Article Text

Abstract
Background and aims Prenatal and early life bacterial colonisation is thought to play a major role in shaping the immune system. Furthermore, accumulating evidence links early life exposures to the risk of developing IBD later in life. We aimed to assess the effect of maternal IBD on the composition of the microbiome during pregnancy and on the offspring’s microbiome.
Methods We prospectively examined the diversity and taxonomy of the microbiome of pregnant women with and without IBD and their babies at multiple time points. We evaluated the role of maternal IBD diagnosis, the mode of delivery, antibiotic use and feeding behaviour on the microbiome composition during early life. To assess the effects of IBD-associated maternal and infant microbiota on the enteric immune system, we inoculated germ-free mice (GFM) with the respective stool and profiled adaptive and innate immune cell populations in the murine intestines.
Results Pregnant women with IBD and their offspring presented with lower bacterial diversity and altered bacterial composition compared with control women and their babies. Maternal IBD was the main predictor of the microbiota diversity in the infant gut at 7, 14, 30, 60 and 90 days of life. Babies born to mothers with IBD demonstrated enrichment in Gammaproteobacteria and depletion in Bifidobacteria. Finally, GFM inoculated with third trimester IBD mother and 90-day infant stools showed significantly reduced microbial diversity and fewer class-switched memory B cells and regulatory T cells in the colon.
Conclusion Aberrant gut microbiota composition persists during pregnancy with IBD and alters the bacterial diversity and abundance in the infant stool. The dysbiotic microbiota triggered abnormal imprinting of the intestinal immune system in GFM.
- inflammatory bowel disease
- pregnancy
- early life microbiome
Statistics from Altmetric.com
Significance of this study
What is already known on this subject?
A positive family history remains the strongest risk factor for developing IBD; however, genetic susceptibility alone explains a small proportion of disease heritability.
IBD affects women during their reproductive years, and around 25% become pregnant after their initial diagnosis. Human studies have shown that profound changes in the microbiome occur during pregnancy and that maternal health status and maternal microbiome play a role in shaping the microbiome and immune system of the neonate.
The role of IBD in the maternal microbiome composition during pregnancy and its impact on the offspring’s microbiome remains unclear.
Significance of this study
What are the new findings?
Women with IBD maintain altered bacterial diversity and composition in their gut during pregnancy as compared with controls, with an enrichment in Gammaproteobacteria and a depletion in Bacteroidetes.
Babies born to IBD mothers present with lower diversity and altered bacterial gut composition up to at least 3 months of life, characterised by an enrichment of Gammaproteobacteria and a depletion of Bifidobacteria, independent of other exposures, compared with babies born to control mothers.
The altered microbiota induced changes within the adaptive immune system of the intestines in germ-free mice, suggesting that maternal IBD may have impact on the immune system of the offspring through microbial factors.
How might it impact on clinical practice in the foreseeable future?
Targeting dysbiosis in pregnant women with IBD or during early infancy could foster the development of a healthy microbiome in the offspring, thereby reducing the future risk of IBD.
Introduction
IBDs, comprising Crohn’s disease (CD) and UC, result from aberrant mucosal immune responses to bacteria in genetically susceptible individuals.1 2 While family history is the strongest risk factor for developing IBD, the genetic loci associated with risk do not fully explain disease heritability.1 IBD affects women during their reproductive years and 25% become pregnant after diagnosis. Some studies have suggested that there may be a higher disease transmission rate from mothers with IBD as compared with fathers.3 4 Dynamic changes in the diversity and abundance of the microbiome have been observed during pregnancy coinciding with adjustments in maternal immunity.5 While this balance is likely more complicated in pregnant women with IBD, it is largely unknown how pregnancy may affect the microbiota of patients with IBD. Moreover, increasing evidence suggests that maternal health status and microbiota during pregnancy may influence the infant’s initial gut colonisation,6 playing a key role in shaping the immune system development.7 Of interest, early life exposures, many of which have an impact on microbiota development, have been linked to the future risk of developing IBD.8–11 However, no information exists on the impact of maternal IBD on the development of the offspring’s gut microbiome. Therefore, we designed a study aimed at characterising the microbiota of pregnant women with and without IBD and of their babies. We then compared the effect of the maternal and infant microbiome on the priming of the immune system of germ-free mice (GFM), a model to study interactions between the microbiome and the nascent immune system.12
Methods
Study design and sample and data collection
Participants for this study were selected from the prospective Exploring MEChanisms Of disease traNsmission In Utero through the Microbiome (MECONIUM) study. Pregnant women with IBD, and age and race/ethnicity-matched controls were recruited after signing an informed consent. Exclusion criteria included inability to provide consent, HIV/AIDS, autoimmune diseases, fetal chromosomal or structural abnormalities and active infection during pregnancy. After enrollment, pregnant women self-collected stool and saliva samples at each trimester. Following delivery, serial stool samples were collected from the infants at 7, 14, 30, 60 and 90 days (figure 1). We chose these early time points due to a relatively uniform feeding behaviour, as well as limited person-to-person contact and exposures associated with walking or crawling. Demographics and relevant clinical history were collected at the time of recruitment (baseline) and at each time point concurring with sampling. For women with IBD, disease type and surgical history were recorded. Additionally, at each trimester, medications (including vitamins and supplements), changes in medication and disease activity (Harvey Bradshaw index13 and physician’s global assessment for CD, and the Mayo score for UC14) were also registered. At delivery, gestational age, mode of delivery, complications and medications, baby’s sex, birth weight and need for neonatal intensive care were recorded. Preterm birth was defined as delivery <37 weeks of gestation and low birth weight as a birth weight ≤2499 grams. Information on infants’ feeding behaviour (exclusive formula feeding, exclusive breastfeeding or mixed) and antibiotic use during follow-up was prospectively obtained at the time of each sampling. Study data were collected and managed using Research Electronic Data Capture electronic data capture tools.15
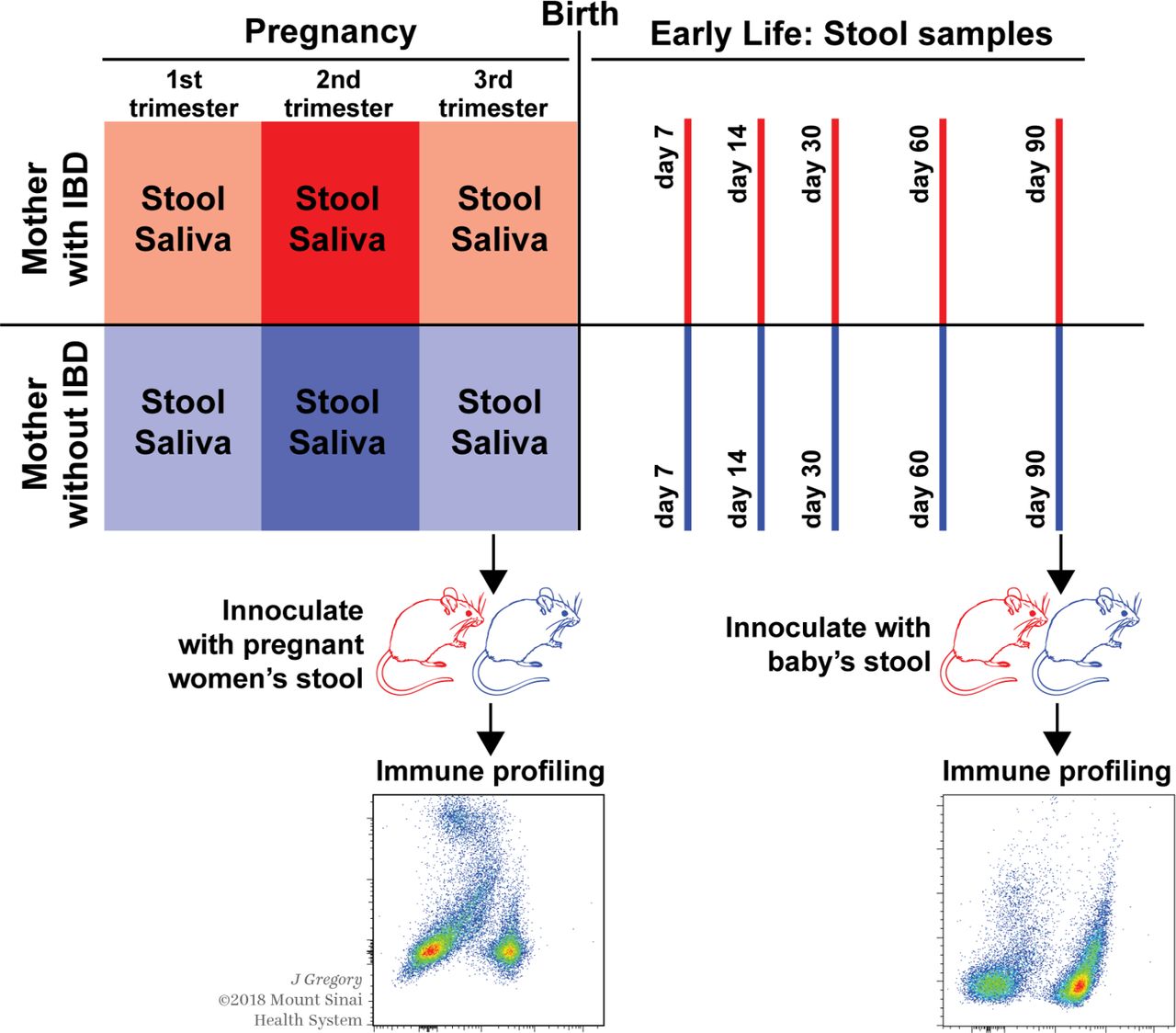
Study design: pregnant women with (upper panel) and without (lower panel) IBD recruited for the study collected a stool and saliva sample at each trimester, coupled with clinical and obstetric history. After delivery, their infants were followed up with serial stool samples and extensive health and exposure metadata at various time points. Stool from mother–baby pairs was selected and gavaged into wild-type germ-free mice aged 6–8 weeks to conduct immune phenotyping.
Sample processing and 16S rRNA sequencing
Total DNA was isolated from each specimen using a bead beating method with the PowerSoil DNA Isolation Kit (Mobio, Carlsbad, California, USA). After quantification by Qubit fluorometer, the DNA was amplified at the phylogenetically informative V3-V4 region of bacterial 16S rRNA gene using dual-barcoded universal primer 347F/803R as described previously.16 The integrity of the amplicons was further verified by precast agarose gel electrophoresis. The resulting ~460 bp sized amplicons were pooled with equal molarity and then sequenced on the Illumina HiSeq2500 platform using the fast-mode pair-end 250 protocol. Sequentially, the 16S reads were merged and filtered by size (>400 bp) and quality score (>Q30) using PANDAseq (PAired-eND Assembler for DNA sequences).17 The processed reads were further split by dual barcodes for each sample and then assigned to operational taxonomic units (OTUs) using open-reference OTU picking method with a default cut-off of 97% similarities and taxonomic classification using the Quantitative Insights Into Microbial Ecology (QIIME) pipeline.18 Duplicate samples were included to assess sequencing reproducibility.
Microbiome analysis of human samples
We calculated the core diversity metrics at each time point and across body sites. Specifically, α-diversity on rarefied tables was estimated using Faith’s phylogenetic diversity,19 and significance between groups was determined using non-parametric tests. Microbial diversity between samples (β-diversity) was evaluated with QIIME using unweighted UniFrac distances20 and visualised using multidimensional scaling plots. Multifactorial permutational analysis of variance (PERMANOVA)21 (Adonis in R package (vegan)) was used for each time point to test the association of maternal IBD status with the microbiome β-diversity while adjusting for covariates (feeding behaviour, mode of delivery, antibiotic exposure, preterm birth and baby’s sex) and multiple hypothesis testing. The linear discriminant analysis effect size (LEfSe) algorithm22 was used to identify taxa associated with maternal IBD status. For infants, samples from all time points were pooled, and the differentially abundant features were ranked by effect size after adjusting for delivery mode due to significant effect of this variable on bacterial diversity. Taxa with ≥2-fold differences by maternal IBD were selected. All statistical tests were two sided. To confirm robustness of our findings, MaAslin (Multivariate Association with Linear Models (MaAslin) was used in third trimester maternal samples for multivariate assessment of associations between taxa abundance and metadata, using default parameters.23 A linear mixed model was applied to the longitudinal offspring 16S sequencing data to estimate the predictors of the most differential bacterial taxa identified in the LEfSe analysis while adjusting for constraint and time-varying covariates. Maternal IBD status, mode of delivery and time of sampling were treated as fixed effects, with a random intercept for each subject ID. Feeding behaviour and antibiotics exposure were entered as time-varying covariates. The model was fitted via restricted maximum likelihood approach with the correlation structure of autoregressive process of order 1.
Microbiota transfer experiments in GFM
Donor selection and slurry preparation
To understand the functional consequences of altered microbiome in pregnant women and their offspring, stool samples from eight CD mother–baby pairs (16 individuals) and from three control mother–baby pairs (six individuals), for a total of 22 donors, were selected for microbiota transfer and gavaged into a total of 78 C57Bl/6 GFM. Donor selection included availability of paired stool samples from mother at third trimester and her baby at 90 days, full-term pregnancies and, for IBD mothers, inactive CD with similar medication patterns. Further details on donor features can be found in online supplementary table S1. Faecal samples were pulverised under liquid nitrogen; working under strict anaerobic conditions, samples were blended into a 40–60 mg/mL slurry in prereduced bacterial culture media. Infant stool samples collected on diaper material were suspended by repeated vortexing in 20 mL of prereduced bacterial culture media with 5 mL sterile 3 mm glass beads. Beads and diaper debris were removed by passing through a 100 µm strainer. Faecal slurries were diluted 1:20 in bacterial culture media and stored at −80°C in 15% glycerol until use.
Supplementary file 1
GFM gavage and sample collection
C57Bl/6 GFM were raised in gnotobiotic isolators and colonised at 6–8 weeks of life by gavage with 200–300 µL of recently thawed human faecal slurry; thereafter, mice were maintained outside the isolator in sterile filter-top cages and handled using strict aseptic techniques.24 25 Eight groups of mice (48 mice) were gavaged with the stool from CD mothers or their respective 3-month-old baby (2–5 GFM per donor). In addition, three groups of mice (30 mice) were gavaged with stool from non-CD, control pregnant women or their 3-month-old baby (3–5 GFM per donor). Five weeks after gavage, mice were euthanised by CO2; mesenteric lymph nodes (MLNs) and large intestine were harvested for immune profile analyses by flow cytometry. Additionally, murine stool samples were collected for 16S rRNA sequencing, following the protocol described above.
Single cell preparation from MLN, spleen and the intestinal lamina propria of mice
Initially, we examined the small intestinal lamina propria, colonic lamina propria, Peyer’s patches and MLN separately (as components of the mucosal immune system including innate lymphoid cells (ILCs)). Additionally, splenocytes were evaluated to represent the systemic immune system. The MLN and spleen were harvested in RPMI media containing collagenase D (Sigma-Aldrich, final concentration 400 IU/mL) and physically disrupted using a 25-gauge syringe. Cell suspension was placed in an incubator at 37°C for 30 min followed by addition of 10 mM EDTA (Invitrogen) and two rounds of washing with RPMI (Gibco).
Single cell suspension from the intestinal lamina propria was prepared as previously described with minor modification.26 Small intestines and colon were harvested and processed separately. Briefly, after Peyer’s patches were removed from the small intestines. Mesenteric fat and the faecal contents were carefully removed from intestines, and the lumen was exposed using a longitudinal incision. The small and large intestinal tissues were washed with Hank’s Balanced Salt Solution (HBSS) for three times, then cut into 1 cm pieces and incubated with 5 mM EDTA (Invitrogen) in HBSS at 37°C for 20 min followed by washing with HBSS for three times. Next, the tissue was minced with scissors and transferred to conical tubes with 10 mL of complete RPMI containing 0.5 mg/mL of collagenase (Sigma-Aldrich) and incubated at 150 rpm for 30 min at 37°C. The single cell suspension was collected and passed through a 100 µm and 40 µm cell strainer and washed with RPMI twice.
After initial experiments, we focused on differences in immune populations within colonic tissues as representative of the intestinal immune system.
Flow cytometry analysis
To prevent non-specific staining, the single cell suspension from MLN and colonic lamina propria was incubated with CD16/32 antibody (2.4G2, BioXcell). At the outset, we characterised T cell subsets, B cell subsets, dendritic cell (DC) subsets and ILCs using panels of fluorescent dyes conjugated with monoclonal antibodies (online supplementary table S2). LIVE/DEAD Fixable Aqua Dead Cell Stain Kit (Invitrogen) was used to determine the viability of cells. Foxp3 staining was performed after fixation and permeabilisation using the Foxp3/transcription factor staining buffer set (Invitrogen). Results from the colonic lamina propria were representative of changes in the mucosal immune system. Therefore, after initial experiments, we focused on analyses of the colonic lamina propria only in the GFM. Samples were acquired using LSR Fortessa (BD Bioscience), and data were analysed using FlowJo V.10 (FlowJo).
ELISA for stool IgA
Murine stool (50 mg) was incubated in 500 µL of PBS at room temperature for 10 min, followed by vortexing for 5 min and bead-beating for 2 min. This step was repeated twice, and the sample supernatants were collected after centrifugation. IgA ELISA was performed by using the Mouse IgA ELISA Quantitation Set (Bethyl Laboratories) according to the manufacturer’s protocol. A MaxiSorp ELISA plate (Thermo Fisher Scientific) was precoated with 100 µL of diluted purified IgA antibody in 0.1M carbonate buffer (10 µl/mL). After overnight incubation at 4°C, the plate was washed with PBS and incubated with PBS containing 1% BSA for 1 hour at room temperature to prevent non-specific response. After washing, 100 µL of 1:200 diluted stool supernatants were plated and incubated for 1 hour. The plates were washed again and 100 µL of antimouse IgA-HRP (1:40 000 dilution) was added. After 1 hour incubation, 100 µL of TMB substrate (BD Pharmingen) was added, followed by 100 µL 1M H2SO4 (as a stop solution). The absorbance (at 450 nm) was measured using an ELISA reader.
Results
Characteristics of study participants
We included the first 200 consecutive participants from the MECONIUM study: 121 pregnant women (40 with IBD), and 79 babies (26 born to mothers with IBD), resulting in a total of 73 mother–infant dyads. The demographic and clinical characteristics of pregnant women were evenly distributed by IBD status with regards to maternal age, gestational age at recruitment, pregnancy complications, use of assisted reproductive technologies and exposure to antibiotics during pregnancy or at delivery (table 1).
Demographic and clinical characteristics of study participants
Mothers with IBD were more likely to be Jewish (p=0.008) and nulliparous (p=0.024). Most pregnant women with IBD (70.0%) presented with inactive disease. When occurred, flares were mostly mild. Patients with CD had a more frequent history of bowel resection (p=0.02) compared with UC and demonstrated differences in medication (online supplementary table S3). Except for a higher proportion of male babies born to IBD mothers (p=0.013; table 1), rates of preterm birth, low birth weight or exposure to antibiotics or probiotics after birth were similar across groups (table 1).
Women with IBD present persistent dysbiosis during pregnancy
After quality control, 619 samples with >1000 sequencing reads each were available for downstream analysis (total read counts: 8 857 479; mean±SD/sample: 133 581±13 977). As expected, diversity was driven primarily by body sites (online supplementary figure S1a-d). Throughout pregnancy, women with IBD had lower gut α-diversity than controls, with differences reaching statistical significance at the first (T1) and second (T2) trimesters of pregnancy (p=0.005 and p=0.01, respectively) but not at the third (T3) trimester (figure 2A). We observed opposite trends in the changes over time of the global α-diversity throughout pregnancy by IBD status: while control women presented a continuous decrease from T1 to T3 (PD_whole tree in T1: 51.2±8.4 vs T3: 45.0±8.6, Wilcoxon p=0.047), patients with IBD demonstrated a non-significant trend towards a slight increase (PD_whole tree in T1: 40.2±7.1 vs T3: 43.8±7.9, p=0.17) (figure 2A). PERMANOVA analysis based on the unweighted UniFrac distances revealed significantly different β-diversity of the gut microbiota by IBD status at each trimester (p=0.017, 0.036 and 0.003 for T1, T2 and T3, respectively; figure 2B). Biomarker analysis comparing the relative abundances of the gut microbiome by IBD diagnosis showed that the major differences were driven by an enrichment of the Gammaproteobacteria class and a depletion of the Bacteroidetes phylum in mothers with IBD as compared with controls (figure 2C). In T3 alone, an unidentified genus from the Enterobacteriaceae family was enriched in pregnant women with IBD (unadjusted p=10−5, Bonferroni corrected p=0.02); corroborating this finding, the LEfSe analysis showed 14 taxa under the Proteobacteria phyla to be enriched (online supplementary figure S2). Further analysis, using MaAslin, conducted in T3 samples only (n=80) and taking into account participant type, race, parity and exposure to antibiotics during pregnancy confirmed a significant enrichment in Enterobacteriaceae family associated with maternal IBD status (false discovery rate (FDR) adjustedp value: 0.03). No significant differences were observed between mothers exposed to thiopurines or biologicals during pregnancy or in those with active versus inactive disease (data not shown).

The gut microbiota composition of pregnant women by IBD diagnosis. (A) The boxplots show the mean and variance of the richness of the microbial community (α-diversity) between pregnant women with (red) and without (blue) IBD (PD_whole tree); significant differences were seen at the first (T1) (p=0.005) and second trimester (T2) (p=0.01) but not in third trimester (T3). (B) Overall microbiota dissimilarities (ß-diversity) between samples grouped by IBD status. Dissimilarities were measured using unweighted UniFrac distances and visualised using a non-metric multidimensional scaling plot. Significant differences in the ß-diversity were observed at each trimester of pregnancy between pregnant women with (red) and without (green) IBD. (C) Differential microbial features between pregnant women with and without IBD as determined by linear discriminant analysis effect size analysis (linear discriminant analysis).
No significant differences in the α-diversity or β-diversity of the oral microbial community were observed within subjects throughout pregnancy or between IBD patients and controls at any trimester (data not shown).
Maternal IBD status predicts the diversity and taxonomy of the infant microbiome
At the time of our analysis, 79 babies had been born and followed up for up to 3 months, with 306 high-quality stool samples available for analysis. Overall, the bacterial diversity changed over time in the pooled set of infant samples (online supplementary figure S3). The gut microbiome of infants born to IBD mothers demonstrated lower diversity and variation in the overall abundance compared with babies born to control mothers (figure 3). Specifically, significant differences in α-diversity were detected at days 7, 14 and 90 based on maternal IBD status (Wilcoxon p=0.05, 0.03 and 0.02, respectively; figure 3A). Furthermore, while maternal IBD diagnosis was a significant predictor of baby’s β-diversity (figure 3B) (univariate and multivariate PERMANOVA p<0.05 for each time point), mode of delivery, antibiotic exposure, type of feeding and preterm birth were also significantly associated with the overall bacterial abundance at various time points (table 2). Stratified analysis in babies born vaginally and via C-section showed that the maternal IBD diagnosis remained a significant predictor of microbiome diversity regardless of the delivery mode. Likewise, differences remained after excluding all samples from babies that were exclusively formula fed (online supplementary figure S4).
Statistical significance for predictors of early life microbiota ß-diversity by time point
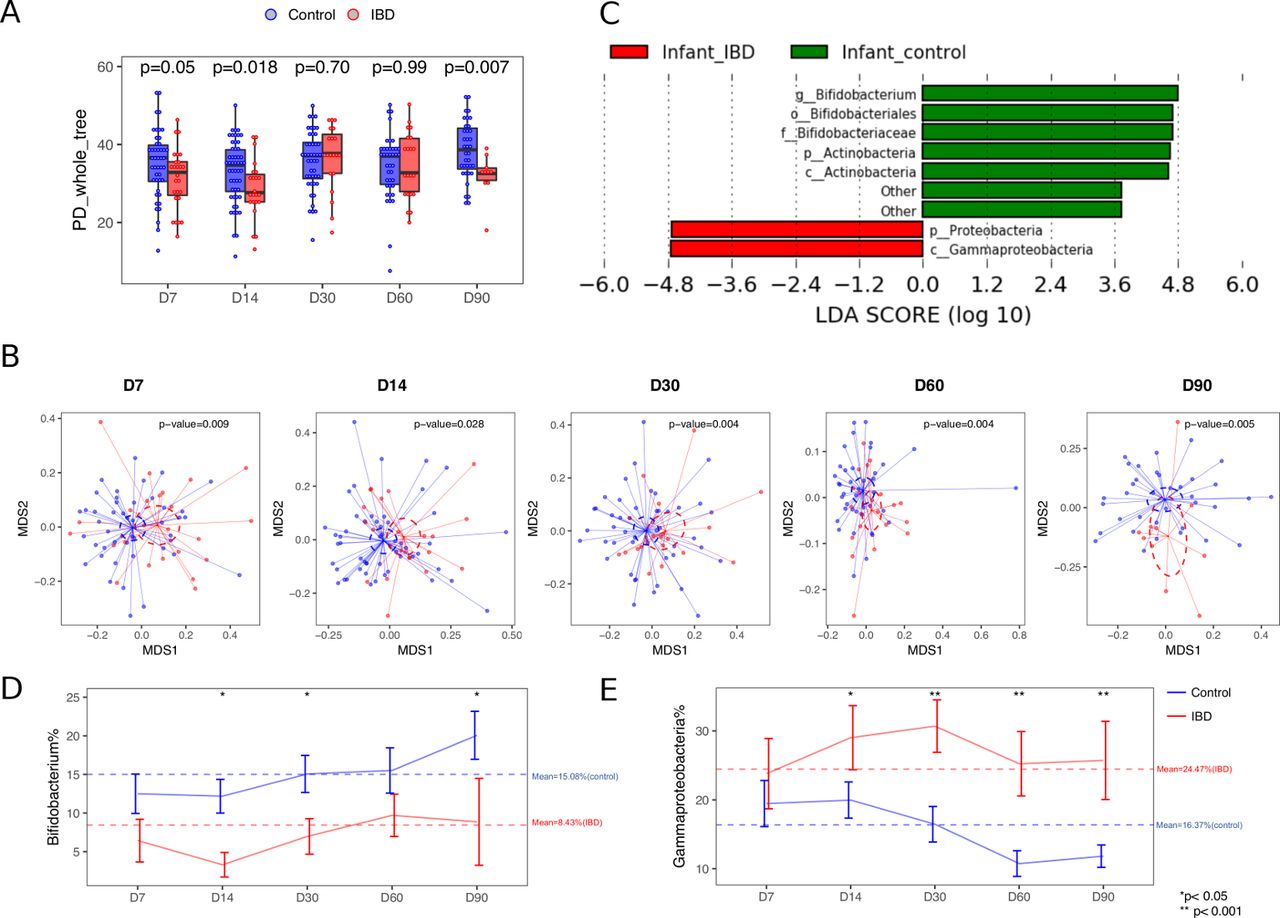
Microbiota profiles across multiple time points during early life by maternal IBD status. (A) The α-diversity of the microbial community within samples obtained at different time points after delivery and compared between babies born to mothers with (red) and without (blue) IBD. The α-diversity was measured using PD_whole tree obtained in rarefied tables at 1000 sequences. (B) The ß-diversity measured by unweighted UniFrac distances at different time points after delivery compared between babies by maternal IBD status. (C) Top discriminative bacteria between babies born to mothers with (red) and without (green) IBD as determined by the LEfSe analysis. (D and E) Representative differential bacterial taxa over time by maternal IBD status. Error bars indicate mean±SD. LDA, linear discriminant analysis; LEfSe, linear discriminant analysis effect size.
We then conducted LEfSe analysis across all time points while adjusting for the delivery mode, one of the major predictors of the early life microbiome. We detected that babies born to IBD mothers presented with an enrichment in the Gammaproteobacteria class bacteria and a reduction in the Bifidobacterium genus, as compared with those born to control mothers (figure 3C), and these differences persisted for the 3-month duration of the study (figure 3D,E). We next tested the impact of different exposures on the relative abundance of the differentially abundant taxa over time. We found that maternal IBD diagnosis and exclusive formula feeding were associated with a reduction in the relative abundance of beneficial Bifidobacterium genus over time (on the log scale 1.4-fold reduction, p=0.007 and 1.7-fold reduction, p=0.048, respectively; table 3). After adjustment for the covariates, maternal IBD status was the only significant predictor of the relative abundance of the colitogenic Gammaproteobacteria taxa over the first 3 months of life (p=0.002) (table 3).
Results of the linear mixed regression model applied to the longitudinal changes in relative abundance of Bifidobacterium and Gammaproteobacteria during the first 3 months of life
IBD status determines the composition of the adaptive immune system in GFM
We first compared the overall bacterial abundance (β-diversity) in the stool of GFM inoculated with the maternal and infant stools (design shown in figure 1) and detected significant differences by donor maternal IBD status (p=0.0001 and p=0.003, respectively, after adjustment for batch and repeated sampling; online supplementary figure S5a-b).
Next, we characterised subsets of T cells, B cells, dendritic cells and ILCs in the intestines of GFM inoculated with maternal or infant stools. Non-IBD mother/baby stools and uninoculated GFM served as controls. Due to the good concordance in the results from the small intestinal lamina propria and the large intestinal lamina propria, we chose to focus on analyses of the colonic lamina propria only. We did not detect any significant differences between the spleen, Peyer’s patches and MLN nor did we observe variation in the dendritic cells (DC) subsets or ILCs between GFM colonised with the stool of IBD pregnant women or their infants compared with no-IBD maternal or corresponding infant stool (data not shown).
Among lymphoid cells, while we did not detect differences in the total T cells as well as CD4+ T cells, there was a significant reduction in CD19+ B cells in the mesenteric lymph node of mice gavaged with the stool of mothers with IBD (online supplementary figure S6).
Additionally, we identified consistent and reproducible differences in the adaptive immune system. Specifically, in the colonic lamina propria of mice inoculated with stool of pregnant women with CD and their 3-month-old infants, the proportion of class-switched memory B cells (live CD45+CD19+CD138−IgD−IgM− cells; online supplementary figures S7 and S8) was significantly lower as compared with the mice inoculated with control maternal or 3-month infant stool, respectively (both p<0.005) (figure 4a). Similar results were observed in MLN after inoculation with CD-exposed baby stool, but not CD maternal stool. Furthermore, among switched memory B cells, the homeostatic IgA+ switched memory B cells were significantly reduced in the colonic LP of mice colonised with either CD mother or their baby stool (p<0.01) but not in MLN (figure 4b). We did not observe significant differences in IgA+ plasma cells or stool IgA levels between the mice gavaged with CD or non-CD stool (online supplementary figure S9).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Germ-free mice inoculated with the stool of mothers with CD and their 3-month-old babies have significant abnormalities in the adaptive immune cells compared with mice inoculated with stool from non-CD controls. (A) Class-switched memory B cells (aqua−, CD45+CD19+CD138−IgD−IgM−) isolated from the colonic lamina propria and mesenteric lymph node (MLN) of GFM, which were inoculated with the stool of non-CD mother, CD mother or their 3-month-old babies (non-CD baby or CD baby). Cumulative data showing the percentage of class-switched B cell among live CD19+ CD138− B cells in the indicated groups of colonic lamina propria or MLN. (B) Cumulative data showing the percentage of IgA+ class-switched B cells (aqua−, CD45+ CD19+ CD138− IgD− IgM− IgA+) among live CD19+ CD138− B cells in the indicated groups of colonic lamina propria or MLN. (C) Cumulative data showing the percentage of regulatory T cells (TREG) (aqua−, CD45+ CD3+CD8−CD4+ FoxP3+) among live CD45+ CD3+ CD8− CD4+ cells in the indicated groups of colonic lamina propria or MLN. *P<0.05, **p<0.01, ***p<0.005. CD, Crohn’s disease; NS, not significant.
We also observed a reduced frequency of regulatory T cells (TREG, live CD45+CD3+CD8−CD4+Foxp3+ cells, online supplementary figures 7 and 8 in the colonic LP (p<0.005) and MLN (p<0.05) of mice inoculated with CD maternal stool compared with control maternal stool; no differences were found between the mice inoculated with the infant stool by maternal CD diagnosis (figure 4C).
Discussion
Our findings show that the microbiome of mothers with IBD was characterised by a lower α-diversity during the first and second trimester of pregnancy and a different β-diversity during each of the three trimesters compared with control mothers (figure 2A,B). The differences were driven by a depletion in the relative abundance of Bacteroidetes and an increase in the relative abundance of Proteobacteria (figure 2C). Importantly, we observed that the offspring of IBD mothers presented with a reduced bacterial diversity as early as by the end of the first week of life and demonstrated differences in the overall gut bacterial composition persisting for up to 3 months of life compared with control babies (figure 3A,B). These data are in line with previous studies reporting the impact of maternal diseases, such as obesity,27 eczema,28 HIV status29 and gestational diabetes,30 on the offspring’s microbiome. In our study, early-life microbial alterations were driven by enrichment in Gammaproteobacteria and a reduction in Bifidobacterium at each time point (figure 3C–E), mirroring some of the patterns observed in their mothers, as well as in adult and childhood-onset IBD patients.31 32 Bifidobacteria are among the first colonisers of the infant gut and are believed to promote health benefits for the host, stimulating maturation of the immune system and contributing to defence against enteropathogens.33 Bifidobacteria species have been shown to protect against development of colitis,34 and in patients with IBD, multistrain Bifidobacterium-containing probiotics have been shown to be effective in specific settings.35 However, Proteobacteria are often associated with intestinal inflammation36 and have been consistently linked to IBD.23
While maternal IBD status was the major and most consistent factor associated with baby’s microbiome composition at each time point, other factors, such as mode of delivery, feeding behaviour, exposure to antibiotics and preterm birth, were also found to affect the overall microbial composition. C-section and exposure to antibiotics during pregnancy and early life have been shown to increase the risk of IBD later in life,37 38 while breastfeeding in infancy has been shown to be protective against the development of IBD.8–11 39 Moreover, when analysing the most differentially prevalent taxa between babies by maternal IBD status over time, exclusive breastfeeding was independently associated with higher relative abundance of Bifidobacterium, which is consistent with prior reports.40
Pregnancy and early life are a critical period during which contact with microbes and specific microbe-associated metabolites are essential for the priming of the immune system development and maturation.41 Therefore, to assess functional consequences of the observed IBD-associated dysbiosis, we inoculated GFM, with the stool of pregnant women with and without CD and their babies.42 Significant differences were observed in the murine microbiome composition based on maternal CD status (online supplementary figure S5. Interestingly, exposure of GFM to the microbiome of pregnant CD donors at T3 resulted in disruptions of critical homeostatic elements of the immune system with a lower proportion of FoxP3+TREG, and IgA-expressing class-switched memory B cells in the colonic LP, and a reduction of FoxP3+TREG also in MLN compared with the exposure to control pregnant women’s stool (figure 4). Furthermore, inoculation with the stool from 3-month-old babies born to mothers with CD also led to a lower proportion of class-switched memory B cells in the colonic LP and MLN, and homeostatic IgA+ B cells in the colonic LP. TREG have a well-established role in the suppression of systemic and mucosal immune activation and in the regulation of intestinal inflammation.43 Similarly, IgA, the predominant antibody at mucosal surfaces, is of critical importance to mucosal homeostasis. IgA is associated with downregulation of proinflammatory epitopes on commensal bacteria,44 secretion of a biofilm that favours the growth of commensals,45 direction of luminal bacteria to M cells,46 maturation of dendritic cells,47 production of interleukin-1048 and FcαRI-mediated suppression of immune responses.49 It has been suggested that patients with IBD have an impaired production of polymeric IgA at the mucosal surfaces,50 51 and a significant reduction in peripheral switched memory IgD-CD27+ B cells has been described in the blood of 1-year-old infants born to IBD mothers treated with combination therapy.52 Previous reports have shown that intestinal bacterial colonisation pattern may affect B cell maturation in humans and that early gut microbiota might promote this maturation.53 Together, reduced frequencies of TREG and IgA+ B cell subsets in the colon of GFM inoculated with IBD-associated stools may indicate reduced frequencies of two critical homeostatic elements of the mucosal immune system that are driven by IBD-associated dysbiosis.
Our data provide a potential link between early life exposures, microbiome and future risk of IBD, underscoring the potential consequences of the aberrant formation of early-life microbiome during the sensitive time window of the immune system development. Of note, low bacterial diversity in early life has been shown to precede development of diseases such as asthma54 or allergies in childhood.55 Considering the central role of gut microbiome in IBD pathogenesis, we speculate that the observed patterns may, at least in part, help to explain the residual familiar risk of IBD in the offspring beyond the established shared genetic risk.
The strengths of our study include a unique prospective cohort of pregnant women with and without IBD and their babies, with serial sample collection and exposure data. For the first time, we were able to investigate the role of the major maternal and early life exposures on the rapid changes in the developing gut microbiome. Our study’s limitations include inability to correct for all potential confounders, such as IBD type, disease location, activity, complications, medications and other IBD-related variables that may impact the microbiome. Small numbers of babies exposed to antibiotics and exclusive formula feeding prevented us from further assessing the impact and interaction of these environmental exposures, which are known to affect infants’ gut colonisation. Moreover, we did not have access to blood from infants, and therefore, we resorted to a validated model system (GFM) to anticipate the immune-related consequences of the imbalanced microbiome.56 Also, we are reporting on changes in gut microbiome and immunological profiles induced by CD-associated donor stools in healthy mice. Yet, the impact of CD-associated dysbiosis on susceptibility to colitis cannot be inferred, neither can the underlying pathophysiological mechanisms driving the observed effects be determined. Future studies are warranted to test our findings in murine models of mucosal inflammation. Finally, although we see clear and reproducible changes within the adaptive immune system, we could not identify similar changes in innate immune cells. Lack of sensitivity, absence of inflammation or changes within alternative cellular subsets are among the possible explanations.
In summary, using a prospective cohort, we detected a significant association of maternal IBD with changes in microbiome composition during pregnancy and in the offspring. Exposure to the microbiome of pregnant patients with CD or their babies led to the development of an imbalanced immune system lacking critical homeostatic elements in a humanised experimental model. Understanding the critical events shaping the microbiome of mothers with IBD and their offspring can ultimately help to develop new strategies aiming at disease prediction and prevention.
Acknowledgments
We would like to thank our study participants for their invaluable contribution to this project.
References
Footnotes
Contributors JT and JH contributed equally. JT, J-FC, IP, JH and BJ conceived and designed the study. CE, NN, LT, QM, EM, C-LC, AK, JG, PL, HL, JS, JC-D and MD managed subject recruitment, data and sample collection and sample processing. GB, IM, JF, LT and AS designed and conducted germ-free mice (GFM) experiments. AS, MU and SM designed and conducted immune profiling experiments. JH, JT, JCC, AS and RH generated and analysed data. JT, J-FC, IP, SM and JH wrote the manuscript. All authors read the manuscript and provided critical comments.
Funding This work was funded by the International Organization for the Study of Inflammatory Bowel Disease (to IP and J-FC.), The Crohn’s and Colitis Foundation (to IP, JT, JCC and J-FC), and the Kenneth Rainin Foundation (to IP, J-FC, JF and SM). We would like to express our gratitude for Dr Jixin Dai and Ms Yi Liu’s generosity in supporting the MECONIUM study.
Competing interests JT received lecture fees from Takeda and Abbvie. JJF is a consultant for Janssen Research & Development and a member of the Scientific Advisory Board of Vedanta Biosciences. JFC reports receiving research grants from AbbVie, Janssen Pharmaceuticals and Takeda; receiving payment for lectures from AbbVie, Amgen, Allergan, Inc. Ferring Pharmaceuticals, Shire, and Takeda; receiving consulting fees from AbbVie, Amgen, Arena Pharmaceuticals, Boehringer Ingelheim, Celgene Corporation, Celltrion, Eli Lilly, Enterome, Ferring Pharmaceuticals, Genentech, Janssen Pharmaceuticals, Landos, Ipsen, Medimmune, Merck,Novartis, Pfizer, Shire, Takeda, Tigenix and holding stock options in Intestinal Biotech Development and Genfit.
Ethics approval All experiments were performed using protocols approved by the local animal ethics committee.
Provenance and peer review Not commissioned; externally peer reviewed.
Patient consent for publication Parental/guardian consent obtained