical follow-up information provided in the database. Strikingly, when looking at all patients with HCC (n=82), those ranking in the highest 20 percentile levels of PTPRD expression in adjacent tissue (figure 4B) had a higher long-term survival rate (p=0.048, log-rank test) (figure 5C) and had less chance of recurrent liver cancer (p=0.02, log-rank test) (figure 5D), irrespective of cancer aetiology. We confirmed the same trend of prognostic association in two additional independent cohorts with shorter patient follow-up (see online supplementary figures S5 and S6).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Dampened protein tyrosine phosphatase receptor delta (PTPRD) expression in liver biopsies is associated with a decreased patient survival and increased tumour recurrence after surgical resection. (A) PTPRD is impaired in tumour lesions of patients with hepatocellular carcinoma (HCC). Expression of PTPRD and actin was assessed in HCC lesions (T), and the corresponding paired adjacent tissue (A) of six patients with HCC by western blotting using specific antibodies. Patient codes are indicated. A random primary human hepatocytes (PHH) lysate served as positive control. (B) PTPRD is significantly (p=0.01, two-sided Wilcoxon signed-rank test) impaired in HCV-associated HCCs. The gene expression database of cohort A was analysed for PTPRD expression in paired liver biopsies from 46 patients infected with HCV.17 Pairs of tumour lesions (dark grey) with the corresponding adjacent non-tumour tissue (light grey) are connected by a dashed line. Same data are summarised side by side as box-and-whisker plot (box=50% of biopsies, line=median, whiskers=minimal and maximal values) in the same panel. (C) Patients from cohort A with high PTPRD expression in adjacent non-tumour tissues (red) exhibit a significantly (p=0.021, log-rank test) higher survival rate from HCC and (D) a significantly (p=0.048, log-rank test) decreased tumour recurrence after surgical resection of the tumours. Product-limit estimation of PTPRD expression in adjacent non-tumour tissue of 82 patients compared with disease progression data from patients. Patient survival from HCC (C) and tumour recurrence after surgical resection (D) were compared between biopsies with highest PTPRD expression (figure 4B) (top 20 percentile; red) and biopsies exhibiting low PTPRD expression (lowest 20 percentile; blue) using Kaplan-Meier estimator. Number of patients at risk (No. at risk) is indicated.
Discussion
HCC is the second largest cause of death from cancer worldwide and poses an increasing burden on global health.37 Chronic HCV infection is a main cause of HCC, and even though recent pharmacological breakthroughs can efficiently eradicate HCV infection, the risk of developing HCC in patients after sustained virological response remains elevated. Moreover, given the epidemiological history of the virus, HCV-associated complications are set to peak over the next decade, and novel preventive strategies are urgently needed. By studying the impact of HCV infection on protein phosphatase expression patterns in liver biopsies, we aimed to identify drivers for HCV-associated disease development. Here, we show that tumour suppressor PTPRD is consistently downregulated upon HCV infection, both in vivo, in chronically infected patient biopsies, and in vitro in infected PHH. PTPRD is a well-established tumour suppressor27 whose chromosomal locus on 9p23–24.1 is regularly subject to genetic deletion or epigenetic inactivation in a broad spectrum of human malignancies including neuroblastoma, glioblastoma,27 lung cancer, cutaneous squamous cell carcinoma,38 laryngeal squamous cell carcinoma,39 melanoma40 and also HCC.28 In addition, PTPRD copy number loss associates with a poor prognosis in breast cancer, colon cancer41 and gastric adenocarcinoma.42 Here, we demonstrate for the first time that PTPRD protein expression is downregulated in patients chronically infected with HCV. Moreover, we show that this downregulation is even more pronounced in tumour lesions of paired liver biopsies compared with adjacent non-tumour tissue of non-infected patients. Taken together, the evidence suggests that a gradual loss of PTPRD expression is a common event in liver disease progression, and it highlights the role of PTPRD as a potential suppressor of hepatocarcinogenesis, irrespective of aetiology.
In this study, we identified a HCV-specific regulatory mechanism that silences PTPRD expression in vivo. We demonstrate that PTPRD mRNA is targeted by miR-135a-5p, a miRNA that is expressed in the liver (see online supplementary figure S7) and that was described to be markedly elevated in HCC lesions from patients with poor prognosis.43 ,44 We demonstrate that miR-135a-5p expression is also significantly elevated in liver biopsies of patients with HCV (figure 3A, B). Interestingly, miR-135a has been recently suggested as a cofactor for HCV replication by interacting with the 5′UTR of the viral genome.45 Our data demonstrate HCV-induced upregulation of miR-135a-5p expression is a cause of PTPRD silencing (figure 3D) and that high miR-135a-5p levels in liver tissues significantly (p=0.04, one-tailed Spearman's correlation test) correlate with low PTPRD levels and vice versa. The existence of a population of samples where both miR-135a-5p and PTPRD are weakly expressed (figure 3E) suggests the existence of additional regulatory mechanisms of PTPRD expression that may as well involve HCV-dependent or independent epigenetic modification of the PTPRD promoter or histones. These findings support the view that miR-135a-5p controls PTPRD in HCV-infected livers, potentially together with additional so far unidentified mechanisms for PTPRD mitigation.
Interestingly, the transcription factor STAT3 is a confirmed target for PTPRD and a key player during liver regeneration. Loss of PTPRD function leads to aberrant STAT3 phosphorylation in glioblastoma.27 ,46 Here we demonstrate a significant association of PTPRD levels with STAT3 transcriptional activity in the livers of patients (figure 4). We have previously demonstrated that STAT3 is an indispensable host factor for HCV infection and that the viral infection is promoted by STAT3 activation.10 Thus, it is conceivable that HCV downregulates PTPRD in order to benefit from a STAT3-driven transcriptional programme. Since STAT3 activity also plays a role in liver disease progression and HCC development,34 ,35 ,36 ,47 this may contribute to the protumourigenic environment during chronic HCV infection. In other words, our model suggests the existence of a perturbed PTPRD–STAT3 axis driving malignant progression of liver disease. We believe that although not associated to other hallmarks of HCC development like inflammation and fibrosis (figure 1), a perturbed PTPRD–STAT3 axis adds additional oncogenic pressure to the liver as STAT3 is associated to HCCs with poor prognosis.34 ,35 ,36 This finding may also be of further clinical relevance since it provides a target for HCC chemoprevention. Indeed, therapeutic intervention on signalling events constitutes a new chemopreventive strategy, as proof of concept has been demonstrated using the clinical epidermal growth factor receptor inhibitor erlotinib to attenuate liver fibrosis and the development of HCC in an animal model.48 Given that PTPRD expression is suppressed by chronic HCV infection and associated with HCC and patient survival, our data suggest a PTPRD-centred signalling network as a potential target for novel chemopreventive strategies for HCV-induced HCC.
Acknowledgments
The authors thank Dr Sébastien Pfeffer (UPR 9002, IBMC, University of Strasbourg, France) for providing small RNA sequencing data of HCVcc-infected Huh7.5.1 cells, Dr Michel Tremblay (McGill University, Montreal, Canada), Dr Catherine Schuster (Inserm U1110, University of Strasbourg, France) and Luc Dupuis (Inserm U1118, University of Strasbourg, France) for helpful discussions; Sarah C. Durand (Inserm U1110, University of Strasbourg, France) for isolation of primary human hepatocytes and excellent technical assistance. Moreover, they thank Emilie Heuillard, Dr Eric Robinet (Inserm U1110, IHU, University Hospital Strasbourg) and the Biological Resource Centre (Centre de Ressources Biologiques (CRB) of the University Hospital Strasbourg, France) for managing the biobank; Dr Frank Chisari (The Scripps Research Institute, La Jolla, California, USA) for the Huh7.5.1 cells; Dr Takaji Wakita (National Institute of Infectious Diseases, Tokyo, Japan) for the HCV JFH1 strain; Charles M. Rice (The Rockefeller University, New York City, New York, USA) and François Helle (Université de Picardie Jules Verne, Amiens, France) for the RFP–NLS-IPS lentivirus. The small RNA-seq analysis of HCV-infected Huh7.5.1 cells was performed at the IGBMC Microarray and Sequencing Platform (Illkirch, France), member of the France Genomique programme.
References
Footnotes
Contributors JL initiated and supervised the study. JL and TFB obtained funding. NVR, AARS, FHTD, JL, CG, DC, NF and AA designed and conducted experiments and analysed data. TP, DS, VD, PP and MHH provided patient liver tissue and corresponding clinical data. AO, KC, MF, HN and YH provided gene expression data from patient cohorts and performed patient survival analysis. SB and MBZ analysed HCV-induced miRNA expression in Huh7.5.1 cells and designed miRNA reporter assays. TC, NP and NVR performed GSEA analysis. NVR and JL wrote the manuscript. MHH, YH, MBZ and TFB critically reviewed the manuscript.
Funding This work was supported by grants from the Wilhelm Sander-Stiftung (Förderantrag 2010.023.1 to TFB), the European Union (Interreg IV-Rhin Supérieur-FEDER-Hepato-Regio-Net to TFB and MBZ; ERC-AdG-2014 HEPCIR to TFB; EU H2020 HEPCAR to TFB), the French Cancer Agency (ARC IHU201301187 to TFB), the University of Strasbourg (IdEx, Projet Attractivité 2014, ANR, to JL) and from the French ANRS (ECTZ4236, ECTZ4446 to JL and AARS). NVR was funded by an IdEx fellowship from the University of Strasbourg (IdEx, Contrat Doctoral 2012, ANR, to MBZ and TFB). This work has been published under the framework of the LABEX ANR-10-LAB-28 and benefits from a funding from the state managed by the French National Research Agency as part of the investments for the future programme.
Competing interests None declared.
Ethics approval Ethics Committees of the University Hospital of Basel (Switzerland), the Centre Hospitalier Universitaire de Reims (France) and the Hôpitaux Universitaires de Strasbourg (France).
Provenance and peer review Not commissioned; externally peer reviewed.