Article Text

Abstract
Cirrhotic portal hypertension is characterised by development of the decompensating events of ascites, encephalopathy, portal hypertensive bleeding and hepatorenal syndrome, which arise in a setting of cirrhosis-associated immune dysfunction (CAID) and define morbidity and prognosis. CAID describes the dichotomous observations that systemic immune cells are primed and display an inflammatory phenotype, while failing to mount robust responses to pathogen challenge. Bacterial infections including spontaneous bacterial peritonitis are common complications of advanced chronic liver disease and can precipitate variceal haemorrhage, hepatorenal syndrome and acute-on-chronic liver failure; they frequently arise from gut-derived organisms and are closely linked with dysbiosis of the commensal intestinal microbiota in advanced chronic liver disease.
Here, we review the links between cirrhotic dysbiosis, intestinal barrier dysfunction and deficits of host-microbiome compartmentalisation and mucosal immune homoeostasis that occur in settings of advanced chronic liver disease. We discuss established and emerging therapeutic strategies targeted at restoring intestinal eubiosis, augmenting gut barrier function and ameliorating the mucosal and systemic immune deficits that characterise and define the course of decompensated cirrhosis.
- chronic liver disease
- cirrhosis
- gut immunology
- bacterial interactions
- bacterial translocation
Statistics from Altmetric.com
Key messages
The gut-liver-immune axis describes important bidirectional communications between the intestinal microbiome, liver and the mucosal immune system.
Host–microbiome interactions are essential to healthy metabolic function in addition to the maturation and maintenance of immune homoeostasis.
The intestinal barrier is critical for conserving normal physiology of the gut microbiome. Pathogenic bacterial adherence to mucosal surfaces and barrier damage is central in mucosal immune priming and may be important in the progression of liver disease.
Decompensated cirrhosis is associated with disruption of intestinal barrier dysfunction and bacterial translocation, which leads to endotoxaemia and systemic inflammation, driving the haemodynamic and immunological sequelae of advanced chronic liver disease.
Targeting the gut microbiome and restoring intestinal barrier function represents an attractive therapeutic target in reducing the risk and severity of decompensation in cirrhotic patients.
Burden of liver disease
There is an increasing burden of chronic liver disease worldwide with cirrhosis and the complications of viral hepatitis accounting for 2 million deaths per year, 3.5% of global mortality. Patients with compensated cirrhosis have a fivefold increase in mortality while those with decompensated cirrhosis are conferred a tenfold increased mortality risk when compared with the general population; median survival is 12 and 2 years, respectively.1 2 In addition to the growing burden of mortality, where cirrhosis and liver cancer now represent the 11th and 16th commonest causes of death worldwide, liver disease is associated with significant morbidity, healthcare resource utilisation and high economic impact.
The gut-liver axis
The gut-liver axis describes a unique relationship of metabolic, neuroendocrine and immune crosstalk between the gastrointestinal tract and hepatic sinusoids, connected in a bidirectional fashion through the portal circulation and biliary tree. The gut barrier is key to maintaining a homeostatic balance of tolerance to commensal organisms and foodborne antigens while mounting host-protective inflammatory reactions to pathobionts and pathogen-associated molecular patterns (PAMPs). To achieve this, the intestinal barrier is composed of a chemical (antimicrobial peptides, secretory immunoglobulin A and mucus layer), physical (epithelial cell tight junctions and microbiota due to colonisation resistance) and immunological barrier (gut-associated lymphoid tissue, GALT), (figure 1). Maintenance of a healthy intestinal barrier is essential to controlling interactions between the host and the microbiome; should these intestinal barriers fail and microbial translocation occur, the liver is the final fail safe to prevent systemic inflammation.
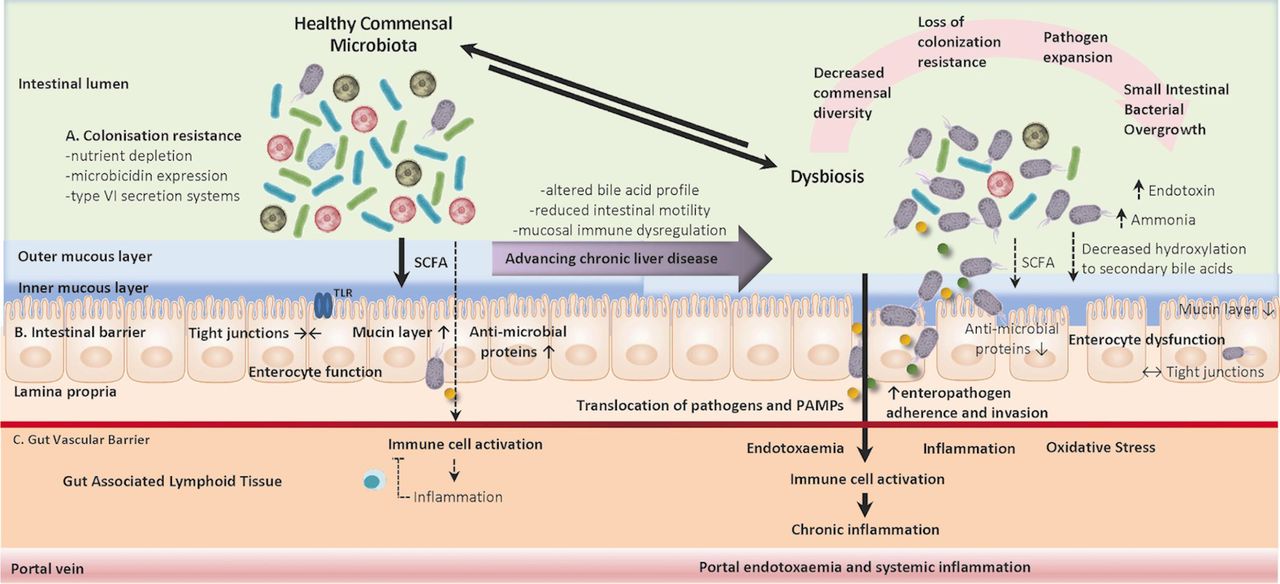
Intestinal barrier dysfunction in cirrhosis: In health, our intestinal immune system is resistant to colonisation by enteric pathogens through a tiered approach to barrier resistance that is supported by commensal micro-organisms at multiple levels. The dysregulation of commensal microbiota and host immune symbiotic interactions leads to the development of chronic inflammatory disorders spanning a diverse array of autoimmune, allergic and metabolic syndromes. Here, we outline some mechanisms of mutual regulatory interactions between host and commensal flora in maintaining compartmentalised and homeostatic host-microbiome interactions. (A) Colonisation resistance: competition for nutrients, the occupation of metabolic niches, the production of microbicidal proteins and targeted release of proteinaceous toxins through type VI excretion systems prevent enteropathogens (B) Host intestinal barrier function: maintenance of the intestinal epithelial barrier and intestinal tight junction proteins regulate paracellular and transcellular trafficking of pathobionts. Furthermore, the epithelial barrier is protected by the mucin layer which provides a physical barrier and is impregnated with microbicidal proteins limiting contact between epithelial cells and foreign antigens. (C) Gut vascular barrier: the gut vascular barrier provides a further firewall of compartmentalisation and defence from micro-organisms that have crossed the intestinal barrier into the lamina propria. Progressive chronic liver disease is associated with increasing perturbations in the healthy gut microbiome which leads to reduced commensal diversity, loss of colonisation resistance, expansion of enteropathogenic species and disruption of the intestinal barriers of defence. Decompensated cirrhosis is associated with bacterial translocation of pathogens and pathogen-associated molecular patterns (PAMPs) which induce a state of chronic systemic inflammation and immune exhaustion rendering patients susceptible to infectious insults. SCFA, short chain fatty acid; TLR, Toll-like receptor.
Previously considered a pauci-immune organ with predominant roles in metabolism, energy storage and detoxification, the liver has increasingly been recognised to contain a diverse repertoire of immune cells and plays a key role in immunological homoeostasis. The liver receives a dual blood supply of portal venous and systemic blood, rich in nutrients and bacterially-derived products leading to a tonic level of antigenic stimulation through which a balance of inflammatory and tolerogenic mechanisms act to maintain hepatic and systemic immune homoeostasis.3
The role of the microbiome in maintaining gut health
The development of culture-independent techniques of 16S ribosomal RNA gene profiling and whole genome sequencing has revolutionised our understanding of the gut commensal microbiota in health and disease and led to the development of reference microbial genome libraries.4 The human commensal microbiota (comprising eukaryotes, bacteriae, archaea and protozoans) is estimated to contain 100 trillion (1014) microbes subdivided to 12 phyla and >2000 species with a compound genome, termed the gut microbiome, outnumbering our host genome 100-fold. The gut microbiome demonstrates temporal and spatial differences influenced by pH, luminal secretions, water absorption and intestinal motility. Predominant autochthonous phyla of Firmicutes, Bacteriodetes, Actinobacteriae and Proteobacteriae act in synergy with luminal enterocytes to meet our diverse metabolic, nutritional and immune requirements.5
Short chain fatty acids
Short chain fatty acids (SCFAs; butyric, acetic and propionic acid) are by-products of bacterial colonic carbohydrate fermentation which are actively transported into colonic enterocytes where they form substrates for mitochondrial citric acid cycling and cellular energy metabolism.6 SCFAs have important roles in reducing luminal pH, stimulating mucin production, intestinal motility and the maintenance of enterocyte viability and tight junction integrity.7 SCFAs also condition intestinal epithelial cells (IEC) to be more responsive to bacterial products, important for controlling the dichotomy between host responses to pathogens and tolerance to commensals.8 Furthermore, SCFAs regulate immune responses in the GALT; they inhibit macrophage and dendritic cell activation, inflammatory cytokine induction and have been shown to shape the T helper cell repertoire.9
Bile acids
Bile acid metabolism reflects one of the important circuits in the bidirectional interplay between the liver and gut microbiome. Bile acids exert selection pressure on the commensal gut bacterial population which in turn regulates the composition, deconjugation and recirculation/excretion of the luminal and hepatic bile acid pools through the expression of bile salt hydrolases. Hepatocyte cholesterol 7α-hydroxylase (CYP7A1) catalyses (in a rate-limiting fashion) the formation of primary bile acids (cholic acid and chenodeoxycholic acid) from cholesterol before their conjugation and biliary excretion. Conjugated bile salts serve primarily to emulsify ingested fats and fat-soluble vitamins prior to their efficient absorption and enterohepatic recirculation via ileal enterocytes. Regulation of bile acid synthesis (via classical and alternative pathways) is through negative feedback regulation of CYP7A1 under the control of farnesoid x receptor (FXR) signalling (leading to enterocyte transcriptional upregulation of fibroblast growth factor 19 (FGF19) which interacts with the receptors b-klotho and FGF receptor 4 expressed on hepatocytes).10 FXR signalling regulates cellular energy metabolism, maintains the intestinal epithelial barrier and the gut vascular barrier and has been shown in experimental settings to attenuate liver injury and tumorigenesis.11 Bile acids additionally regulate mucosal immune tolerance and control both innate immune inflammatory signalling and adaptive immunity by modulating the differentiation of Th17 and Treg cells, a process which becomes disrupted in cirrhosis.12 Reduced primary bile acid formation and excretion in chronic liver disease represents a likely mechanism driving negative selection of key autochthonous taxa towards the development of dysbiosis.13
Colonisation resistance and antimicrobial resistance
In addition to the production of bioactive metabolites, commensal gut microbiota confer resistance to colonisation by pathogens. There are various mechanisms through which this has been shown to occur, including direct suppression of colonisation through nutrient competition in addition to the production of bacteriocidins, larger antimicrobial peptides and type VI secretion systems.14 Furthermore, B cells in intestinal lymphoid follicles secrete immunoglobulin A which is delivered to the intestinal lumen via a process of transcytosis and can, in turn, shape the microbiota; the absence of intestinal lymphoid follicles is associated with significant alterations in the commensal bacterial composition.15 Perturbations of the autochthonous microbiota with the onset of dysbiosis underly a central mechanism of immune dysregulation and susceptibility to enteric pathobionts in chronic liver disease.16
Gut microbiota in intestinal immune development
It is important to recognise the importance of bacterial biodiversity in the development and regulation of the intestinal immune system. Germ-free mice demonstrate thinner intestinal walls, with a reduced mucin layer, small Peyer’s patches and a sparse immune cell population within the lamina propria.17 IEC and lymphoid cells express pattern recognition receptors (PRRs) including Toll-like receptors (TLRs) and NOD-like receptors. These recognise and bind conserved cognate bacterial antigens such as components of the peptidoglycan cell wall, lipopolysaccharide (LPS), flagellin and bacterial DNA CpG motifs.18 Binding of these ligands triggers a transcriptional signalling cascade leading to the release of cytokines, which are involved in the shaping of intestinal naïve T cell differentiation to effector T cells with diverse homoeostatic roles. Microbial metabolites of Bacteriodetes and Firmicutes phyla interact with the host immune system where they can direct lymphoid organogenesis and regulate the stimulation and differentiation of a striking range of T cell-effector populations.19 Colonisation of germ-free mice by segmented filamentous bacteria simultaneously induces Th1, Th2, Th17 and regulatory T cell responses and can drive the coordinated maturation of intestinal T cell effector functions (figure 2). Conversely, the immune system reciprocally shapes commensal bacterial structure through mechanisms that contribute to pathogenic colonisation resistance.20
Key messages
Bacterial colonisation of the gastrointestinal tract occurs in infancy and matures towards a healthy, stable and diverse intestinal microbiome.
Host-microbiome interactions are essential for host metabolism, vitamin synthesis, pathogen resistance and immune system development.
Gut-Liver-Immune homoeostatic interactions are in fragile balance and become dysregulated in chronic liver disease.
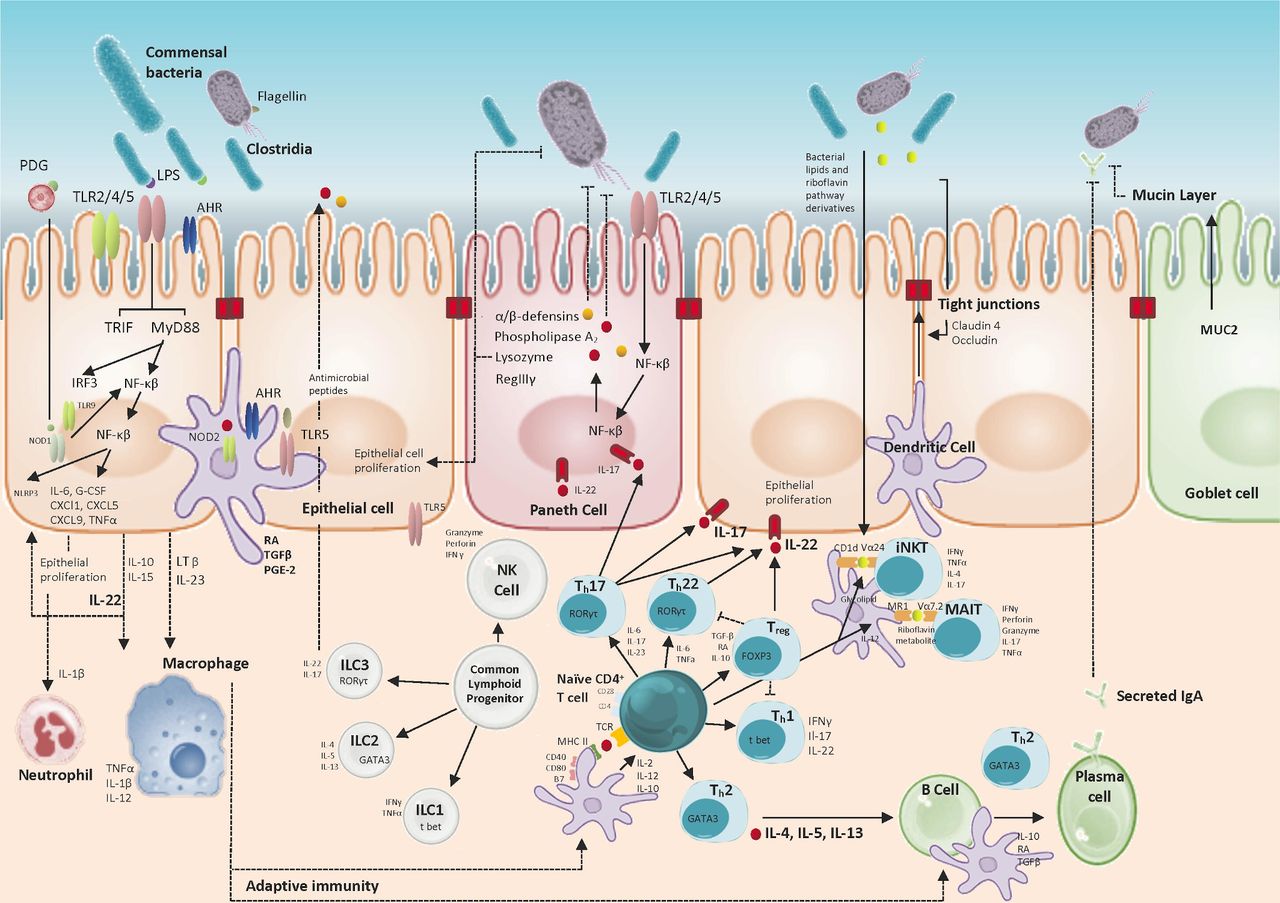
The role of the microbiota in maintaining homeostatic epithelial immune surveillance: Host–microbial interactions support mucosal barrier health in addition to maintaining mucosal immune responses; this enables synchronous tolerogenic and inflammatory responses to commensal and pathogenic organisms. Tolerance in part is reliant on spatial containment of commensal organisms; the intestinal epithelium represents a physical barrier separating gut luminal content from the intestinal immune cells populating the lamina propria and mesenteric lymphatics. Goblet and Paneth cells contribute to microbial colonisation resistance through the production of mucin proteins creating a physical barrier to bacterial interactions with the epithelium and through the production of microbicidal proteins (defensins, phospholipase A2, RegIIIγ) which penetrate the mucin layer in a concentration gradient. Expression of microbicidal proteins is stimulated by symbiotic commensal microbiota and the mucin layer is further bolstered by antigen-specific immunoglobulin A secreted from plasma cells. Intestinal epithelial cells (IECs) are exposed to a constant stream of antigens and act as sensors of the gut microbiome through a wide range of conserved pattern recognition receptor families (ie, NOD/TLR/RLR) and the aryl-hydrocarbon receptor (AHR) which mediate downstream signalling pathways that can promote epithelial barrier function. IECs produce interleukin-10 (IL-10) through Toll-like receptor 4 (TLR4) and AHR signalling which contributes to tolerogenic responsiveness to commensal bacteria and has important roles in autocrine stimulation of IEC proliferation and repair. Additionally, commensal stimulation of IEC promotes tolerogenic responses of innate myeloid populations (dendritic cells, macrophages) through the production of transforming growth factor beta (TGF-β), IL-10 and retinoic acid (RA) that subsequently shape tissue-resident lymphoid populations and promote the differentiation of antibody-producing plasma cells. IL-22 is highly expressed by Th17 cells and plays an important role in stabilising the intestinal barrier and maintaining compartmentalised and homeostatic control of gut microorganisms in addition to upregulating antioxidative genes in hepatocytes. Conversely, Th17 production of IL-17 is proinflammatory, profibrotic and associated with a tumourigenic adaptive immune response. Differential lymphoid populations of αβ-T cells, innate lymphoid cells and unconventional T cells are essential to maintaining the intestinal barrier in addition to host immune responses to bacterial, viral and parasite challenges through coordinated cytokine and cytotoxin release and chemotactic signalling. iNKT, invariant natural killer T; LPS, lipopolysaccharide; MAIT, Mucosa-associated invariant T; NOD, nucleotide-binding oligomerisation domain-like receptor; NRLP3, NOD-, LRR- and pyrin domain-containing protein 3; PDG, peptidoglycan; RLR, retinoic acid-inducible gene I receptor.
Gut-liver-immune axis in precirrhotic liver diseases
Non-alcoholic fatty liver disease (NAFLD) describes the excess accumulation of fat in the liver in the absence of a clear secondary cause (ie, alcohol consumption) and occurs in two main groups, NAFL and, in the presence of immune engagement, non-alcoholic steatohepatitis (NASH); the latter associated with a significantly increased risk of fibrosis, cirrhosis and hepatocellular carcinoma. NAFLD is characterised by increased hepatic lipid uptake, de novo lipogenesis and impairments in lipid export and fatty acid oxidation. NAFLD is rapidly emerging among the most prevalent chronic liver diseases and reflects increasing levels of obesity, metabolic disease and diabetes globally.
The intestinal microbiome has been implicated in the development of NAFLD, patients have bacterial overgrowth with reduced microbial diversity, an increased proportion of Firmicutes and a reduced proportion of Bacteriodetes that reverses with weight loss.21 A prospective study in patients with biopsy-proven NAFLD with early (stages 0–2) and advanced (stages 3–4) fibrosis showed differential abundance of the Firmicutes, Bacteriodetes and Proteobacteria phyla that predicted significant liver disease.22 Differences in the taxonomic composition of the intestinal microbiome in NAFLD change metabolic function; there is augmented dietary energy harvest and an increase in availability of bile acids, endogenous alcohols and volatile organic compounds. When paired with reduced SCFAs and choline this reduces the integrity of the intestinal barrier.23 Intestinal barrier dysfunction in NAFLD is an early manifestation of intestinal dysbiosis and is associated with portal translocation of pathobionts and endotoxins which propagates hepatic inflammation and metabolic dysfunction. Germ-free mice fed a high-fat diet accumulate less liver fat than conventionally housed mice,24 and hepatic steatosis can be induced after colonisation with intestinal microbiota from both obese mice25 and humans with NAFLD.26 Innate pathogen recognition receptors have been shown to propagate liver injury in NASH; TLR-4 mutant mice fed a methionine/choline deficient diet develop less hepatic steatosis and liver injury and TLR-9 knockout mice developed less steatohepatitis and fibrosis on a choline-deficient diet compared with wild-type mice.27
Chronic alcohol misuse and alcohol-related liver disease
Alcohol-related liver disease encompasses a spectrum of disease ranging from steatosis and steatohepatitis, to the development of fibrosis and acute alcoholic hepatitis, with or without cirrhosis. Chronic alcohol consumption induces lipogenesis through the induction of the sterol regulatory element-binding transcription factor 1 c in addition to impairing hepatocyte cellular energy metabolism and downregulating receptors that control enzymes involved in mitochondrial fatty acid oxidation (PPAR-α, PPAR-γ). Chronic alcohol-induced liver injury is mediated through the generation of reactive oxygen species, lipid peroxidation and glutathione depletion leading to hepatocyte cell death and the release of damage-associated molecular pathogens, which initiate a process of hepatic inflammation through innate PRRs.28 Alcohol and acetaldehyde, a toxic metabolite, damage the intestinal epithelial barrier, leading to altered expression of the tight-junction proteins zonula occludens-1 and claudin-1, increasing intestinal paracellular permeability and the portal translocation of bacterial and fungal peptides.29
Chronic alcohol misuse and the development of ARLD occurs in parallel with progressive changes in the intestinal microbiome.30 Early studies of ARLD demonstrated both bacterial overgrowth and endotoxaemia as a central mediator of TLR-4 activation and inflammation in alcoholic steatohepatitis that can be ameliorated by intestinal decontamination.31 Alcohol-related cirrhosis is associated with progressive dysbiosis, higher Enterobacteriaceae and lower Lachnospiraceae, Ruminococcaceae and Clostridiales XIV.32 Furthermore, there have been observed differences in the intestinal mycobiome with a reduction in the richness and diversity and a significant overgrowth of Candida spp in patients with chronic alcohol misuse irrespective of the stage of liver disease.33 Increased viral diversity was observed in faecal samples from patients with ARLD, with the most significant changes in samples from patients with alcoholic hepatitis. Specific viral taxa, such as Staphylococcus phages and Herpesviridae, were associated with increased disease severity and 90-day mortality in patients with alcoholic hepatitis.34
In acute severe alcoholic hepatitis dysbiosis is present with larger abundance of Bifidobacteria, Streptococci and Enterobacteria that correlates with the alcoholic hepatitis score. These changes in the intestinal microbiome are associated with alterations in the functional intestinal metabolomic signature with increased plasma bile acids, reduced faecal secondary bile acids and endotoxaemia.35 Transplantation of intestinal microbiota from humans with acute alcoholic hepatitis into germ-free and conventionally housed mice leads to increased intestinal permeability, increased hepatic T cells and higher levels of liver inflammation and necrosis which is ameliorated on subsequent second transfer of non-alcoholic hepatitis microbiota.36
Response to liver injury: regeneration, fibrosis and carcinogenesis
Signalling between the intestinal microbiome and the innate immune system via PRRs has been well documented to impact on the hepatic response to chronic injury and development of fibrosis.37 In healthy settings, the commensal microbiome plays a protective role in response to acute liver insult and reducing the progression of fibrosis. However, in settings of chronic liver injury hepatic fibrosis has been directly related to inflammatory signalling triggered by gram-negative endotoxin-producing intestinal microbes. Here, transforming growth factor-β-mediated signalling of hepatic stellate cells (HSC) by Kupffer cells occurs through an innate TLR4-MyD88-NFκB mechanism in response to LPS challenge.38 Antibiotic administration attenuates HSC activation and liver fibrosis in an experimental hepatic fibrosis model and is associated with increased intestinal tight junction protein expression and reduced LPS translocation.39 Furthermore, normalisation of the intestinal microbiome in animal models of diet-induced NASH has been shown to directly reverse portal hypertension.40
These mechanisms of intestinal dysbiosis, barrier dysfunction and translocation of PAMPs not only drive the propagation of hepatocellular inflammation and fibrosis but also lead to cumulative DNA damage and a platform for carcinogenesis.41 LPS-TLR-4 signalling has been implicated in the propagation of hepatocellular carcinogenesis, TLR-4 knock out mice display an 80–90% reduction in hepatocellular tumour number and size in a mouse model of chronic liver injury and fibrogenesis.42 These observations highlight the potential role for the gut-liver-immune axis as a target for the prevention and/or treatment of hepatocellular carcinoma in advanced stage liver disease.
Cirrhotic dysbiosis
The microbiome is profoundly altered in cirrhosis with a significant reduction in bacterial diversity and reductions in autochthonous microbial communities. Alterations in bile acid metabolism, intestinal motility and mucosal immunology all combine to lead to significant changes in the microbial composition of the gastrointestinal tract, which is compounded by widespread proton pump inhibitor usage and repeated antibiotic exposures. Small bowel bacterial overgrowth occurs in the context of delayed intestinal transit and further drives bacterial translocation. This cirrhotic dysbiosis is associated with the presence of ascites, spontaneous bacterial peritonitis and encephalopathy.43
Dysbiosis implies a loss of not only diversity with relative overexpression of pathological species but also loss of keystone taxa (important species that drive community composition, function and metabolism). Chen et al 44 described proportional reductions in the frequency of Bacteriodetes and Lachnospiracae seen in association with enrichment of Fusobacteria, Proteobacteria and Streptococaccae in cirrhotic patients. Qin et al 45 further reported reductions in Bacteriodes and Firmicutes phyla with higher levels of Proteobacteria and Fusobacteria. At the genus level buccal pathogens, including Veillonella, Streptococcus, Clostridium and Prevotella were enriched in cirrhotic patients suggesting oral colonisation of the intestinal microbial community in advanced chronic liver disease. A semiquantitative ratio of ‘good’ versus ‘bad’ bacteria, termed the cirrhosis dysbiosis ratio, has been shown to correlate with MELD score, the presence of decompensation and endotoxin levels.32
Intestinal barrier dysfunction in cirrhosis
Gut barrier disruption is well recognised in cirrhosis with increased interenterocyte spacing, shortening and widening of the microvilli, oedema of the lamina propria, fibromuscular proliferation and a decreased villous/crypt ratio seen46; these structural changes are associated with reductions in enterocyte expression of the tight-junction proteins occludin and claudin-1.47 Additionally, compromised antimicrobial host defence allows bacterial colonisation of the inner mucous layer of the intestine and predisposes to bacterial translocation in experimental cirrhosis.48 Dysbiotic pathogens such as Enterobacteriaceae use pathogenic virulence factors to become mucosally adherent and modulate the local immune response.49
B lymphocytes within the GALT play an important role in luminal IgA secretion and the maintenance of epithelial integrity, protection against gut pathogens (through selective IgA coating) and maintenance of intestinal bacterial homoeostasis through the secretion of commensal-specific IgA.50 51 The B-cell compartment is expanded in the intestinal epithelium and lamina propria in cirrhosis and has been putatively linked to increased portal bacterial translocation; further research is required to determine their functional role in this context.52 Paneth cells have recently been found to promote angiogenesis and regulate portal hypertension in response to microbial signals via secretion of proangiogenic signalling molecules.53
Translocation is not solely limited to the epithelial gut interface; gut-vascular barrier endothelial integrity has been shown to control translocation of antigens into the portal circulation under the influence of plasmalemma vesicle-associated protein-1, dependent on wnt/β-catenin signalling pathways which can be dysregulated by intestinal pathogens, such as Salmonella enterica serovar Typhimurium.54 Increasing permeability to bacteria and PAMPs secondary to multifaceted impairment of intestinal barrier function leads to bacterial translocation to mesenteric lymph nodes and extraintestinal sites (figure 1). Portal blood from cirrhotic patients has a composition similar to colonic mucosal biopsies, with marked dysbiosis and heightened Proteobacteria in particular Enterobacteriaceae 55; this intestinal translocation is directly associated with portal inflammation.56 Restoring the gut barrier is an important therapeutic target in liver disease.
Cirrhosis-associated immune dysfunction
The role of the intestinal innate immune system in providing the first line of defence against enteric pathogens is essential to healthy survival and there is an emerging wealth of data to report immune dysregulation and loss of bacterial compartmentalisation in the context of stable and decompensated cirrhosis. Not only are bacterial infections highly prevalent in patients with cirrhosis with over-representation of antimicrobial-resistant organisms, they are frequently severe and associated with acute decompensation of cirrhosis and acute-on-chronic liver failure (ACLF). The development of bacterial sepsis in cirrhosis confers a damning prognosis, with a 30% mortality at 1-month and 40% 1-year transplant-free survival.57
The frequency and severity of bacterial infections that complicates cirrhotic portal hypertension is attributable to a complex interplay between the gut, liver and immune system, driving a spectrum of systemic immunological disarray spanning the innate and humoral systems at multiple levels, which is broadly termed cirrhosis-associated immune dysfunction (CAID).58 CAID describes a paradigm where patients with advanced cirrhosis exist in a state of persistent low-level systemic inflammation with high baseline levels of proinflammatory cytokines in addition to impaired pathogen surveillance and predisposition to the development of infection (figure 3). Dysregulation of pleiotropic components of the reticuloendothelial, mucosal and systemic immune systems in cirrhosis are summarised in table 1. Superimposed sepsis aggravates systemic circulatory dysfunction and functional hypovolaemia associated with cirrhosis and is a common precipitant of decompensation and multiorgan dysfunction.59
Key messages
The intestinal barrier is critical for conserving normal intestinal physiology. Intestinal barrier dysfunction allows pathogenic bacterial adherence to mucosal surfaces and translocation of pathogen-associated molecular patterns, which drive fibrosis in chronic liver injury.
The gut microbiome is perturbed in cirrhosis with reduced diversity of autochthonous bacterial communities and relative over-expression of intestinal pathogens that progresses with advancing liver disease.
Cirrhotic dysbiosis is associated with intestinal barrier dysfunction, bacterial translocation and endotoxaemia, which drives systemic inflammation and cirrhosis-associated immune dysfunction.

{kind=link}
{kind=link}
{kind=link}
Mechanisms of cirrhosis-associated immune dysfunction: cirrhosis-associated immune dysfunction is a hallmark of advanced cirrhosis and patients are at risk of frequent and severe bacterial infections that often precipitate decompensation of chronic liver disease and acute-on-chronic liver failure. A plethora of innate and humoral immunological deficits are observed in the setting of advanced chronic liver disease, in part related to reduced synthesis of complement and opsonising proteins and in part secondary to sinusoidal capillarisation leading to altered vascular flow with porto-systemic shunting of portal blood. This is compounded by intestinal dysbiosis and disruption of the intestinal barrier creating a situation permissive to the translocation of pathogens and pathogen-associated molecular patterns (PAMPs) into the portal and systemic circulations. These stimuli activate pattern recognition receptors (PRR) on host immune cells and trigger the upregulation and release of cytokines and chemokines initiating an inflammatory response. Bacterial translocation and systemic endotoxaemia in advanced stages of decompensated cirrhosis leads to a state of chronic inflammation which compounds the haemodynamic and physiological sequelae of cirrhosis and impairs the ability to effectively respond to and clear pathogenic insults. FGF-19, fibroblast growth factor 19; FGFR4, fibroblast growth factor receptor 4; FXR, farnesoid x receptor; iNKT, invariant natural killer T; MAIT, Mucosa-associated invariant T; SCFA, short chain fatty acid.
Cirrhosis-associated innate and humoral immune deficits
The microbiome as a therapeutic target in chronic liver disease
Therapies that target dysbiosis and reduce bacterial translocation in the setting of cirrhosis and portal hypertension present attractive solutions to reduce the susceptibility to infection and improve clinical outcomes. This is particularly important considering the high prevalence of multidrug-resistant organisms identified in this population.60
Faecal microbial transplantation
The recognition of the role of dysbiosis as a driver of intestinal barrier dysfunction and bacterial translocation has led to the evaluation of FMT in models of cirrhotic and pre-cirrhotic dysbiosis (table 2). FMT has a beneficial impact on patients with hepatic encephalopathy with sustainable changes in commensal diversity and taxonomic representation in addition to improving cognitive performance and reducing hospitalisation rates. FMT improves indices of liver disease severity and 12-month survival (87.5% vs 33.3%) when compared with historical controls in patients with steroid-ineligible severe alcoholic hepatitis. FMT has been trialled in primary sclerosing cholangitis in single-centre settings and shown to be safe and associated with significant increases in taxonomical biodiversity in addition to improvements in alkaline phosphatase. FMT is being further evaluated through variable routes of administration in ongoing studies of cirrhosis, alcoholic hepatitis, NAFLD and chronic hepatitis B.
Published studies evaluating faecal microbial transplantation in cirrhotic and non-cirrhotic liver diseases
Potential pitfalls in FMT delivery on a wider scale remain pertaining to the compositional uncertainty of donor stool, risks relating to the potential for transmissible infectious diseases in addition to the microbiome’s wider effects on host-metabolism and immunological surveillance. Furthermore, there are practical considerations relating to the necessarily expansive nature of donor screening requirements, acceptability and uptake from patients on a wider scale, and safe and cost-effective methods of screening, storage and administration.
Antibiotics
Intestinal dysbiosis drives gut barrier dysfunction and shifts mucosal immune surveillance towards a state of inflammatory innate and humoral immune dysregulation that becomes more pronounced with advanced cirrhosis. Oral non-absorbable antibiotic administration reduces bacterial translocation and intestinal mucosal inflammation in a rat model of cirrhosis with reduced activation of B, T and NK cells in the intestinal epithelium and lamina propria.52 Fluroquinolone antibiotics have an established role in the prophylaxis of patients deemed to be at risk of spontaneous bacterial peritonitis (low-protein ascites or previous episode of peritonitis)61; fluoroquinolone use leads to a dendritic cell-mediated increase in peripheral Tregs and ameliorates the proinflammatory environment in cirrhosis.62 Rifaximin is a non-absorbable antibiotic with broad-spectrum antimicrobial activity against aerobic and anaerobic enteric microbes that is effective in treating recurrent hepatic encephalopathy.63 Furthermore, recent data suggest improvements in the myriad complications of advanced decompensated cirrhosis in patients awaiting liver transplantation.64 Further data are required prior to recommendations on expanded indications for the initiation of rifaximin therapy in decompensated cirrhosis. The benefits and duration of antimicrobial prophylaxis need to be weighed against concerns regarding the development of multidrug-resistant infections.65
Statins
Statins inhibit cholesterol synthesis through action on the rate-limiting enzymatic step hydroxymethylglutaryl-coenzyme A; beyond this they display hepatoprotective effects through pleiotropic mechanisms with demonstrable anti-inflammatory and anti-fibrotic effects.66 Statin therapy has been associated with a reduction in portal pressure and reduced mortality but not rebleeding in a multicentre randomised-controlled trial in the secondary prevention of variceal haemorrhage.67 Statins are being further trialled in combination with rifaximin in the LIVERHOPE-EFFICACY study, a phase III randomised controlled trial in the prevention of ACLF in decompensated cirrhosis (NCT NCT03780673).
Probiotics
Probiotics are live organisms which provide direct and indirect host benefits after ingestion and have been shown to favourably impact the production of SCFAs, gut barrier integrity, alter colonic pH and modulate the immune system.68 An elegant study by Macho Fernandez et al 69 demonstrated that the probiotic strain Lactobacillus salivarius Ls33 induced anti-inflammatory local dendritic cell responses (interleukin 10, IL-10 mediated) through an NOD2 signalling pathway that conferred in vivo protection in experimental models of colitis. Furthermore, distinctive strains of Lactobacillus and Bifidobacterium have shown protective effects in experimental models of colitis associated with an expansion of γδ-T cells and Treg cells occurring in the presence of reduced levels of tumor necrosis factor-α, MCP-1, IL-12 and interferon-γ and increased levels of IL-10.70 Probiotics have been demonstrated to have a beneficial impact on patients with cirrhosis and hepatic encephalopathy and are associated with improvements in symptom burden, blood ammonia levels and infection rates, although have not been shown to improve mortality in a meta-analysis.71 Larger clinical trials are required to confirm the clinical benefits of rationally designed probiotics in precirrhotic and cirrhotic liver diseases.
Prebiotics
Prebiotics are non-digestible food ingredients that exert effects on the intestinal microbiome with secondary beneficial effects on host homoeostasis.72 There is emerging evidence to support the role of early administration of prebiotic formula to infants in reducing allergic and infectious morbidity by indirect immunodevelopmental responses. Galacto-oligosaccharides improve intestinal barrier function by augmenting tight junction assembly and mitigate inflammatory responses to mycotoxin challenge.73 Lactulose is the best-known prebiotic in chronic liver disease and has a well-established role in the treatment of hepatic encephalopathy. Lactulose is metabolised by colonic bacteria and exerts positive selection pressure with expansion of bifidobacterial populations.74 In addition to this, lactulose acidifies the colonic microenvironment and leads to a reduction in the absorption of ammonium salts released as by-products of intestinal bacterial metabolism.
Short chain fatty acids
SCFAs are produced by bacterial fermentation of non-digestible polysaccharides. Butyrate is inversely correlated with the severity of liver disease and systemic inflammation in cirrhosis.75 Beta-hydroxy-beta-methylbutyrate supplementation is being trialled in cirrhosis (NCT03892070) and may have a role in particular in patients with sarcopenia associated with chronic liver disease. Further trials are needed to study the effect of SCFAs in liver disease.
Bacteriophage therapy
Bacteriophages are viruses that can infect and replicate within specific bacteria and allow selective editing of microbial communities with emerging roles in the targeting of drug-resistant bacteria. Bacteriophage treatment, directed against cytolysin-positive strains of Enterococcus faecalis (which is increased in alcoholic hepatitis and independently correlates with 90-day and 180-day survival), was shown to ameliorate liver injury, steatosis and inflammation in a humanised mouse model of ethanol-induced liver injury.76 Bacteriophage therapy represents a promising therapeutic modality for dysbiosis-associated conditions. Clinical trials in humans are now indicated to assess whether a phage approach would have utility in cirrhosis and further data are needed to present a robust and cohesive regulatory case for their clinical use. There remain obvious challenges ahead for phage therapy, particularly the narrow host range of most phages and development of resistance to phages that requires further research.
Carbon nanoparticles
Yaq-001 is a novel product composed of nanospheres of macroporous carbon that bind and prevent the translocation of bacterially derived molecules such as endotoxin and LPS occurring in settings of cirrhotic dysbiosis. In bile duct ligated rats Yaq-001 was associated with beneficial changes in the composition and function of the microbiome and improvement of circulating innate immune cell function. It is delivered in an oral sachet and is currently being evaluated in the multicentre CARBALIVE-SAFETY study in decompensated cirrhosis (NCT03202498).
FXR agonism
Obeticholic acid (OCA) is a potent FXR agonist which regulates bile acid metabolism in addition to impacting inflammatory, fibrotic and metabolic pathways. FXR-activation by OCA has been shown to induce a limited number of gram-positive facultative anaerobes which confer probiotic health benefits.77 OCA demonstrates effective biochemical responses in primary biliary cholangitis and is being evaluated in a phase III multicentre trial in NASH with improvements in fibrosis reported at the 18-month interim analysis.78 OCA has been shown to stabilise both the gut muco-epithelial barrier and gut vascular endothelial barrier reducing bacterial translocation in an experimental model of portal hypertension.79 FGF19 is a transcription factor under feedback regulation by bile acids binding to FXR which regulates bile acid synthesis and has effects on glucose and lipid metabolism. It represents an important target for metabolic and cholestatic liver diseases. M70 (an FGF19 analogue) has been shown to ameliorate bile acid toxicity and improve histological features of NASH with pleiotropic modes of action including anti-steatotic, anti-inflammatory and antifibrotic pathways in mouse models of NAFLD.80
Non-selective B-blockade
Non-selective B-blockers (NSBBs) are used in the treatment of portal hypertension and work to counteract increased cardiac output and reducing splanchnic blood flow. NSBBs have been shown to reduce intestinal permeability and markers of bacterial translocation (IL-6/LPS-binding protein).81 One mechanism of action is via improvements in intestinal hypomotility occurring in the context of sympathetic activation in cirrhosis which may reduce the development of small intestinal bacterial overgrowth and has been linked to reduced incidence of spontaneous bacterial peritonitis in cirrhosis.82
Albumin
Human albumin treatment has well-established indications in the setting of advanced cirrhosis, alongside vasopressors in type 1 hepatorenal syndrome, improving mortality in spontaneous bacterial peritonitis and in the prevention of postparacentesis circulatory dysfunction. In addition to improving the effective circulating volume in cirrhosis and preventing vasoconstrictor responses with activation of the renin–angiotensin system, albumin has further biological properties relating to endotoxin-binding and immunomodulatory actions at the vascular endothelium.83 Trials of long-term administration with albumin in patients with decompensated cirrhosis have shown conflicting results with respect to reductions in complications of cirrhosis and mortality, possibly attributable to differences in patient selection and albumin dosing regimens.84 85 Further trial data are awaited in this setting from the ATTIRE trial, a UK multicentre phase III trial (EudraCT 2014-002300-24) and the MICROB-PREDICT study evaluating microbiome-based markers to identify patients at risk of ACLF likely to benefit from albumin.
Immune signalling targets in cirrhosis
Targeting immune signatures that underpin intersections of perturbed host–microbiome interactions in cirrhosis represent an exciting and novel complementary strategy of therapeutic manipulation of the gut-liver-immune axis. LPS-TLR4 signalling is causally associated with the development of hepatocellular and systemic inflammation in both precirrhotic liver disease and cirrhosis. Both TLR4-mutant and TLR9-knockout mice are protected from the development of fibrosis after bile-duct ligation38 and blockade of the TLR4 receptor ameliorates LPS-induced ACLF failure in rat models of cholestatic cirrhosis.86 Recognition of the role of adaptive immunity and effector T cells in mucosal immune homoeostasis has led to evaluation of novel signalling targets in settings of chronic liver disease. Blockade of IL-17 promotes resolution of inflammation and fibrosis in animal models of both cholestatic and hepatocellular liver injury,87 while IL-22 transgenic mice reduce liver fibrosis and accelerate regeneration in a model of CCl4-induced chronic liver injury.88
Microrna therapy
MicroRNAs are non-coding regulators of gene expression, regulating cell function and homoeostasis, and their aberrant expression has been found in virtually all diseases. It is increasingly recognised that microRNAs are instrumental in intestinal pathologies, in particular, through their targeting of transcripts encoding proteins of the intestinal barrier and their regulators.89 Therefore, molecules targeting specific micro-RNAs that improve intestinal barrier function, for example, miR-320a inhibits Escherichia coli-induced damage to the intestinal barrier in ulcerative colitis may also reduce bacterial translocation in chronic liver disease.90
Key messages
Restoring intestinal eubiosis is a promising therapeutic strategy in patients with advanced chronic liver disease.
Prebiotics and non-absorbable antibiotics have established roles in the treatment of hepatic encephalopathy and the prevention of bacterial peritonitis.
Faecal microbial transplantation has been shown to reduce severity and frequency of episodes of recurrent hepatic encephalopathy, however, larger trial data on its safety and efficacy in decompensated cirrhosis and other settings of chronic liver disease is required.
Bacteriophage therapy represents a promising tool for the selective manipulation of microbial communities in settings of chronic liver disease.
Conclusions
The development of culture-free technologies has allowed for a rapid growth in our understanding of the homoeostatic roles of resident gut microbiota and their symbiotic interactions with the host. Reduced bacterial diversity with disruption of the balance of indigenous microbial communities in cirrhosis has led to a spotlight being shone on the gut microbiome, dysbiosis and intestinal barrier disruption as central disease mechanisms which make attractive targets for novel therapeutic development.
Acknowledgments
“The MICROB-PREDICT project has received funding from the European Union’s Horizon 2020 research and innovation programme under grant agreement No 825694. This reflects only the author's view, and the European Commission is not responsible for any use that may be made of the information it contains.”
References
Footnotes
Twitter @ThomasTranah, @DrLAEdwards, @DebbieShawcros1
Contributors THT wrote the first draft of the review and compiled the tables and figures. This was then extensively edited by LAE and DLS prior to BS making the final revisions. All authors have approved the final submitted version of the manuscript. DLS accepts overall responsibility for the final submitted version of this review.
Funding The authors have not declared a specific grant for this research from any funding agency in the public, commercial or not-for-profit sectors.
Competing interests BS has consulted for Ferring Research Institute, Intercept Pharmaceuticals, HOST Therabiomics and Patara Pharmaceuticals. BS’s institution UC San Diego has received grant support from BiomX, NGM Biopharmaceuticals, CymaBay Therapeutics, Synlogic Operating Company and Axial Biotherapeutics. DLS has consulted for Norgine, Shionogi, Mallinckrodt and Kaleido Biosciences. She has delivered paid lectures for Falk Pharma, Norgine and Alfa Sigma. DLS institution King’s College London has received grant support from Norgine and European Union’s Horizon 2020 research and innovation programme under grant agreement No 825694. THT and LAE have no conflicts to declare.
Patient and public involvement Patients and/or the public were not involved in the design, or conduct, or reporting, or dissemination plans of this research.
Provenance and peer review Commissioned; externally peer reviewed.