Article Text

Abstract
Objective We examined the impact of maternal low-dose aspartame and stevia consumption on adiposity, glucose tolerance, gut microbiota and mesolimbic pathway in obese dams and their offspring.
Design Following obesity induction, female Sprague-Dawley rats were allocated during pregnancy and lactation to: (1) high fat/sucrose diet (HFS) +water (obese-WTR); (2) HFS +aspartame (obese-APM; 5–7 mg/kg/day); (3) HFS +stevia (obese-STV; 2–3 mg/kg/day). Offspring were weaned onto control diet and water and followed until 18 weeks. Gut microbiota and metabolic outcomes were measured in dams and offspring. Cecal matter from offspring at weaning was used for faecal microbiota transplant (FMT) into germ-free (GF) mice.
Results Maternal APM and STV intake with a HFS diet increased body fat in offspring at weaning and body weight long-term with APM. Maternal APM/HFS consumption impaired glucose tolerance in male offspring at age 8 weeks and both APM and STV altered faecal microbiota in dams and offspring. Maternal obesity/HFS diet affected offspring adiposity and glucose tolerance more so than maternal LCS consumption at age 12 and 18 weeks. APM and STV altered expression of genes in the mesolimbic reward system that may promote consumption of a palatable diet. GF mice receiving an FMT from obese-APM and obese-STV offspring had greater weight gain and body fat and impaired glucose tolerance compared with obese-WTR.
Conclusion Maternal low-calorie sweetener consumption alongside HFS may disrupt weight regulation, glucose control and gut microbiota in dams and their offspring most notably in early life despite no direct low-calorie sweetener consumption by offspring.
- diet
- glucose metabolism
- intestinal bacteria
- obesity
- energy metabolism
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
Significance of this study
What is already known on this subject?
Maternal diet during pregnancy and lactation contributes to the developmental programming of long-term health and disease susceptibility in offspring.
Some evidence suggests that direct low-calorie sweetener consumption or exposure beginning in utero may promote weight gain and impair glucose tolerance and learning performance.
Low-calorie sweetener consumption can alter gut microbiota composition.
What are the new findings?
Independent of maternal obesity/high fat sucrose diet, maternal aspartame and stevia consumption increased adiposity in male and female offspring in early life and impaired glucose tolerance in male offspring, particularly with aspartame.
Consumption of low-calorie sweetener and HFS diet by obese pregnant and lactating rats altered gut microbiota of offspring causing (shown with faecal microbiota transplant) impaired adiposity and glucose tolerance in early life, despite no direct consumption of sweeteners by offspring.
Maternal aspartame and to a lesser extent stevia alongside HFS consumption altered expression of genes in the brain associated with feeding behaviour in offspring, which could promote consumption of palatable food if presented to offspring.
Significance of this study
How might it impact on clinical practice in the foreseeable future?
Perturbations to gut microbiota and metabolism during critical periods of development may increase the risk of metabolic disease later in life. Understanding the impact of dietary ingredients on maternal metabolism and gut microbiota may help to define optimal maternal diet, ultimately promoting a healthier future for both mother and child.
Introduction
The incidence of obesity has risen at an unprecedented rate in the past 30 years.1 Alongside the dramatic increase in global obesity rates, the number of women entering pregnancy with obesity and those with excessive gestational weight gain has also steadily risen.2 It is well established that maternal health and nutrition have a long-lasting impact on offspring health, and obesity and excessive weight gain during pregnancy are associated with poor pregnancy outcomes such as macrosomia, preterm delivery and greater obesity risk later in life.3
In response to higher obesity rates, consumption of low-calorie sweeteners (LCS) has risen, particularly in women and children.4 In 2007–2008, 15% of children and 32.9% of female adults in the USA consumed products containing LCS.4 In the Danish National Birth Cohort, almost half of women with gestational diabetes consumed LCS beverages during pregnancy with 9.3% consuming them daily. Daily consumption was positively associated with large birth size and overweight/obesity at 7 years in their offspring.5 LCS consumption in young females (<10 years) has been associated with early menarche—a known risk factor for chronic diseases.6 7 In addition, the presence of LCS has been detected in breastmilk, in particular saccharin, sucralose and acesulfame potassium, presenting a potential mode of transmission from mother to infant for some but not all LCS.8 9 Aspartame, a common artificial LCS, and stevia, a natural LCS extracted from a plant native to South America, are ~200 and 200–400 times sweeter than sucrose, respectively.10 Stevia is gaining popularity as a sweetener, and was historically used in Paraguay and Brazil to treat diabetes.11
Although LCS reduce the energy content of food, recent research suggests these sweeteners may promote weight gain.12 These findings may have particular relevance in a maternal population, since LCS are considered safe to consume during pregnancy and lactation.8 13 Mechanisms underlying the relationship between metabolic perturbations and LCS consumption are still emerging; however alterations in the gut microbiota may be one link between LCS consumption and impaired glucose tolerance.14 15 Communication between the gut microbiota and the brain along the microbiota-gut-brain axis is also implicated in regulating central appetite and food reward signalling16 and may be altered by LCS. In particular, the mesolimbic reward system is a likely contributor to obesity, as it regulates hedonic feeding mechanisms, which may override metabolically driven homeostatic feeding, potentially leading to overconsumption of food and weight gain.17
Archibald et al 18 reviewed the evidence for early life LCS exposure and the developmental origins of childhood obesity from human and rodent studies and concluded that while some observational studies suggest children consuming LCS have increased obesity risk, fewer studies have examined prenatal LCS exposure. In rats, increased body weight, fasting glucose and impaired learning performance were seen in males exposed to high doses of aspartame (50 mg/kg) beginning in utero and continuing with chronic direct postnatal exposure19; however, this remains controversial.20 Recently, Olivier-Van Stichelen and colleagues found that prenatal exposure to acesulfame potassium and sucralose altered offspring microbiota at 20 days of age in mice.9 Therefore, our objective was to examine whether maternal consumption of low-dose aspartame or stevia during pregnancy and lactation alters glucose tolerance and gut microbiota in pregnant and lactating dams. Further, we determined whether offspring metabolic health (ie, adiposity, insulin sensitivity, gut microbiota) was impacted by maternal LCS consumption despite no direct exposure, and if changes to offspring gut microbiota were causal to impaired metabolic outcomes. With a growing number of women entering pregnancy with overweight or obesity and the high rates of LCS usage in this population, we examined an obese maternal rat model given that LCS consumption in lean rats has been examined.9 19 21 22
Material and methods
Animals
150 female Sprague-Dawley rats (Charles River) aged 8 weeks were housed in a temperature and humidity-controlled room with a 12-hour light/dark cycle. Rats underwent an obesity induction phase for 10 weeks where they consumed a high fat/high sucrose diet (HFS; Dyets #102412) ad libitum.23 The top tertile of weight gainers was allocated to one of three groups (n=15/group) throughout pregnancy and lactation: (1) HFS +Water (obese-WTR) (obese control group); (2) HFS +aspartame (obese-APM) (5–7 mg/kg; Fluka); 3) HFS +stevia (obese-STV) (2–3 mg/kg; Rebaudioside A, Sigma-Aldrich). A lean reference group (lean-reference, n=15) was included as a benchmark of normal pregnancy and offspring outcomes. Sweeteners were administered in drinking water (changed weekly) and dose adjusted based on body weight and fluid intake. Female rats were bred with male Sprague-Dawley rats and at birth, litters were culled to 10 offspring (n=5M and 5F) to minimise confounding due to variable litter size. Cross-fostering within treatments of similar age was used to increase small litter size to 10. Offspring were weaned onto control diet (AIN-93; Dyets) and water at age 3 weeks and followed until age 18 weeks. In our analysis, all offspring within the same litter were considered as n=1.
Maternal and offspring body weight was measured weekly. Body composition was measured using dual-energy X-ray absorptiometry (DXA) (Hologic, Inc.)24 in offspring at 3, 12 and 18 weeks and dams at weaning. Dams’ food and fluid intake was measured prior to mating and in the first and third week of gestation and lactation for five consecutive days by weighing food cups and fluid bottles. Offspring food intake was measured at age 6, 12 and 18 weeks for five consecutive days. Oral glucose tolerance tests (OGTT) and insulin tolerance tests (ITT) were performed in dams (gestation and lactation day 13–16) and in offspring at age 8 and 17 weeks. At the termination of the study (dams at weaning; offspring at 3, 12 and 18 weeks of age), rats were over-anaesthetised with isoflurane and aortic cut. Samples from the cecum and the brain were collected.
Detailed methods used for OGTT, ITT, short chain fatty acids (SCFA), mesolimbic reward pathway gene expression and faecal and cecal microbiota composition are available in online supplementary material.
Supplemental material
Faecal microbiota transplant
Cecal matter collected from male obese-APM, obese-STV and obese-WTR offspring at weaning and stored at −80°C was pooled and mixed with 5 mL of sterile phosphate buffered saline solution in an anaerobic chamber to create a homogeneous solution. 400 µl of the phosphate-buffered saline (PBS)-cecal matter solution was gavaged into male germ-free mice age 8–10 weeks (n=10/group). Body weight was measured at baseline, 7, 10 and 14 days following gavage. Fifteen days post-gavage an OGTT was performed and the next day mice underwent a DXA scan and were euthanised.
Statistical analysis
All data are presented as mean±SEM. One-way analysis of variance (ANOVA) with Tukey post hoc was used to determine differences between groups. For outcomes with repeated measures, a two-way ANOVA was used with time as within-subject factor and maternal diet as between-subject factor. Data from the lean reference group are displayed for key metabolic outcomes as a reference point between a lean/healthy pregnancy and the obese phenotype. Given that there was no intervention and the lean-reference group were used for reference purposes alone, they were not included in the statistical analysis. The ‘n’ of experimental groups is given in online supplementary table 1. Analysis was completed using SPSS V.24.0 software (SPSS Inc.). Data were considered significant at p≤0.05. Statistical analysis for 16S rRNA Illumina sequencing was performed as previously described.25
Supplemental material
Results
LCS consumption increases adiposity in offspring in early life
In dams, no difference in body weight (figure 1A) or body composition (table 1) was observed.
Anthropometrics of dams at weaning*

Dam and offspring body weight. (A) Dam body weight during gestation and lactation. (B) Offspring birth weight. (C) Male offspring weight gain from weaning. (D) Female offspring weight gain from weaning. Values are mean±SEM. a,b superscripts indicate significant differences between groups where labelled means without a common superscript letter differ, p≤0.05 (ie, ‘a’ and ‘b’ differ; ‘ab’ does not differ from ‘a’ or ‘b’). Obese-APM, obese aspartame (n=10 dams gestation, n=9 dams lactation, n=9 M and 8 F offspring); obese-STV, obese stevia (n=8 dams gestation, n=7 dams lactation, n=7 M & F offspring); obese-WTR, obese control (n=13 dams gestation, n=6 dams lactation, n=6 M and F offspring); lean-reference, lean control (n=14 dams gestation, n=11 dams lactation, n=10 M and F offspring; not included in statistical analysis). G, gestation; L, lactation; W, week.
Stevia consumption reduced the fertility of dams, but all dams that became pregnant had a successful birth (findings previously published26). There were no differences in pup birth weight between groups (figure 1B). At weaning, there was a trend (p=0.066) for higher body weight in obese-APM versus obese-WTR male offspring (table 2) whereas female obese-APM and obese-STV offspring were significantly heavier than obese-WTR (table 3). To account for differences in weaning weight, we calculated weight gain from weaning for all subsequent time points and showed that obese-APM continued to show greater weight gain even when taking into account their higher starting point at 3 weeks of age. Male obese-APM offspring had greater weight gain than obese-WTR at 6 and 12 weeks (figure 1C) and female obese-APM had greater weight gain than obese-WTR and obese-STV from 12 to 18 weeks. Similar patterns were seen with absolute body weight although female obese-APM were consistently heavier than obese-WTR and obese-STV from 6 to 15 weeks and heavier than obese-STV at 18 weeks (online supplementary figure 1A,B).
Anthropometrics of male offspring at weaning, week 12 and week 18*
Anthropometrics of female offspring at weaning, week 12 and week 18*
Independent of the effects of maternal obesity/HFS diet, aspartame and stevia increased body fat percentage in male (table 2) and female (table 3) offspring at weaning. By 12 and 18 weeks, obese-APM, obese-STV and obese-WTR offspring had similar fat mass, suggesting an early impact of maternal LCS consumption on body composition that is overtaken by the long-lasting impact of maternal obesity/HFS diet. No differences in energy and fluid intake in dams or energy intake in offspring were seen (online supplementary figure 2) although male obese-WTR offspring consumed 50 and 43 kcal/day more, respectively, than obese-APM and obese-STV at age 6 weeks (females consumed 19 and 31 kcal/day more, respectively). Despite apparent lower energy intake, obese-APM male offspring had greater weight gain at age 6 weeks (figure 1C).
LCS consumption impairs insulin sensitivity in dams and maternal aspartame consumption reduces glucose tolerance/insulin sensitivity in male offspring in early life
Independent of the effect of obesity/HFS, obese-APM dams had reduced insulin sensitivity during gestation, evident by greater blood glucose levels at 60, 90 and 120 min following insulin load compared with obese-WTR (figure 2A). Obese-STV had higher blood glucose levels than obese-WTR at 120 min following insulin load (figure 2A). No differences in insulin sensitivity were seen during lactation (figure 2B). Obese-APM male offspring had reduced insulin sensitivity in early life. During the ITT at eight weeks, male obese-APM offspring had higher blood glucose levels at 60 and 120 min compared with obese-WTR (figure 2C). This was accompanied by higher fasting insulin in male obese-APM (5.3±0.3) and obese-STV (5.1±0.3) offspring compared with obese-WTR (4.9±0.07). By 17 weeks, obesity/HFS exerted more of an effect than LCS (figure 2D) and a significant diet effect indicated that maternal stevia consumption may in fact improve insulin sensitivity in male offspring at age 17 weeks. Insulin sensitivity in female offspring at 8 weeks did not differ (figure 2E) but similar to males, maternal stevia consumption showed an improvement in insulin sensitivity at 17 weeks (figure 2F).

Insulin sensitivity of dams and offspring. (A) Dam glucose response during ITT at gestation. (B) Dam glucose response during ITT at lactation. (C) Male glucose response during ITT at age 8 weeks. (D) Male glucose response during ITT at age 17 weeks. (E) Female glucose response during ITT at age 8 weeks. (F) Female glucose response during ITT at age 17 weeks. Values are mean±SEM. a,b superscripts indicate significant differences between groups where labelled means without a common superscript letter differ, p≤0.05. Obese-APM, obese aspartame (n=10 dams gestation, n=9 dams lactation, n=9 M and 8 F offspring); obese-STV, obese stevia (n=8 dams gestation, n=7 dams lactation, n=7 M and F offspring); obese-WTR, obese control (n=13 dams gestation, n=6 dams lactation, n=6 M and F offspring). ITT, insulin tolerance tests.
Glycaemia during OGTT in dams at gestation and lactation (figure 3A–D) was not independently affected by LCS. However, male obese-APM and obese-STV offspring had higher glucose at 0 and 15 min during the OGTT compared with obese-WTR at age 8 weeks (figure 3E). Glucose AUC was also greater in obese-APM males compared with obese-WTR (figure 3F). By 17 weeks, the influence of maternal obesity/HFS on glycaemia (figure 3G) and glucose AUC (figure 3H) became greater than LCS with only glucose at 30 min being significantly higher in obese-APM versus obese-WTR (figure 3G). There were no independent LCS effects on female offspring glycaemia and glucose AUC at 8 weeks (figure 3I,J). Maternal obese-APM consumption appears to have a sex-specific impact on offspring glucose tolerance and insulin sensitivity in early life which is overtaken by a long-lasting effect of maternal obesity/HFS on glucose tolerance in both sexes during adulthood (figure 3K,L).

Glucose tolerance of dams and offspring. (A) Dam glucose response during OGTT at gestation. (B) Dam AUC from gestation OGTT. (C) Dam glucose response during OGTT at lactation. (D) Dam AUC from lactation OGTT. (E) Male offspring glucose response during OGTT at age 8 weeks. (F) Male AUC from OGTT at age 8 weeks. (G) Male offspring glucose response during OGTT at age 17 weeks. (H) Male AUC from OGTT at age 17 weeks. (I) Female offspring glucose response during OGTT at age 8 weeks. (J) Female OGTT AUC at age 8 weeks. (K) Female offspring glucose response during OGTT at age 17 weeks. (L) Female AUC from OGTT at age 17 weeks. Values are mean±SEM. a,b superscripts indicate significant differences between groups where labelled means without a common superscript letter differ, p≤0.05. ***superscript indicates p<0.0005. Obese-APM, obese aspartame (n=10 dams gestation, n=9 dams lactation, n=9 M and 8 F offspring); obese-STV, obese stevia (n=8 dams gestation, n=7 dams lactation, n=7 M and F offspring); obese-WTR, obese control (n=13 dams gestation, n=6 dams lactation, n=6 M and F offspring); lean-reference, lean control (n=14 dams gestation, n=11 dams lactation, n=9 M and F offspring; not included in statistical analysis). AUC, area under the curve; OGTT, oral glucose tolerance tests.
Mesolimbic reward pathway is altered by maternal aspartame and stevia consumption
Maternal LCS or HFS diet exposure did not consistently alter mesolimbic reward pathway in dams (figure 4A,B), perhaps because dietary exposures in adulthood may not impact neural parameters to the same degree as early exposures.27 However, in male offspring, maternal obese-APM and obese-STV increased ventral tegmental area (VTA) dopamine transporter (DAT) mRNA levels at weaning (figure 4C) that persisted to 18 weeks (figure 4D). At age 18 weeks, male obese-APM and obese-STV offspring had greater nucleus accumbens (NAc) D2 receptor compared with obese-WTR (figure 4E). In female offspring, maternal obese-APM increased NAc Mu-opioid receptor levels at weaning compared with all other groups (figure 4F). Female obese-APM and obese-STV offspring had increased levels of NAc D2 receptor compared with obese-WTR at weaning (figure 4G). Obese-WTR and obese-APM female offspring had higher VTA tyrosine hydroxylase (TH) mRNA levels compared with obese-STV at weaning (figure 4H). At age 18 weeks, female obese-APM offspring had greater NAc DAT levels compared with obese-WTR (figure 4I). Taken together, these data indicate that maternal LCS exposure can induce long-lasting effects on mesolimbic dopamine circuitry.

Mesolimbic reward gene expression in dams and offspring. mRNA levels of (A) dam nucleus accumbens (NAc) tyrosine hydroxylase (TH) at end of lactation, (B) dam ventral tegmental area (VTA) mu-opioid receptor at end of lactation. (C) Male offspring VTA dopamine transporter (DAT) at weaning. (D) Male offspring VTA DAT at age 18 weeks. (E) Male offspring NAc D2 receptor at age 18 weeks. (F) Female offspring NAc mu-opioid receptor at weaning. (G) Female offspring NAc D2 receptor at weaning. (H) Female offspring VTA TH at weaning. (I) Female offspring NAc DAT at age 18 weeks. Values are mean±SEM. Superscripts indicate significant difference between groups; *p<0.05; **p<0.01. Obese-APM, obese aspartame (n=7 dams, n=9 M and 8 F offspring); obese-STV, obese stevia (n=7 dams, n=7 M and F offspring); obese-WTR, obese control (n=6 dams, n=5 M and F offspring).
Faecal microbiota is impacted by LCS consumption in dams and early life in offspring
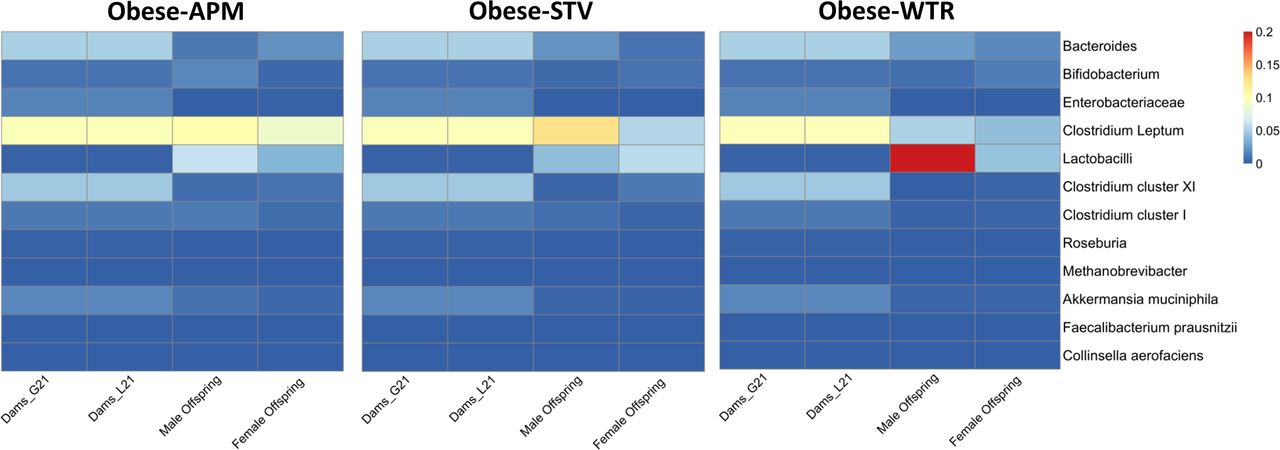
We used qPCR to quantify changes over time in select bacteria that have been shown to be relevant to obesity (eg, Bifidobacterium, shown to be low in obesity and negatively correlated with serum lipopolysaccharide28 29 and Enterobacteriaceae, a proinflammatory pathobiont30). In dams and offspring, we detected time-dependent variations in bacterial abundance (online supplementary figures 3–5) that likely reflect the effects of the LCS imposed on the normal variation that occurs in dams transitioning from non-pregnant to pregnant and then lactating status and in offspring with increasing age. In offspring, microbial differences according to maternal diet appear more robust in early life (week 6) and tend to converge to a more common signature by 18 weeks. As a result we plotted qPCR relative abundance in a heat map depicting dam microbiota at G21 and L21 and offspring microbiota at week 6 (figure 5).

Heat map of the relative abundance of select taxa (quantified with qPCR) in faecal matter of dams at gestation D21 and lactation D21 and male and female offspring at 6 weeks of age. Obese-APM, obese aspartame (n=10 dams gestation, n=9 dams lactation, n=9 M and 8 F offspring); obese-STV, obese stevia (n=8 dams gestation, n=7 dams lactation, n=7 M and F offspring); obese-WTR, obese control (n=13 dams gestation, n=6 dams lactation, n=6 M and F offspring).
The heat map depicts a similar higher relative abundance of Clostridium leptum in obese-APM and obese-STV dams and offspring. Obese-WTR dams also had higher Clostridium leptum, but this pattern of higher abundance was not seen in their offspring. Although the relative abundance of many of the other taxa was similar between dams and offspring, dams had higher abundance of Akkermansia muciniphila and Enterobacteriaceae than offspring and Lactobacilli tended to show sex-specific variation in offspring.
Maternal LCS consumption impacted offspring cecal microbiota
Cecal microbiota composition of dams and offspring at 18 weeks at the family level is shown in online supplementary figure 6. No differences in alpha diversity were detected with the exception of obese-STV dams, which showed a potential rescuing of reduced diversity resulting from an obesogenic diet (online supplementary figure 7). Only trends towards differences in beta diversity in dams (p=0.06) or their offspring (males p=0.17; females p=0.08) were detected (online supplementary figure 7). Linear discriminant analysis effect size (LEfSe) analysis showed that Obese-APM dams had reduced abundance of Enterococcaceae, Enterococcus, Parasutterella and increased abundance of Clostridium cluster IV compared with obese-WTR dams (figure 6A). Porphyromonadaceae was overabundant in obese-STV dams and male and female obese-APM and obese-STV offspring compared with obese-WTR (figure 6B,C–F). Sporobacter was altered in obese-STV dams and male offspring (figure 6B,D).

Linear discriminant analysis effect size describing the greatest differences between treatment communities. Maternal low-calorie sweetener treatment impacted cecal microbiota composition in dams at the end of lactation/weaning and offspring at 18 weeks. Graphs represent histogram of linear discriminant analysis scores between: (A) APM and WTR dams; (B) STV and WTR dams; (C) male APM and WTR offspring; (D) male STV and WTR offspring; (E) female APM and WTR offspring; (F) female STV and WTR offspring. Obese-APM, obese aspartame (n=7 dams, n=9 M and 8 F offspring); obese-STV, obese stevia (n=7 dams, n=7 M and F offspring); obese-WTR, obese control (n=5 dams, n=5 M and F offspring).
Gut microbiota composition at weaning directly influences weight gain and glucose intolerance
Germ-free mice that received cecal microbiota transplant from offspring of aspartame or stevia consuming dams (now conventionalised, CONV) had significantly greater body weight 7, 10 and 14 days following inoculation compared with CONV-WTR mice (figure 7A). CONV-APM mice had significantly greater glucose values 0, 30, 90 and 120 min and CONV-STV mice had greater glucose values 90 and 120 min following a glucose load compared with CONV-WTR mice (figure 7B). CONV-APM and CONV-STV mice had significantly greater glucose AUC indicating reduced glucose tolerance (figure 7C). CONV-APM and CONV-STV mice displayed increased fat mass (figure 7D) and percent body fat (figure 7E) with no difference in lean mass (figure 7F) compared with CONV-WTR. Microbiota composition of mice at 14 days is shown in online supplementary figure 8 at the family level and as community differences with LEfSe analysis which show a consistent increased relative abundance of Porphyromonadaceae in LCS-recipient mice similar to that seen in APM and STV offspring and dams (except APM dams; figure 6).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Faecal microbiota transplant from obese-APM and obese-STV offspring significantly increased body weight, glycaemia, fat mass and percentage of body fat without impacting lean mass. (A) Body weight. (B) Blood glucose concentrations during oral glucose tolerance test. (C) Glucose AUC from the oral glucose tolerance test. (D) Fat mass. (E) Percent fat mass. (F) Lean mass. Values are mean±SEM. a,b Superscripts indicate significant differences between groups where labelled means without a common superscript letter differ, p≤0.05. * indicates p<0.05; ** indicates p<0.01; *** indicates p<0.0005. CONV-APM, conventionalised aspartame (n=9); CONV-STV, conventionalised stevia (n=10); CONV-WTR, conventionalised water control (n=9). AUC, area under curve.
Maternal aspartame consumption altered SCFA production in dams
Maternal LCS consumption significantly increased cecal propionate, butyrate and isobutyrate concentrations in obese-APM and obese-STV dams compared with obese-WTR (online supplementary figure 9A–D). Obese-APM also had higher isovalerate and valerate concentration than obese-WTR dams (online supplementary figure 9E,F). No significant differences were seen in offspring (data not shown).
Discussion
The results of our study assessing maternal consumption of aspartame and stevia are consistent with emerging human evidence from observational studies that LCS intake is associated with increased body mass index (BMI) and cardiometabolic risk.31 We present evidence that maternal aspartame and stevia consumption during pregnancy and lactation perturb insulin sensitivity in dams during gestation, increase adiposity in male and female offspring at weaning and aspartame impairs insulin and glucose tolerance in male offspring. In addition, maternal LCS consumption alters maternal and offspring gut microbiota, maternal cecal SCFA concentrations and offspring expression of genes within the mesolimbic reward system. When cecal matter from weanling obese-APM and obese-STV offspring was transplanted into germ-free mice, the aberrant phenotype was transferred, suggesting that early life changes in gut microbiota composition from indirect exposure to LCS mediate adverse outcomes. Importantly, our findings reveal that perturbations are transmitted to offspring despite no direct LCS exposure.
Early safety work concluded that aspartame poses no risk to mother and fetus in terms of toxicity if consumed within adequate daily intake (ADI) levels.32 While these early studies examined short-term effects on toxicity and fetal neurodevelopment at doses that exceeded the ADI, recent studies have examined metabolic consequences of nutritionally relevant doses.15 33 In mice, lifetime exposure to aspartame beginning in utero in lean mothers resulted in increased body weight and fasting glucose and reduced insulin sensitivity.19 21 Consistent with sex differences seen in the developmental programming literature, changes were significant in males, with trends in females.21 Similarly, we found that maternal aspartame consumption had greater impact on male offspring glucose control than female. This is not surprising since male fetuses are more vulnerable to intrauterine insults and adverse perinatal conditions, like diet and stress.34
It is well established that the gut microbiota is an important contributor to health and plays a causal role in development of an obese phenotype in rodents.23 35 Suez et al 15 found that mice exposed to saccharin for 11 weeks exhibited glucose intolerance alongside microbiota changes and that microbiota transplant from saccharin treated mice to germ-free mice produced marked glucose intolerance. Similarly, Palmnäs et al 33 showed distinct changes in Clostridium leptum and Enterobacteriaceae in adult male rats consuming aspartame. When we inoculated germ-free mice with cecal microbiota (CONV-D) from obese-APM and obese-STV offspring at weaning, they developed increased body weight, impaired glucose tolerance and greater fat mass compared with CONV-D obese-WTR mice. These results suggest that the gut microbiota at weaning played a causal role in mediating adverse glucocentric and body composition outcomes. Similarly, Suez and colleagues demonstrated that saccharin consumption could alter gut microbiota composition that directly mediated glucose intolerance.15 However, this study is the first to show that indirect exposure to LCS during gestation and lactation is sufficient to impact gut microbiota composition in offspring and impair glucocentric parameters. While gut microbiota is known to be vertically transmitted from dams to offspring during birth,36 we cannot rule out that horizontal transfer via coprophagia might also participate in microbiome transfer from dams to offspring.
Increased Porphyromonadaceae has been observed in a non-alcoholic steatohepatitis mouse model alongside aberrant glucose tolerance and weight gain,37 and in high-fat diet (HFD)-induced glucagon-like peptide (GLP-1) resistant mice.38 We observed an increase in Porphyromonadaceae in obese-STV dams and male and female obese-APM and obese-STV offspring, which may provide further evidence linking Porphyromonadaceae growth to a greater tendency for development of the metabolic syndrome.37 In our faecal microbiota transplant (FMT) study, Porphyromonadaceae was 209% higher in CONV-APM and 176% higher in CONV-STV mice compared with CONV-WTR. We also observed an alteration in genera Sporobacter in obese-STV dams and obese-STV male offspring compared with obese-WTR dams, which is enriched in type-2 diabetes patients,39 further implicating the potential role of microbiota in mediating metabolic changes following LCS exposure. Although taxonomical differences between dams and offspring exist (likely in part due to offspring consuming a low-fat diet (AIN93) and obese dams consuming an HFS diet), the persistence of some bacteria such as Porphyromonadaceae across generations and in the germ-free mice suggests that certain taxa may play an important role in the metabolic derangements.
Currently, it is hypothesised that aspartame may impact gut microbiota by altering intestinal alkaline phosphatase (IAP) activity. IAP is a duodenal brush border enzyme that improves insulin sensitivity by dephosphorylating lipopolysaccharides, and is inactivated by phenylalanine, a by-product of aspartame metabolism. Mice consuming aspartame have reduced IAP activity alongside glucose intolerance.40 Others suggest that reduced IAP alters gut pH and thus gut microbial ecology,41 and therefore IAP may be a link between direct aspartame consumption and the altered microbiota observed in dams. Rebaudioside A, a compound of stevia commonly used in sweeteners, is converted to steviol in the colon by gut microbiota.42 43 Although steviol has shown little impact on total gut microbiota composition, a small growth of total aerobes and coliforms has been reported.43 Moreover, antimicrobial effects of stevia on gut microbiota (Lactobacillus reuteri) have been observed.44
SCFAs are microbial metabolites produced following fermentation of unabsorbed food stuff (eg, dietary fibre) by the gut microbiota.45 SCFAs provide ~10% of daily energy and numerous benefits to the host including energy for colonocytes (butyrate).45 While dietary supplementation with SCFA has been shown to have anti-obesity effects,46 the contrary presence of elevated faecal SCFA in obesity47 has raised the question of the precise role of SCFA in obesity. In agreement with Palmnäs et al,33 direct obese-APM consumption increased propionate concentration in dams compared with all groups. Interestingly, direct stevia consumption also increased cecal propionate concentration. Obese-APM and obese-STV dams also had higher cecal butyrate levels which may indicate greater ability to extract energy from food as observed in ob/ob mice35 or less efficient SCFA absorption. Our findings appear consistent with the higher faecal SCFA levels recently shown to be associated with gut dysbiosis, adiposity and cardiometabolic risk factors in humans.46 Our results suggest that gut microbiota metabolites may participate in the aberrant metabolic changes seen with direct LCS exposure.
The mesolimbic reward pathway contains dopaminergic neurons that originate in the VTA located in the midbrain, and project to forebrain structures including the NAc.48 Activation of the mesolimbic reward pathway promotes food-seeking behaviour, and maternal HFD has been shown to impact this pathway in dams and offspring.49 Obese-APM consumption in dams significantly increased VTA DAT mRNA in male weanlings that persisted to 18 weeks of age suggesting a long-lasting influence of maternal LCS on somatodendritic dopamine regulation in offspring. Ideally we would have performed a HFD challenge in our offspring to examine their preference for HFD given previous studies showing a relationship between maternal HFD consumption and offspring reward system.50 Furthermore, Labouesse et al showed that mice overexpressing D2 receptor (similar to our offspring) had reduce Ucp1 mRNA levels in brown adipose tissue and gained weight on a high fat but not a chow diet.51 Although we cannot rule out the possibility that LCS exposure also reduced energy expenditure in our rats, analysis of white adipose tissue revealed no differences in genes related to energy expenditure in our study. Mu-opioid receptor upregulation has been linked to greater intake of palatable food, specifically food that is high in fat and may occur as a result of heightened responsiveness to endogenous and exogenous opioids in this region.52 In our LCS offspring, altered mesolimbic gene expression did not translate into greater food intake and may not have directly contributed to the observed metabolic phenotype. However, our results suggest that long-lasting changes in NAc mu-opioid receptor expression as a result of maternal diet during pregnancy/lactation may impact food preference of offspring in the absence of direct HFS diet and LCS exposure during their lifetime. Since central resistance to hyperleptinaemia commonly seen in obesity can occur not only in the hypothalamus but also in the mesolimbic pathway,53 it is possible that changes in the mesolimbic reward pathway in this study are due to the higher leptin levels seen in dams consuming LCS (data not shown).
Conclusion
In summary, we found that maternal LCS consumption impacted metabolic outcomes and the mesolimbic reward system in dams and offspring. Although obese-APM and obese-STV were similar in terms of detrimental effects on maternal insulin sensitivity during gestation and offspring adiposity at weaning, obese-APM had a more profound impact on worsening male glucose tolerance and consistently altering mesolimbic reward system gene expression. Furthermore, maternal LCS exposure altered gut microbiota composition in both dams and offspring with a notable increase in Porphyromonadaceae. Through our FMT study, we demonstrate that alterations in cecal microbiota composition from indirect LCS exposure in offspring at weaning drove metabolic impairment and adverse body composition in offspring in early life. Given that dams consumed LCS throughout pregnancy and lactation, we cannot decipher whether in utero or postnatal exposure is more important. Therefore, future studies should examine independent effects of gestation and lactation exposure to LCS (through cross-fostering) and use a pair-fed design to isolate the effects of the obesogenic diet. This study not only supports findings that LCS may not be metabolically inert, but that indirect exposure may also have long-lasting impact on offspring and development of central feeding circuits.
Acknowledgments
The authors would like to acknowledge Kristine Lee, Faculty of Kinesiology, and Dawn Martin, University of Calgary, for technical assistance; Shelly Wegener, Dr Richard Pon and Matthew Workentine, Centre for Health Genomics and Informatics at the University of Calgary for their technical assistance and support with the 16S rRNA sequencing and analysis; and Dr Lindsay Naef for teaching brain dissection and collection.
Footnotes
Contributors JEN designed research, carried out experiments, collected data, analysed data, generated figures and wrote paper. NAC, TK and LAJ helped carry out experiments. ACN and HRR helped analyse 16S rRNA sequencing data. JS and SLB provided input into research design. ENT, FC, CT and KDM helped perform the faecal microbiota transplant experiment. SM performed short chain fatty acids analysis. RAR designed research, wrote paper and had primary responsibility for final content. All authors read and approved the final manuscript.
Funding This work was supported by a Canadian Institutes of Health Research grant (MOP115076). JEN is supported by Alberta Children’s Hospital Research Institute and Canadian Institutes of Health Research. NAC was supported by a Talisman Energy Fund Healthy Living and Injury Prevention Studentship. TK is supported by an Alberta Innovates Health Solutions Doctoral Scholarship, Eye’s High Doctoral Scholarship and Vanier Canada Graduate Scholarship. LAJ was supported by an Alberta Children’s Hospital Research Institute Summer Studentship. FC is supported by a Faculty of Kinesiology Dean’s Doctoral Scholarship and Alberta Children’s Hospital Research Institute scholarship. SM is supported by an Alberta Innovates Postgraduate Fellowship and Eye’s High Postdoctoral Fellowship.
Competing interests None declared.
Patient consent for publication Not required.
Ethics approval The University of Calgary Animal Care Committee granted ethical approval for this study.
Provenance and peer review Not commissioned; externally peer reviewed.
Data availability statement Data are available upon reasonable request up until 3 years after publication.