Article Text

Abstract
Objective Patients with increased PD-L1+ host cells in tumours are more potent to benefit from antiprogrammed death-1/programmed death ligand-1 (PD-L1) treatment, but the underlying mechanism is still unclear. We aim to elucidate the nature, regulation and functional relevance of PD-L1+ host cells in hepatocellular carcinoma (HCC).
Design A total of untreated 184 HCC patients was enrolled randomly. C57BL/6 mice are given injection of Hepa1-6 cells to form autologous hepatoma. ELISpot, flow cytometry and real-time PCR are applied to analyse the phenotypic characteristics of PD-L1+ cells isolated directly from HCC specimens paired with blood samples or generated from ex vivo and in vitro culture systems. Immunofluorescence and immunohistochemistry are performed to detect the presence of immune cells on paraffin-embedded and formalin-fixed samples. The underlying regulatory mechanisms of metabolic switching are assessed by both in vitro and in vivo studies.
Results We demonstrate that PD-L1+ host macrophages, which constructively represent the major cellular source of PD-L1 in HCC tumours, display an HLA-DRhighCD86high glycolytic phenotype, significantly produce antitumourigenic IL-12p70 and are polarised by intrinsic glycolytic metabolism. Mechanistically, a key glycolytic enzyme PKM2 triggered by hepatoma cell derived fibronectin 1, via a HIF-1α-dependent manner, concurrently controls the antitumourigenic properties and inflammation-mediated PD-L1 expression in glycolytic macrophages. Importantly, although increased PKM2+ glycolytic macrophages predict poor prognosis of patients, blocking PD-L1 on these cells eliminates PD-L1-dominant immunosuppression and liberates intrinsic antitumourigenic properties.
Conclusions Selectively modulating the ‘context’ of glycolytic macrophages in HCC tumours might restore their antitumourigenic properties and provide a precise strategy for anticancer therapy.
- macrophages
- glucose metabolism
- hepatoma
- cancer immunobiology
- immunotherapy
Data availability statement
All data relevant to the study are included in the article or uploaded as supplementary information.
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
Significance of this study
What is already known on this subject?
Hepatocellular carcinoma (HCC) is the fifth most common cancer worldwide, with an extremely poor prognosis.
Patients with increased PD-L1+ host cells in tumours are more potent to benefit from antiprogrammed death-1/programmed death ligand-1 (PD-L1) treatment.
Macrophages constitute a major component of the leucocyte infiltrate in hepatocellular carcinoma (HCC) tumour stroma.
Macrophage polarisation is closely linked to changes in the cellular metabolic programmes.
What are the new findings?
PD-L1+ host macrophages display an HLA-DRhighCD86high phenotype and significantly produce antitumourigenic IL-12p70 in HCC.
Intrinsic glycolytic metabolism shapes PD-L1+ macrophages with antitumourigenic properties in HCC.
PKM2/HIF-1α axis, elicited by hepatoma cell-derived fibronectin 1, triggers pluripotent polarisation of macrophages.
PD-L1 blockade eliminates PD-L1-dominant immunosuppression and liberates intrinsic antitumour capability of glycolytic macrophages.
How might it impact on clinical practice in the foreseeable future?
Increased PKM2+ glycolytic macrophages predict poor prognosis of patients, blocking PD-L1 on those cells liberates intrinsic antitumourigenic activties.
Selectively modulating the ‘context’ of glycolytic macrophages in HCC tumours might provide a precise strategy for anticancer therapy.
Introduction
Programmed death ligand-1 (PD-L1) is a promising therapeutic target in aggressive cancers.1–4 PD-L1 is chiefly detected in inflamed epithelial tissues, cancer cells or host stromal cells; its expression is assumed as a predictive biomarker of benefit from anti-PD-1/PD-L1 treatment.5–7 However, several clinical studies have emphasised that patients with increased PD-L1+ host cells are more likely to benefit from anti-PD-1/PD-L1 treatment than those with increased PD-L1+ cancer cells.8–10 At present, the nature, regulation and functions of PD-L1+ host cells in human cancers are indeed unclear.
Macrophages constitute a major component of the leucocyte infiltrate in tumour stroma.11 In response to environmental signals, these cells acquire special phenotypic characteristics that are associated with diverse functions.12 13 It is worth noting that polarisation of macrophages is closely related to the changes of cellular metabolic programme.14 15 In both mice and humans, glycolytic metabolism is involved in classical activation of macrophages (M1), whereas mitochondrial oxidative phosphorylation (OXPHOS) is restricted to alternative activation of macrophages (M2).16 Up until now, little is known about whether and how metabolic changes might occur and regulate the phenotypes and functions of macrophages in a tumour environment.
Hepatocellular carcinoma (HCC) is the fifth most common cancer worldwide, with an extremely poor prognosis.17 18 By using HCC as a model system, the present study shows that PD-L1+ host macrophages, which constructively represent the major cellular source of PD-L1 in HCC tumours, display an HLA-DRhighCD86high glycolytic phenotype, significantly produce antitumourigenic IL-12p70 and are polarised by intrinsic glycolytic metabolism. Mechanistically, a key glycolytic enzyme PKM2 triggered by hepatoma cell-derived fibronectin 1 (FN1), via a HIF-1α-dependent manner, concurrently controls the antitumourigenic properties and inflammation-mediated PD-L1 expression in glycolytic macrophages. More importantly, although increased PKM2+ glycolytic macrophages predict poor prognosis of patients, blocking PD-L1 on these cells eliminates PD-L1-dominant immunosuppression and liberates intrinsic antitumourigenic properties.
Materials and methods
Patients and specimens
Tissues and paired blood samples were obtained from patients who underwent curative resection at the First Affiliated Hospital of Nanjing Medical University (online supplemental table 1). None of the patients had received anticancer therapy before sampling, and those with concurrent autoimmune disease, HIV or syphilis were excluded. Ninety-one patients who had complete follow-up data were used for immunohistochemistry and immunofluorescence analysis and/or assessments of disease-free survival (cohort 1; online supplemental table 1). Paired fresh samples of blood and tumour tissues from 93 HCC patients who underwent surgical resection between December 2018 and December 2021 were used for isolating peripheral and tissue-infiltrating leukocytes (cohort 2; online supplemental table 1). Clinical stages were classified according to the guidelines of the International Union Against Cancer.
Supplemental material
Animals, cell lines, reagents and primers
Wild-type C57BL/6J mice (6–8 weeks old, female) were purchased from the Nanjing Biomedical Research Institute of Nanjing University. Human Liver cell (L02), HEK293T and Huh7 cell lines were obtained from the American Type Culture Collection. The antibodies used in fluorescence-activated cell sorting/analyses are shown in online supplemental table 2. The antibodies used in immunoblotting and immunohistochemistry/immunofluorescence are shown in online supplemental table 3. The primers used in real-time PCR are listed in online supplemental table 4, and all other reagents are summarised in online supplemental table 5 unless indicated in the text.
Statistical analysis
Results are presented as mean±SEM. No data have been excluded. All statistical tests were performed as two sided. For data normally distributed, we applied the Student’s t-test, and the non-parametric exact Wilcoxon signed-rank test was used to compare data not normally distributed. For multiple comparisons, an analysis of variance followed by Bonferroni’s correction was applied. R values were calculated based on the analysis of Pearson’s correlation. All statistical tests were performed with GraphPad Prism (V.6) software. Differences with p<0.05 were considered statistically significant.
Additional methods
Other methods are detailed in online supplementary materials and methods.
Results
Intrinsic glycolytic metabolism polarises PD-L1+ macrophages in human hepatoma
We used confocal microscopy to examine the expression pattern of PD-L1 in HCC specimens (n=10) and found that PD-L1 was expressed mainly by CD68+ macrophages and was only weakly expressed by other stromal and hepatoma cells (figure 1A). To further confirm the source of PD-L1, we prepared single cell suspensions from HCC tumour and paired non-tumoural liver and blood (n=9; online supplemental figure 1A,B). PD-L1 was weakly expressed on blood monocytes (figure 1B). Although the percentage of PD-L1+ macrophages (CD14+/dim cells) was also increased in peritumoural liver tissues, the intensity of PD-L1 expression was low (figure 1B). As expected, most tumour tissues contained a significantly greater proportion of PD-L1+/high macrophages (figure 1B). In contrast, PD-L1 was hardly or only weakly expressed by other stromal cells, including B cells, T cells, NK cells and neutrophils, as well as CD45– non-haematopoietic cells in HCC tumours (online supplemental figure 1C,D). Thus, host macrophages are major sources of PD-L1 in HCC tumours.
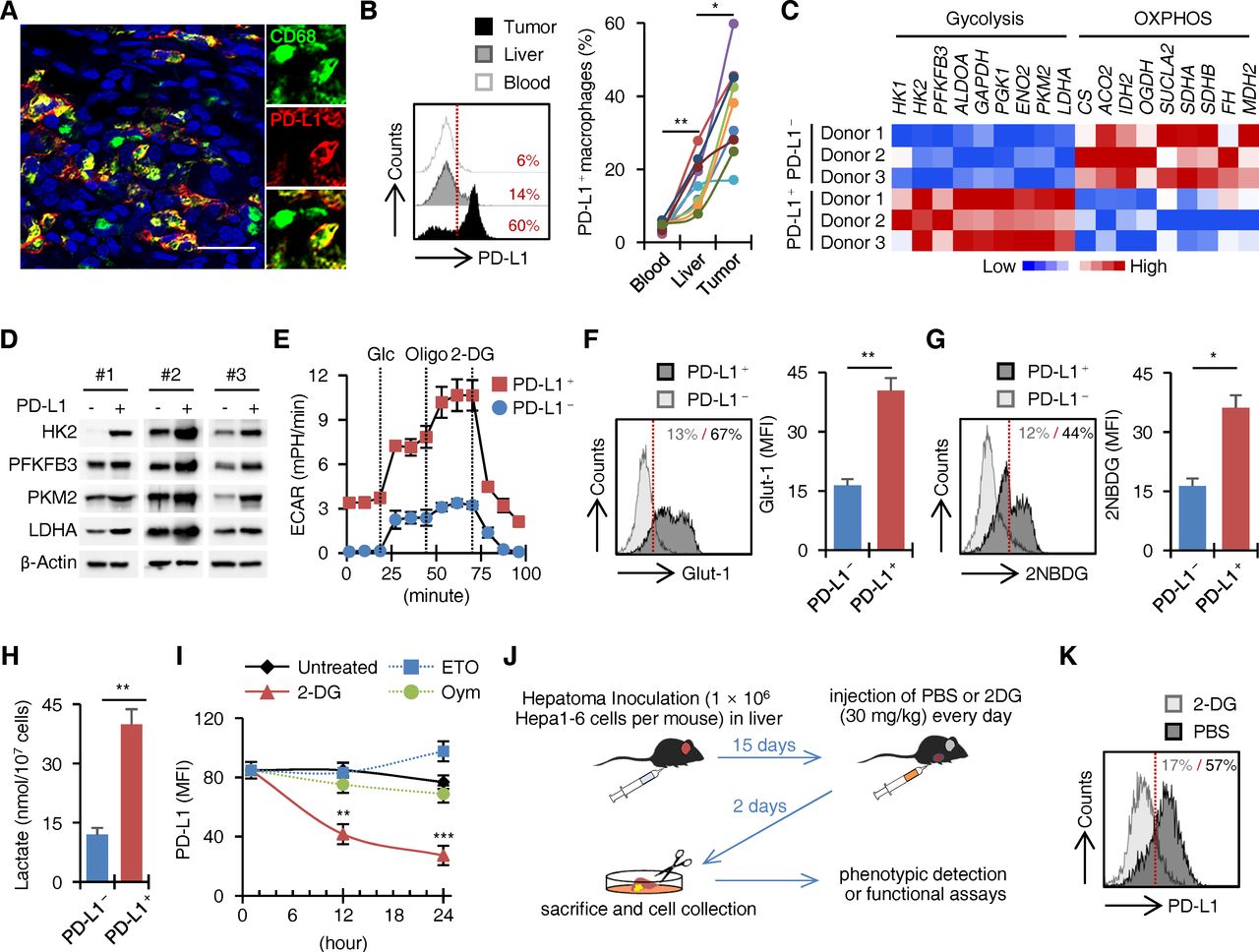
Glycolysis triggers PD-L1 on macrophages in human HCC tumours. (A) Confocal microscopy analysis of CD68+PD-L1+ cells in HCC tumours (n=10). Scale bar=100 µm. (B) FACS analysis of PD-L1 on monocytes/macrophages from HCC samples paired with blood samples (n=9). (C–H) Analysis of metabolic gene expression (C and D, n=3 for each), extracellular acidification rate (ECAR) (E, n=4), GLUT-1 expression (F, n=5), 2NBDG incorporation (G, n=5) and 20-hour lactate production (H, n=6) in PD-L1+ and PD-L1− macrophages purified from human HCC tumours. (I) Effects of metabolic inhibitors 2-deoxyglucose (2-DG), etomoxir (ETO) and oligomycin (Oym) on ex vivo PD-L1 expression in macrophages from human HCC tumours (n=5). (J and K) Mice bearing Hepa1-6 hepatoma for 15 days were injected with PBS or 2-DG intraperitoneally as described (J). PD-L1 expression on tumour macrophages was determined by FACS (K, n=6). *P<0.05, **p<0.01, ***p<0.001. HCC, hepatocellular carcinoma; PD-L1, programmed death ligand-1.
It is noteworthy that macrophage polarisation is closely linked to changes in the cellular metabolic programmes.14–16 To examine whether metabolic process determined macrophage PD-L1 expression, we applied real-time PCR to compare the transcriptional profiles of metabolic enzymes in PD-L1+ and PD-L1– macrophages from HCC tumours (n=3; online supplemental figure 1E). In general, PD-L1– macrophages chiefly expressed metabolic enzymes related to mitochondrial OXPHOS (figure 1C), revealing a metabolic quiescent state. Interestingly, PD-L1+ macrophages markedly expressed the key rate-limiting glycolytic enzymes (figure 1C,D and online supplemental figure 1F). We evaluated the extracellular acidification rate, which quantifies proton production as a surrogate for lactate production and thus reflects overall glycolytic flux.19 In support, PD-L1+ macrophages exhibited substantially higher basal and maximal glycolytic rates compared with PD-L1– macrophages (figure 1E). We also detected an obvious upregulation of glucose transporter Glut1 in PD-L1+ macrophages (figure 1F), suggesting an enhancement of glucose incorporation. Accordingly, PD-L1+ macrophages cultured ex vivo displayed a greater capacity to incorporate the fluorescent glucose analogue 2-(N-[7-nitrobenz-2-oxa-1,3-diazol-4-yl] amino)-2-deoxyglucose (2-NBDG) (figure 1G) and produced significantly more lactate (figure 1H).
The results described previously suggested that glycolytic metabolism may be involved in PD-L1 upregulation on tumour macrophages. To test this, we used specific inhibitors to reprogramme the metabolic processes of macrophages isolated from human HCC tumours ex vivo. Consistent with our hypothesis, a significant decline of PD-L1 expression in macrophages after exposing to 2-deoxyglucose (2-DG), a glucose analogue that inhibits glucose incorporation, were observed (figure 1I). By comparison, PD-L1 expression was marginally affected by incubating macrophages with etomoxir (ETO), a widely used small-molecule inhibitor of fatty acid oxidation,20 or with oligomycin, an inhibitor of ATP synthase (figure 1I).21 Furthermore, the roles of glycolytic metabolism in PD-L1+ macrophage induction were investigated in vivo (figure 1J). Consistently, intraperitoneal injection of 2-DG in Hepa1-6 hepatoma-bearing mice during the final 2 days of the experiment largely reduced the expression of PD-L1 in tumour macrophages (figure 1K).
PD-L1 blockade liberates antitumourigenic activity of glycolytic macrophages
We next asked whether inhibiting the glycolytic metabolism in tumour macrophages abrogated PD-L1-elicited immune privilege. To address that possibility, we purified PD-L1+ and PD-L1– macrophages, as well as autologous infiltrating T cells from HCC tumour tissues, and then performed ELISpot assays (online supplemental figure 2A). Tumour T cells cocultured with the PD-L1+ macrophages, but not the PD-L1– macrophages, exhibited an impaired production of IFN-γ (figure 2A). Consistent with our hypothesis, suppressing the PD-L1 expression by pre-exposing PD-L1+ macrophages to a glycolytic inhibitor 2-DG for 1 day (figure 1I) markedly restored the ability of tumour T cells to produce IFN-γ at levels comparable with that produced by untreated tumour T cells (figure 2A). Of note, we also blocked the PD-L1 by preincubation of PD-L1+ macrophages with the monoclonal antibody (mAb) MIH1 in parallel. Remarkably, this treatment eliminated the immune privilege induced by macrophage PD-L1 and significantly and additionally enhanced the production of IFN-γ by T cells (figure 2A). Therefore, these data suggest that PD-L1+ macrophages contain intrinsic antitumourigenic properties, which may be sustained by glycolytic metabolism but concealed by PD-L1 signals.
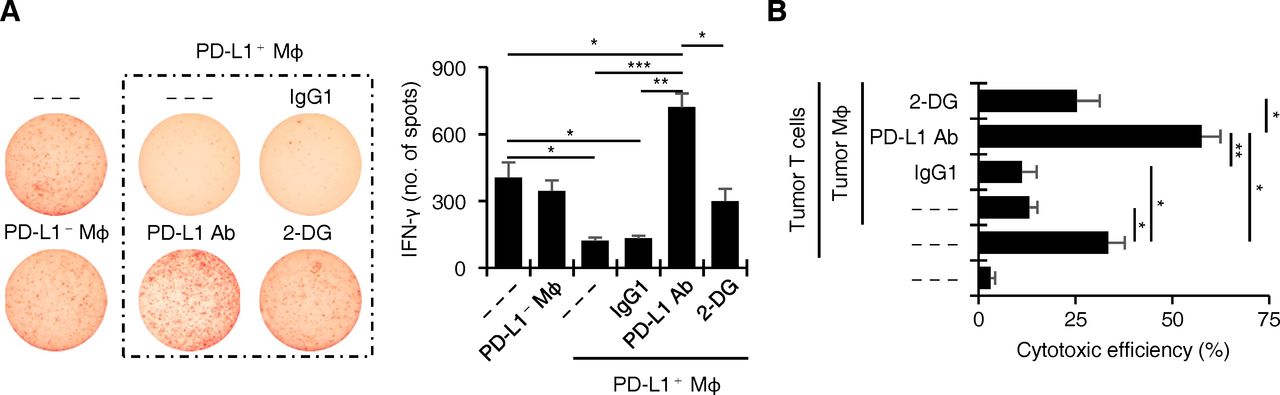
Glycolytic macrophages show intrinsic antitumourigenic properties. (A) IFN-γ detection by ELISpot in HCC tumour-derived T cells cultured alone or with PD-L1− or PD-L1+ macrophages (Mφ), or with those cells pre-treated with 2-DG, anti-PD-L1 antibody, or IgG1 as shown in online supplemental figure 2A (n=5). (B) cytotoxic effects of tumour T cells on CFSE-labelled autologous mouse Hepa1-6 hepatoma cells in the presence or absence of tumour Mφ that were left untreated or pre-treated with 2-DG, anti-PD-L1 antibody, or IgG1 as shown in online supplemental figure 2B (n=6). propidium iodide+ Hepa1-6 cells were measured by FACS. results are expressed as mean±SEM of three independent experiments. *p<0.05, **p<0.01, ***p<0.001.
We afterward established an ex vivo autologous cytotoxic system using tumour-specific T cells derived from Hepa1-6 hepatoma-bearing mice (online supplemental figure 2B). As expected, tumour T cells cultured ex vivo effectively killed autologous Hepa1-6 cells, and this process was hampered by addition of autologous F4/80+ tumour macrophages, but was further enhanced by adding those macrophages plus a specific antibody against murine PD-L1 (figure 2B). Supporting our hypothesis, tumour T cells cultured ex vivo with tumour macrophages that were pre-exposed to 2-DG only displayed the basal capabilities to kill autologous Hepa1-6 cells, but not an enhanced cytotoxic effect (figure 2B), suggesting that inhibition of glycolytic metabolism in macrophages also impairs their abilities to stimulate specific T cell response.
Glycolytic metabolism controls antitumourigenic properties and inflammation-elicited PD-L1 expression simultaneously in macrophages
Considering the intrinsic antitumourigenic properties of PD-L1+ macrophages in human HCC (figure 2), we next probed the activation status and cytokine production profile of these cells. Interestingly, PD-L1+ macrophages markedly expressed MHC class II molecule HLA-DR and costimulatory CD86, as compared with their PD-L1– counterparts (figure 3A); a marked correlation between the intensities of PD-L1 and HLA-DR was detected in macrophages isolated from HCC tumours (n=11) (figure 3B). The coexistence of HLA-DR and PD-L1 on macrophages was further confirmed by examining expression of these proteins in serial sections of HCC tumour samples (n=8). Most of the HLA-DRhighPD-L1+ CD68 cells were accumulated in the invading edge, which also represented the main site of CD8+ cytotoxic T cells (figure 3C). In line with these findings, we further demonstrated that PD-L1+ tumour macrophages, but not their PD-L1– counterparts, effectively expressed proinflammatory IL-1β, IL-6, and TNF-α, as well as the antitumourigenic IL-12p70 (figure 3D,E), a prominent inducer of Th1 responses in humans and in mice.22 23 Correspondingly, we demonstrated that, although the anti-PD-L1 Ab additionally enhanced the production of IFN-γ by tumour T cells in an ELISpot detection system, this process could be significantly abolished by additional neutralisation of IL-12p70 (online supplemental figure 3A,B). Analogously, IL-12p70 alone markedly increased the production of IFN-γ by tumour T cells ex vivo, which was considerably attenuated by adding the PD-L1+ HEK293T transfectants, but not by the control transfectants (online supplemental figure 3C,D).

PD-L1+ macrophages show a proinflammatory activated phenotype. (A) FACS analysis of CD86 and HLA-DR on PD-L1+ and PD-L1− macrophages from HCC tumours (n=11). (B) Correlation between PD-L1 and HLA-DR expression in macrophages from human HCC tumours (n=11). (C) Immunohistochemistry analysis of CD68, PD-L1, HLA-DR and CD8 expression in HCC tumours (n=8). (D and E) Analysis of transcriptional and translational levels of IL-1β, IL-6, IL-12 and TNF-α in PD-L1+ and PD-L1− macrophages from HCC tumours (n=7). (F–I) Analysis of glut-1 expression (F), 2NBDG incorporation (G), ECAR (H) and 20-hour lactate production (I) in HLA-DRhigh and HLA-DRlow macrophages purified from human HCC tumours (n=5). (J and K) Effects of 2-DG on CD86, HLA-DR, CD206, CD23 and CD163 expression (J), as well as cytokine production (K), in macrophages from HCC tumours (n=4). (L and M) Mice (n=6) bearing Hepa1-6 hepatoma were injected with PBS or 2-DG as described in figure 1J. CD86 and I-A/I-E expression (L), as well as cytokine expression (M), in tumour macrophages were determined by FACS and real-time PCR, respectively. Results are expressed as mean±SEM of at least three independent experiments. *P<0.05, **p<0.01, ***p<0.001. 2-DG, 2-deoxyglucose; ECAR, extracellular acidification rate; HCC, hepatocellular carcinoma; PD-L1, programmed death ligand-1.
Given that glycolytic metabolism was crucial for PD-L1+ macrophage induction (figure 1), we assessed whether such a mechanism was also responsible for macrophage activation. In fact, HLA-DRhigh macrophages from HCC tumour tissues were more potent in incorporating and utilising glucose (figure 3F,G), and these cells exhibited higher basal and maximal glycolytic rates and produced significantly more lactate, as compared with the HLA-DRlow/– macrophages (figure 3H,I). Accordingly, inhibition of glycolytic metabolism in macrophages from HCC tumour tissues by 2-DG effectively impaired the expression of HLA-DR and CD86, as well as the production of IL-1β, IL-12p70 and TNF-α (figure 3J,K). In contrast, such a treatment hardly affected the expression of macrophage M2 makers CD23 and CD206 but weakly increased the expression of CD163 (figure 3J). Similar results were obtained in Hepa1-6 hepatoma-bearing mice: suppression of glycolytic metabolism by 2-DG during the final 2 days of the experiment markedly attenuated the expression of IA-IE, CD86 and inflammatory cytokines in tumour macrophages (figure 3L,M).
We then investigated whether PD-L1 expression and proinflammatory cytokine production in macrophages were inter-related. Indeed, blocking the PD-L1 by a mAb in PD-L1+ macrophages did not affect the production of IL-12p70, as well as proinflammatory IL-1β, IL-6 and TNF-α (online supplemental figure 4A). Inversely, expression of PD-L1 in tumour macrophages was inhibited 40%–50% by blocking IL-1β and TNF-α but was not reduced by blocking IL-6 or IL-12p70 (figure 4A and online supplemental figure 4B). Comparably, recombinant IL-1β effectively induced upregulation of PD-L1 on blood monocytes. Although recombinant TNF-α alone had a marginal effect, it did synergistically increase the IL-1β-mediated PD-L1 expression (figure 4B). These findings indicate that IL-1β signalling contributes to PD-L1 induction in activated macrophages dominantly, which is consistent with a recent observation in liver cancer.24 Consistently, neutralisation of IL-1β plus TNF-α in Hepa1-6 hepatoma-bearing mice (figure 4C) successfully inhibited macrophage PD-L1 expression (figure 4D), suppressed tumour growth (figure 4E) and increased the infiltration of functional CD8+ T cells (figure 4F), but such treatment did not affect IL-12p70 expression (figure 4G). Therefore, glycolytic metabolism in macrophages simultaneously contributes to inflammation-mediated PD-L1 suppression and IL-12-dominant antitumourigenic activities.

Inflammatory cytokines induce PD-L1 in tumour macrophages. (A) Twenty-four-hour blockade of IL-1β or TNF-α reduced PD-L1 expression in macrophages from human HCC tumours (n=5). (B) Exposure of blood CD14+ cells to IL-1β and/or TNF-α for 24 hours led to PD-L1 upregulation (n=5). (C–G) Mice bearing Hepa1-6 hepatoma were injected intraperitoneally with PBS or anti-IL-1β antibody plus anti-TNF-α antibody as shown (C, each n=6). Tumour macrophage PD-L1 expression (D), tumour size (E), CD8+ cell infiltration and function (F) and tumour macrophage Il12a expression (G) were determined. Results are expressed as mean±SEM of at least three independent experiments. *P<0.05, **p<0.01, ***p<0.001. HCC, hepatocellular carcinoma; PD-L1, programmed death ligand-1.
Roles of hepatoma environments in glycolytic metabolism-elicited macrophage polarisation
To probe the mechanisms involved in glycolytic metabolism-elicited macrophage polarisation by a tumour environment, we first set out to establish conditions under which this process can be reliably reproduced in vitro. Human monocytes were incubated with supernatants from cultures of primary HCC cells (HCC-SN) or normal liver L02 cells (Liver-SN). Exposure to 20% HCC-SN, but not Liver-SN, for 24 hours did result in marked upregulation of the key rate-limiting glycolytic enzymes GLUT1, HK2, PFKFB3, PFKL, PKM2 and LDHA in monocytes, but returned to a normal level after 3 days (figure 5A,B). Consistent with this, compared with the monocytes incubated with HCC-SN for 3 days, the cells exposing to HCC-SN for 24 hours displayed at least threefold higher maximal glycolytic rates and produced significantly more lactate (figure 5C,D). These data imply that active glycolytic metabolism mainly occurs in the early differentiation stage of macrophages in a tumour environment. In line with this, we observed that the expression of HLA-DR and PD-L1 was significantly upregulated on monocytes after their exposure to HCC-SN for 24 hours but was reduced on day 3 (figure 5E,F). Similar patterns of cytokine productions were obtained in HCC-SN-exposed monocytes, including the accumulations of proinflammatory IL-1β and TNF-α, as well as antitumourigenic IL-12p70, in the culture media at their early differentiation stage and a subsequent decline on day 3 (figure 5G). Furthermore, in support of conclusion mentioned previously that glycolytic metabolism-mediated proinflammatory response triggers PD-L1 expression in macrophages (figure 4), using a competitive inhibitor 2-DG to block the glycolytic metabolism of HCC-SN-exposed monocytes at their early differentiation stage (figure 5H) effectively impaired the production of IL-1β, IL-12p70 and TNF-α (figure 5I) and suppressed HLA-DR and PD-L1 expression (figure 5J).

Hepatoma environments facilitate glycolysis and PD-L1 expression in macrophages. (A–G) Exposure of blood CD14+ cells to primary HCC-SN, but not liver-SN, led to marked increases of glycolytic metabolic enzymes (A and B), ECAR (C), lactate production (D), HLA-DR expression (E), PD-L1 expression (F) and inflammatory cytokine production (G) after 24 hours but returned to a normal level after 3 days (n=7 for each). (H–J) Treatment of 2-DG for 24 hours suppressed HCC-SN-mediated upregulation of lactate production (H), inflammatory cytokine production (I) and HLA-DR and PD-L1 expression (J) in CD14+ cells (n=7 for each). Results are expressed as mean±SEM of three independent experiments. *P<0.05, **p<0.01. 2-DG, 2-deoxyglucose; ECAR, extracellular acidification rate; HCC-SN, supernatants from cultures of primary HCC cells; PD-L1, programmed death ligand-1.
PKM2/HIF-1α axis is crucial for pluripotent polarisation of macrophages in HCC
We further explored the signalling pathways involved in glycolytic metabolism-elicited macrophage polarisation. Considering the finding that the key rate-limiting glycolytic enzymes HK2, PFKFB3 and PKM2 were markedly elevated in HCC-SN-exposed monocytes (figure 5B), we used specific inhibitors to abolish the activities of these proteins during at their early differentiation stage (figure 6A and online supplemental figure 5). The results showed that suppressing PKM2 activity using ML-265 markedly reduced the production of IL-1β, IL-12p70 and TNF-α, as well as the expression of HLA-DR and PD-L1, by HCC-SN-exposed monocytes, although inhibiting the activity of HK2 by 3-BP or PFKFB3 by 3-PO marginally affected (figure 6B–D). We further used siRNA to confirm the roles of PKM2 proteins in regulating pluripotent polarisation of macrophages (figure 6E) and proved that this treatment effectively impaired the inflamed activities, as well as HLA-DR and PD-L1 expression, in HCC-SN-exposed monocytes (figure 6F,G). Consistent with this in vitro observation, confocal microscopy demonstrated that, in human HCC tumour tissues, PKM2 was selectively and highly expressed by PD-L1+ macrophages and was only weakly expressed by the PD-L1– stromal and hepatoma cells (figure 6H).

Glycolysis upregulates PD-L1 on macrophages via PKM2/HIF-1α axis-dependent manner. (A–D) Effects of 3-BP (an hK2 inhibitor), 3-PO (a PFKEB3 inhibitor) or ML-265 (a PKM2 inhibitor) (A) on inflammatory cytokine production (B), HLA-DR expression (C) and PD-L1 expression (D) by CD14+ cells exposing to primary HCC-SN for 24 hours (n=5). (E–G) Transfection of siPKM2 (E) suppressed cytokine production (F), as well as HLA-DR and PD-L1 expression (G), by CD14+ cells exposing to primary HCC-SN for 24 hours (n=5). (H) Confocal microscopy analysis of CD68, PKM2 and PD-L1 distribution in HCC tumours (n=5). Scale bar=100 µm. (I) Transfection of siPKM2 suppressed STAT3 activation and HIF-1α expression by CD14+ cells exposing to primary HCC-SN (n=5). (J and K) Effects of STAT3 and HIF-1α inhibition on HLA-DR and PD-L1 expression (J), as well as inflammatory cytokine production (K), by CD14+ cells exposing to primary HCC-SN for 24 hours (n=5). (L and M) Immunohistochemical analysis of PKM2+CD68+ cell distribution in HCC tumours (L, n=91). Patients were further divided into two groups according to the median value of the PKM2+CD68+ cell density in the tumour regions (less PKM2high macrophages, ≤90 cells (n=46); more PKM2high macrophages, >90 cells (n=45)). The disease-free survival rate of these patients was analysed with the Kaplan-Meier method and log-rank test (M). Scale bar=100 µm. Results are expressed as mean±SEM of at least four experiments. *P<0.05, **p<0.01. HCC, hepatocellular carcinoma; HCC-SN, supernatants from cultures of primary HCC cells; PD-L1, programmed death ligand-1.
It is noteworthy that, besides serving as a key rate-limiting glycolytic enzyme, PKM2 can translocate into the nucleus to maintain HIF-1α stabilisation and STAT3 phosphorylation.25–27 Indeed, silencing PKM2 expression in HCC-SN-exposed monocytes largely suppressed the activities of both HIF-1α and STAT3 (figure 6I). In support, suppressing the stabilisation of HIF-1α by inhibitor α-ketoglutarate significantly antagonised pluripotent polarisation of tumour macrophages (figure 6J,K), and the effect was comparable with that of PKM2 inhibition. In contrast, abolishing the activation of the STAT3 signal had no effect (figure 6J,K). These data indicate that PKM2/HIF-1α axis participates in pluripotent polarisation of tumour macrophages.
To investigate the impact of PKM2+ macrophages on clinical features of human HCC, we collected and analysed clinical data on 91 patients with HCC (online supplemental table 1) and divided these subjects into two groups according to the median value of dual immunohistochemical staining PKM2+CD68+ cell density (figure 6L). We found a striking inverse association between PKM2+CD68+ cell density in HCC tumours and disease-free survival (p=0.019; figure 6M). The density of PKM2+CD68+ cells was also associated with tumour size (p=0.045), tumour, node, metastases stage (p=0.009) and cirrhosis (p=0.021; online supplemental table 6). In multivariate analysis, the number of PKM2+CD68+ cells was an independent prognostic factor for disease-free survival (p=0.048; online supplemental table 7).
FN1 derived from hepatoma cells orchestrates glycolytic metabolism-elicited macrophage polarisation
In some cancer patients, high levels of FN1 are associated with poor prognosis.28–30 We detected extremely higher expression of FN1 in HCC tumour tissues compared with paired non-tumoural liver tissues (figure 7A). Analogously, increased level of FN1 was detected in HCC-SN, as compared with liver-SN (figure 7B). These findings prompted us to investigate whether FN1 contributed to HCC-SN-induced metabolic reprogramming and pluripotent polarisation of macrophages. Stimulation for 24 hours with recombinant FN1 elicited upregulation of the key rate-limiting glycolytic enzyme PKM2 in monocytes (figure 7C) and increased activities of those cells to incorporate and utilise glucose (figure 7D,E). Similarly, recombinant FN1 mimicked the effect of HCC-SN to induce production of IL-1β, IL-12p70 and TNF-α and expression of HLA-DR and PD-L1 in monocytes (figure 7F,G).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Tumour-derived FN1 facilitated sequential glycolysis, inflammatory cytokine production and PD-L1 expression in macrophages. (A) Relative expression of FN1 in tumour tissue and paired non-tumoural liver tissue (n=33). (B) FN1 concentrations in liver-SN and HCC-SN (n=5). (C–G) Treatment of FN1 led to increases of PKM2 expression (C), 2-NBDG incorporation (D), lactate production (E), inflammatory cytokine production (F), as well as CD86, HLA-DR and PD-L1 expression (G), in blood CD14+ cells. (H–K) Effect of TLR4 blockade on FN1-elicited or HCC-SN-elicited lactate production (H), cytokine production (I), HLA-DR expression (J) and PD-L1 expression (K) in blood CD14+ cells (each n=4). (L–O) Knock-down of Fn1 (L) in Hepa1-6 hepatoma suppressed glycolytic enzyme expression (M), inflammatory cytokine expression (N), as well as CD86, IA-IE and PD-L1 expressions (O), in tumour macrophages (each n=5). Results are expressed as the mean±SEM. *P<0.05, **p<0.01, ***p<0.001. 2-NBDG, 2-(N-[7-nitrobenz-2-oxa-1,3-diazol-4-yl] amino)-2-deoxyglucose; FN1, fibronectin 1; HCC-SN, supernatants from cultures of primary HCC cells; PD-L1, programmed death ligand-1.
It is noteworthy that TLR4 is an important physiological receptor of FN1.31–33 We therefore used an antibody to specifically shield the TLR4 signals in the presence of either recombinant FN1 or HCC-SN. As expected, pretreatment of monocytes with this anti-TLR4 mAb effectively suppressed their glycolytic metabolism, cytokine production and HLA-DR and PD-L1 expression in these cells stimulated by exposure to HCC-SN or recombinant FN1 (figure 7H–K). Moreover, we knocked down the FN1 gene in human hepatoma cells Huh7 and in mouse Hepa1-6 hepatoma (figure 7L and online supplemental figure 6A). These treatments significantly impaired the abilities of glycolysis and cytokine production, as well as the expression of CD86, HLA-DR and PD-L1, in macrophages in vitro and in hepatoma-bearing mice (figure 7M–O and online supplemental figure 6B-D). Thus, FN1 derived from hepatoma cells may represent an important factor resulted in glycolytic metabolism-elicited macrophage polarisation.
Discussion
Our present study provides evidence that PD-L1 expressed by host macrophages represents a mechanism involved in cancer immune privilege and reflects an existence of classical innate antitumourigenic immunity. PD-L1 blockade liberates antitumourigenic activity of macrophages.
It is noteworthy that PD-L1 leads to the exhaustion of tumour-specific T cells.9 34 Thus, it is reasonable to assume that PD-L1 expression might correlate with clinical response to anti-PD-1/PD-L1 treatment. However, several clinical studies have shown that, although the underlying mechanism is still unclear, PD-L1 expressed by cancer cells and host stromal cells represents different biological and clinical significance.8 9 In fact, PD-L1 expressed by cancer cells can be regulated by many biological processes, including chromosomal rearrangements, copy number alterations, oncogenic pathway dysregulation and epigenetic modulation.35–37 Nevertheless, PD-L1 expressed by host cells is induced mainly by the local tumour environments.8 10 In this study, we show that PD-L1+ host macrophages, representing the major source of PD-L1 in HCC tumours, exhibit a glycolytic phenotype. We demonstrate that suppression of glycolytic metabolism ex vivo abrogates PD-L1 expression in tumour macrophages. Interestingly, suppression of PD-L1 by specific Ab abolishes PD-L1-dominant immune privilege and additionally enhances glycolytic macrophage-mediated specific T cell cytotoxicity. It is plausible that it is not PD-L1 per se, but rather the cells that express PD-L1, that determine the therapeutic efficiency of anti-PD-1/PD-L1 treatment.38 This notion is supported by our observation that PD-L1+ glycolytic macrophages exhibit an HLA-DRhighCD86high phenotype and produce significant antitumourigenic IL-12p70.
Despite recent success in demonstrating the importance of glycolysis in sustaining the polarisation of classical activated macrophages (M1) and OXPHOS in regulating alternative activation of macrophages (M2),13 the direct metabolic mechanisms tumour-infiltrating macrophages during cancer progression are still unclear. Our data show that, in HCC tumour tissues, glycolytic metabolism is involved in PD-L1+ macrophage polarisation, whereas OXPHOS is restricted to the energy needs in PD-L1– macrophages. More importantly, we show that glycolytic metabolism simultaneously controls antitumourigenic and protumourigenic activities of PD-L1+ macrophages. On the one hand, suppression of glycolytic metabolism reduces the PD-L1 expressed by tumour macrophages, which in turn restores the abilities of tumour T cells to produce IFN-γ and kill autologous hepatoma cells. On the other hand, inhibition of glycolytic metabolism also reduces the expression of MHC class II molecule HLA-DR and costimulatory CD86, as well as the production of antitumourigenic IL-12p70, by tumour macrophages. Therefore, although targeting glycolytic metabolism of macrophages abrogates PD-L1-mediated immune privilege, this strategy also suppresses antitumourigenic innate immunity. This hypothesis is compatible with previous studies showing that glycolytic metabolism also contributes to the effector function of cytotoxic T and NK cells.39 40
In addition to its role in glycolysis, PKM2 supports the function of transcription factors including HIF-1α and STATs.25–27 Our study shows that PKM2/HIF-1α axis has a significant role in regulating PD-L1 expression in tumour macrophages. Consistent with this, the density of PKM2+CD68+ cells in tumour tissues was correlated with advanced disease stages and poor survival in patients with HCC. It should be emphasised that, although the increase of PKM2+CD68+ cells in tumour tissue predicts a poor prognosis, this situation can be used as a predictive biomarker of benefit from anti-PD-1/PD-L1 treatment, because blocking PD-L1 signal liberates the antitumourigenic activity of PKM2+CD68+ cells. Therefore, studying the mechanisms that can selectively modulate or reverse the phenotype of glycolytic macrophages might provide a precise strategy for anticancer therapy.
In patients with cancer, FN-1 expression is usually higher in malignant tumour than in corresponding benign or normal tissue.28–30 Experimental studies have revealed that FN-1 does not only promote tumour growth and metastasis, but also modulates the functions of immune cells.31–33 In support of those findings, the results of three sets of experiments in our investigation provide evidence that FN-1 is an important factor that is produced by hepatoma cells to trigger glycolytic metabolism-mediated pluripotent polarisation of macrophages. First, recombinant FN-1 is able to mimic the ability of HCC-SN to induce the pluripotent polarisation of macrophages. Second, pretreatment with an anti-TLR4 mAb to antagonise the interactions between FN-1 and its receptors partially suppressed proinflammatory responses and subsequent PD-L1 expression in HCC-SN-exposed monocytes. Third, silencing of FN-1 in Hepa1-6 hepatoma-bearing mice attenuated the glycolysis, cytokine production and PD-L1 expression in tumour macrophages in vivo. Therefore, FN-1 generated in hepatoma environments may constitute an important mediator of glycolytic metabolism-elicited pluripotent polarisation of macrophages. Notably, besides producing FN-1, hepatoma cells also secrete molecules (eg, hyaluronan fragments), which also contributed to the expression of PD-L1 in macrophages.41 Studying the mechanisms that can selectively modulate the functional activities of macrophages might provide a novel strategy for anticancer therapy.
In the current study, FN-1 derived from cancer cells promotes glycolytic activation of macrophages by triggering TLR4, and thereby induce the macrophages to express significant amount of PD-L1, which in turn impaired the intrinsic antitumourigenic activities of glycolytic macrophages. Our results provide important insights into the significance about how glycolytic macrophages in tumours may perform a suppressive role by stimulating PD-L1 expression, which would be helpful for the rational design of novel immune-based anticancer therapies.
Data availability statement
All data relevant to the study are included in the article or uploaded as supplementary information.
Ethics statements
Patient consent for publication
Ethics approval
All human samples were anonymously coded in accordance with local ethical guidelines (as stipulated by the Declaration of Helsinki). Written informed consent was obtained from each patient, and the study protocol was approved by the Review Board of the First Affiliated Hospital of Nanjing Medical University (GZR2020/09). Participants gave informed consent to participate in the study before taking part.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Footnotes
L-GL, Z-LZ and X-YW are joint first authors.
LL, G-BZ, MZ and YC are joint senior authors.
Contributors All authors have full access to all data used in the study and take responsibility for the integrity of the data and the accuracy of the data analysis. L-GL, Z-LZ, X-YW, B-YL, J-YL, SL and G-BZ acquired experimental data. L-GL, Z-LZ and YC provided administrative, technical or material support. L-GL, G-BZ, M-XZ and YC were involved in study design and obtaining funding.
Funding This work was supported by project grants from the National Key Research and Development Program of China (2017YFA0205200), the National Natural Science Foundation of China (82071767, 81772602, 91742105, 91942309), the Jiangsu Provincial Key Research Development Program of China (BE2018750), the Key Laboratory of Emergency and Trauma, Ministry of Education (KLET-201913), the Research Start Project of Zhuhai People's Hospital (No.2020ycqd001), and the Science and Technology Development Fund, Macau SAR (0011/2019/AKP).
Competing interests None declared.
Provenance and peer review Not commissioned; externally peer reviewed.
Supplemental material This content has been supplied by the author(s). It has not been vetted by BMJ Publishing Group Limited (BMJ) and may not have been peer-reviewed. Any opinions or recommendations discussed are solely those of the author(s) and are not endorsed by BMJ. BMJ disclaims all liability and responsibility arising from any reliance placed on the content. Where the content includes any translated material, BMJ does not warrant the accuracy and reliability of the translations (including but not limited to local regulations, clinical guidelines, terminology, drug names and drug dosages), and is not responsible for any error and/or omissions arising from translation and adaptation or otherwise.