Article Text

Abstract
Objective Chronic obstructive pulmonary disease (COPD) is a global disease characterised by chronic obstruction of lung airflow interfering with normal breathing. Although the microbiota of respiratory tract is established to be associated with COPD, the causality of gut microbiota in COPD development is not yet established. We aimed to address the connection between gut microbiota composition and lung COPD development, and characterise bacteria and their derived active components for COPD amelioration.
Design A murine cigarette smoking (CS)-based model of COPD and strategies evaluating causal effects of microbiota were performed. Gut microbiota structure was analysed, followed by isolation of target bacterium. Single cell RNA sequencing, together with sera metabolomics analyses were performed to identify host responsive molecules. Bacteria derived active component was isolated, followed by functional assays.
Results Gut microbiota composition significantly affects CS-induced COPD development, and faecal microbiota transplantation restores COPD pathogenesis. A commensal bacterium Parabacteroides goldsteinii was isolated and shown to ameliorate COPD. Reduction of intestinal inflammation and enhancement of cellular mitochondrial and ribosomal activities in colon, systematic restoration of aberrant host amino acids metabolism in sera, and inhibition of lung inflammations act as the important COPD ameliorative mechanisms. Besides, the lipopolysaccharide derived from P. goldsteinii is anti-inflammatory, and significantly ameliorates COPD by acting as an antagonist of toll-like receptor 4 signalling pathway.
Conclusion The gut microbiota–lung COPD axis was connected. A potentially benefial bacterial strain and its functional component may be developed and used as alternative agents for COPD prevention or treatment.
- intestinal microbiology
- immunology
Data availability statement
Data are available on reasonable request. All data relevant to the study are included in the article or uploaded as online supplemental information. The data that support the findings of this study are available from the corresponding author, on reasonable request.
Statistics from Altmetric.com
Significance of this study
What is already known on this subject?
Although there were many previous studies, ways of management of chronic obstructive pulmonary disease (COPD) were still encountered with difficulty.
The microbiota composition in the respiratory tract was shown to closely related to COPD severity and development. However, whether the gut microbiota–lung COPD axis also existed was not established.
Furthermore, there were no microbiota derived agent(s) identified to effectively ameliorate COPD.
What are the new findings?
The gut microbiota–lung COPD axis was established in this study.
A bacterial strain that showed COPD amelioration effect was isolated.
An anti-inflammatory lipopolysaccharide (LPS) molecule purified from this bacterium can ameliorate COPD.
How might it impact on clinical practice in the foreseeable future?
The beneficial bacterial strain (Parabacteroides goldsteinii MTS01), together with its derived LPS (P. goldsteinii-LPS) may be developed as an agent for clinical practice in COPD.
Introduction
Chronic obstructive pulmonary disease (COPD) is a multidimensional progressive lung inflammatory disease, with pathological changes in the large and small airways.1 2 COPD has a huge global prevalence (251 million cases in 2016) and causes high mortality (3.17 million deaths in 2015), with a prediction of being the third leading cause of mortality in 2020.3–5 There are many measures for COPD management; however, persistent and progressive pulmonary inflammation is still common among patients.6 7
The roles of microbiota in development of host health are emphasised,8–10 some supporting the role of respiratory tract microbes towards the onset, progression, promotion and severity of COPD chronicity.11 Emerging evidences indicate close interactions between gut microbiota composition and lung immunity.12 However, the relationship between gut microbiota composition and COPD development is still to be well characterised.
We established the gut microbiota–lung COPD axis using a mouse cigarette smoking (CS) model, and strategy of faecal microbiota transplantation (FMT) and antibiotics (ABX) treatment. An intestinal commensal Parabacteroides goldsteinii (Pg) was highlighted to be significantly negatively associated with severity of COPD. Oral administration of isolated Pg MTS01 on its own significantly ameliorated CS-induced COPD syndromes, characterised by reducing inflammation and improving cellular ribosomal biogenesis activity and mitochondrial functions in intestine, systematically restoring aberrant amino acids metabolism in sera, and reducing lung tissue inflammations. The lipopolysaccharide (LPS) derived from Pg-LPS was subsequently characterised to act as an toll-like receptor 4 (TLR-4) receptor antagonist and worked as an active component that ameliorated CS-induced COPD. Our study highlighted the important causality role of gut microbiota in COPD pathogenesis and established the gut-lung COPD axis. Furthermore, a potential probiotic bacterium whose LPS might work as a COPD ameliorating agent was reported.
Methods
Model of CS-induced COPD
C57BL/6 mice (8–10 weeks old) were purchased from the National Laboratory Animal Center (Taipei, Taiwan) and kept under sterile conditions (following a 12 hours light/dark cycle), with 1-week acclimatisation period. Mice were fed with normal chow diet (Laboratory Autoclavable Rodent Diet 5010; LabDiet, USA) during the entire experiment.
The CS-induced COPD model used was modified from He et al.13 Briefly, mice were exposed to CS from 12 3R4F cigarettes (Kentucky University) twice a day (twenty-four cigarettes per day), 5 day-week for 12 weeks, with body weights being monitored weekly. Pulmonary function was assessed by forced pulmonary manoeuvre system (Buxco Research Systems, Wilmington, USA), and bronchoalveolar lavage fluid (BALF) and paraffin sections of the lungs were obtained in every mouse to confirm the COPD phenotypes. Detailed methods of animal experiments and measurements can be found in online supplemental methods.
Supplemental material
Analysis of gut microbiota structure
We followed the protocol per our previous studies14–16 and detailed methods and workflow can be found in online supplemental methods and online supplemental figure S1. In brief, partial least squares discriminant analysis (PLS-DA) analysis was used to evaluate sample-species complexity (Beta diversity analysis). The statistically significant biomarkers (LDA score (log10) >4.0 with p<0.05) in CS mice were identified using the LEfSe analysis and also confirmed by using metagenomeSeq (q<0.05). Furthermore, bacterial species significantly altered by ABX treatment (q<0.05) or FMT (p<0.05) were identified by using metagenomeSeq.
Pg cultivation and genome sequencing
Pg MTS01 was isolated from faeces of mice in ABX experiments and grown at 37°C in anaerobic condition. Illumina and Pacbio platforms were used for Pg MTS01’s full-genome sequencing. Detailed methods of experiments can be found in online supplemental methods.
ScRNAseq analysis of colon and lung tissues and sera metabolomics analysis
The colon and lung tissues of CTL, CS and CS administered with Pg mice groups were collected after 12 weeks and subjected for 10 x Genomics Single Cell 3’ Gene Expression analysis. The sera metabolites were analysed by GC-MS and LC-MS. Detailed methods of experiments and analysis can be found in online supplemental methods.
Pg-LPS purification and its antagonistic effects
Pg-LPS were isolated by using the hot phenol-water extraction. The antagonistic effect was analysed on HEK-Blue-mTLR4 cells, mouse CD11c+ dendritic cells (DC) cells, human PBMC cells and mouse B cells treated with Escherichia coli O111:B4 LPS. Detailed methods of experiments can be found in online supplemental methods.
Statistical analysis
Data are shown as means±SD and medians±IQR for parametric and non-parametric analysis, respectively. Differences between two groups are assessed using unpaired two-tailed Student’s t-test. Data sets involving more than two groups are assessed by one-way analysis of variance, followed by non-parametric Kruskal-Wallis test with Newman-Keuls multiple comparison (see figure legends).
Results
Evidence of gut microbiota involvement in COPD development
To see whether gut microbiota may contribute to CS induced COPD development, we used a murine CS model of chronic pulmonary inflammation (online supplemental figure S2). CS mice smoking for 12 weeks significantly lost body weight (online supplemental figure S2a, b), and increased infiltration of immune cells in BALF (online supplemental figure S2c). By contrast, weight gain and decreased infiltrated immune cells were observed in CS mice cohoused with control (CO-CTL) group (online supplemental figure S2a–c). Comparatively, control group cohoused with CS (CO-CS) group developed aberrant phenotypes (online supplemental figure S2a–c).
Concordantly, histological analysis of lung tissue sections revealed CS-exposed mice show peripheral emphysematous changes characterised by alveolar wall destruction and enlargement of airspace compared with CTL mice. By contrast, CO-CTL group restored the pathology, while CO-CS group exacerbated (online supplemental figure S2d, e). Concordantly, similar phenomena on production of interleukin (IL)-1β and tumour necrosis factor (TNF)-α in CD11C+ DCs, and F4/80+CD11b+macrophage cells (online supplemental figure S2f, g), IL-17A production in CD4+NKP46+Th17 cells, and IL-10 production in CD4+FoxP3+Treg cells were also observed (online supplemental figure S2h). Furthermore, assessment of pulmonary function also showed consistent results (online supplemental figure S2i–l). Although the gut microbiota was not determined, cohousing results speculated that beneficial and adverse gut bacteria may respectively exist in CTL and CS mice.
ABX treatment reduces CS-induced COPD pathogenesis
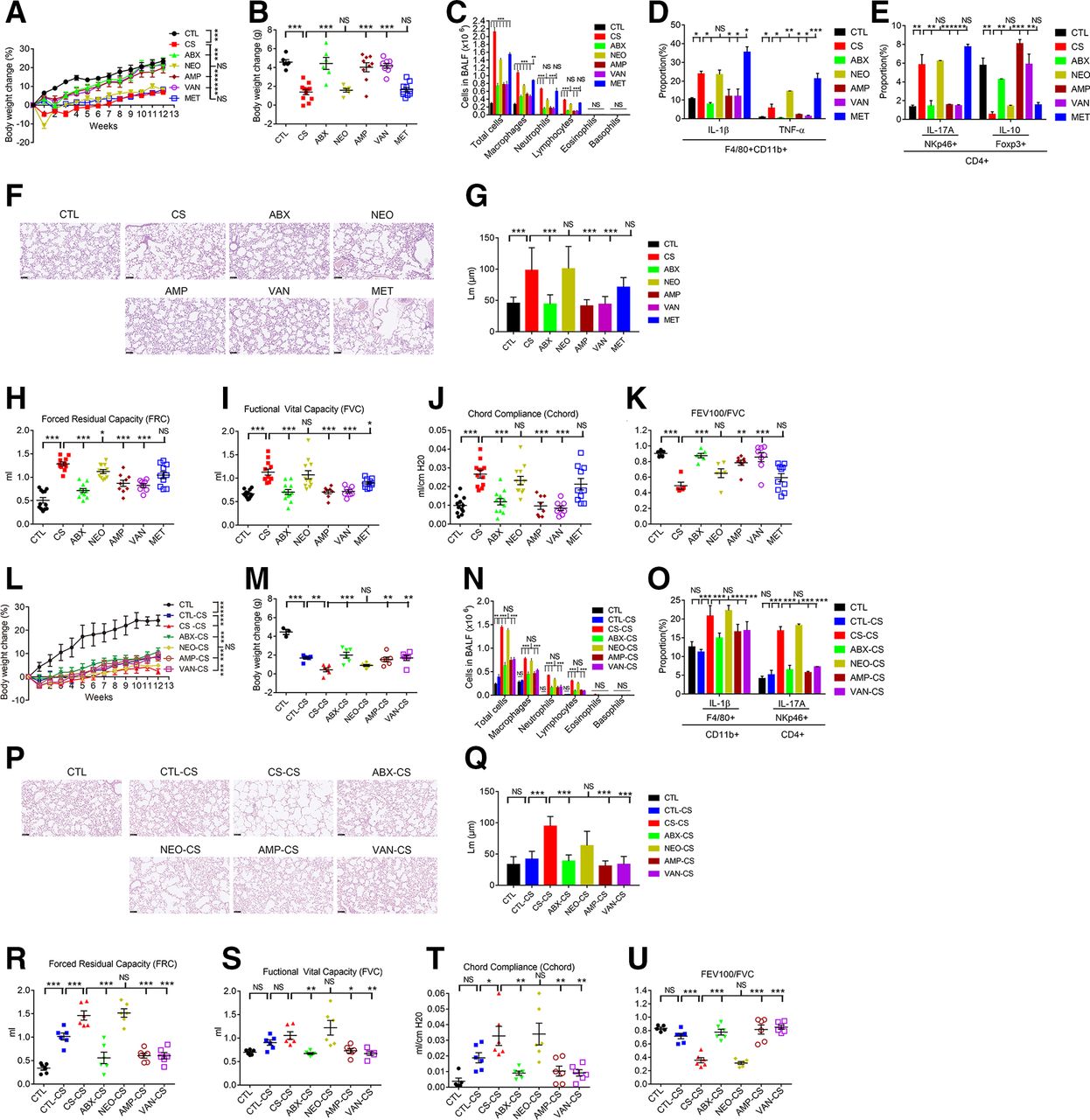
To determine the importance of gut microbiota composition in COPD development, individual (vancomycin (VAN), neomycin (NEO), metronidazole (MET) and ampicillin (AMP)) and combinational ABX17 were orally administrated to mice 2 weeks before CS. Aberrant phenotypes observed in CS mice (figure 1A–K) were significantly ameliorated in the ABX, AMP and VAN-treated groups. In contrast, oral NEO or MET treatment did not significantly affect COPD pathogenesis (figure 1A–K). Thus, commensal microbiota composition affect the inflammations and pathogenesis of CS-induced COPD and treatment with optimal ABX predisposes the mice to reduced pathogenesis in the lung of COPD.

Establishment of the gut microbiota-COPD development relationship. Mice were orally administered with antibiotics (NEO, AMP, VAN, MET and ABX) to evaluate the effect of antibiotics treatment on COPD development in CS mice. Percentage of body weight change was measured throughout the 12-week period (A). Body weight change (B), BALF cellular composition (C), flow cytometry analysis on production of IL-1β and TNF-α in F4/80+CD11b+ macrophages (D), IL-17A in CD4 +NKp46+Th17 cells and IL-10 in CD4 +FoxP3+Treg cells (E), histopathology in lung (F) and the mean linear intercept of lung tissue (G) and lung functions analyses including FRC (H), FVC (I), Cchord (J) and FEV100/FVC (K) were measured after 12 weeks. Through use of the faeces microbiota of antibiotics-treated mice as the donor, FMT was performed to see their effects on COPD development in CS recipient mice. Percentage of body weight change was measured throughout the 12-week period (L). Body weight change (M), BALF cellular composition (M), flow cytometry analysis on production of IL-1β in F4/80+CD11b+macrophages and IL-17A in CD4 +NKp46+Th17 cells (O), lung histopathology (P) and the mean linear intercept of lung tissue (Q), and lung functions analyses including FRC (R), FVC (S), Cchord (T), and FEV100/FVC (U) were measured after 12 weeks. For FMT, faeces microbiota of control (CTL), CS, ABX, NEO, AMP and VAN were collected, followed by oral administration to recipient CS mice. Except panels F and P, the data representing means±SD or medians±IQR were analysed using the Newman-Keuls multiple comparison post hoc one-way ANOVA analysis (n=6–12 for panel a-j and n=5–6 for panel k-s). *p<0.05; **p<0.01; ***p<0.001. ANOVA, analysis of variance; ABX, antibiotics; AMP, ampicillin; BALF, bronchoalveolar lavage fluid; CS, cigarette smoking; COPD, chronic obstructive pulmonary disease; FMT, faecal microbiota transplantation; IL-1β, interleukin 1β; MET, metronidazole; NS, not significant; TNF-α, tumour necrosis factor α; VAN, vancomycin.
Gut microbiota composition plays a causal effect on COPD development
The beneficial effects of ABX treatment may be mediated through the changed gut microbiota composition. To see whether the changed gut microbiota structure may play a causal effect on COPD development, FMT experiment was performed. Donor faecal microbiota from CTL or CS mice treated by either ABX, NEO, AMP or VAN were transferred to CS-recipient mice. Faecal microbiota from CTL, ABX-treated, AMP-treated or VAN-treated mice significantly ameliorated COPD syndromes (figure 1L–U). In contrast, microbiota derived from CS or NEO-treated mice failed to reverse COPD traits of CS recipients (figure 1L–U). Briefly, the COPD ameliorative effects produced by treatment of CS mice with ABX, AMP or VAN are transferrable through gut microbiota transplantation. Therefore, the changed gut microbiota structure may play a causal effect on COPD pathogenesis.
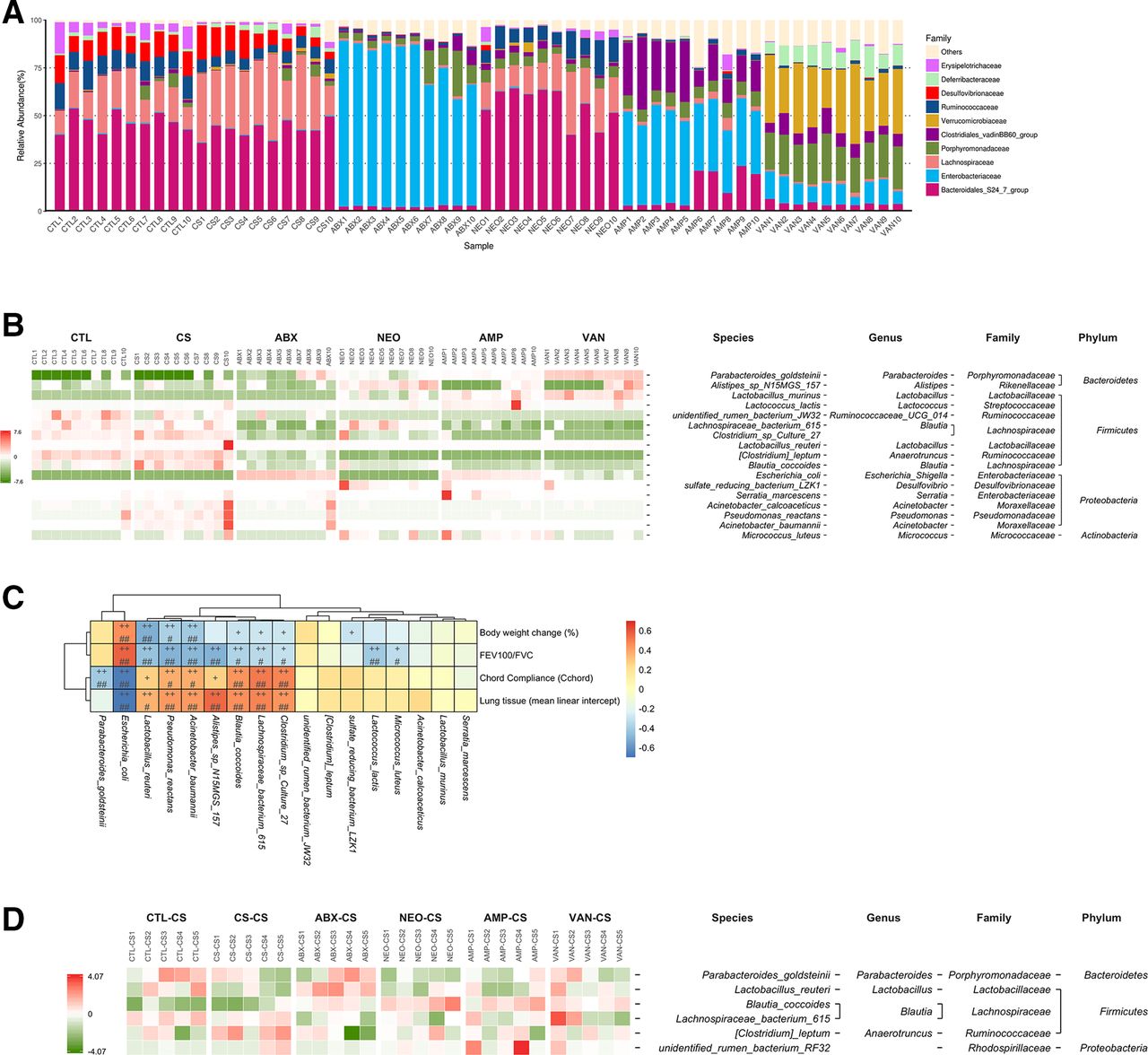
The gut microbiota compositions in each mice group were determined using 16S microbiota ribosomal DNA sequencing (figure 2A). Relative abundance of Erysipelotrichaceae, Bacteroidales and Ruminococcaceae were significantly reduced, while those of Lachnospiriaceae were increased in the faeces of CS mice in contrast to the CTL mice (online supplemental figure S3a–c). ABX treatment and FMT both affects the composition of gut microbiota in CS mice (figure 2a and online supplemental figure S3d, e). Seventeen bacterial species significantly altered by antibiotics (ABX, NEO, AMP and VAN) in comparison with the CS group were identified by using metagenomeSeq (figure 2B) and the correlation between the 17 identified bacterial species and COPD traits was further analysed (figure 2C). P. goldsteinii and E. coli were significantly negatively correlated with the COPD severity (figure 2C), and the relative abundance of Pg and E. coli in 16S sequencing data was mostly consistent with those validated by qPCR, except that the relative abundance of Pg in the AMP group was not significantly altered in qPCR data (online supplemental figure S3f–i). Furthermore, bacterial species significantly altered by FMT between any two groups indicated only Pg was increased by trend in ABX, VAN and AMP treated mice groups (figure 2D and online supplemental figure S3j). Additionally, a positive correlation between COPD severity and Lachnospiriaceae which was significantly increased in CS mice was observed (figure 2C and online supplemental figure S3k). Therefore, together with all the data obtained, Pg was selected for subsequent studies.
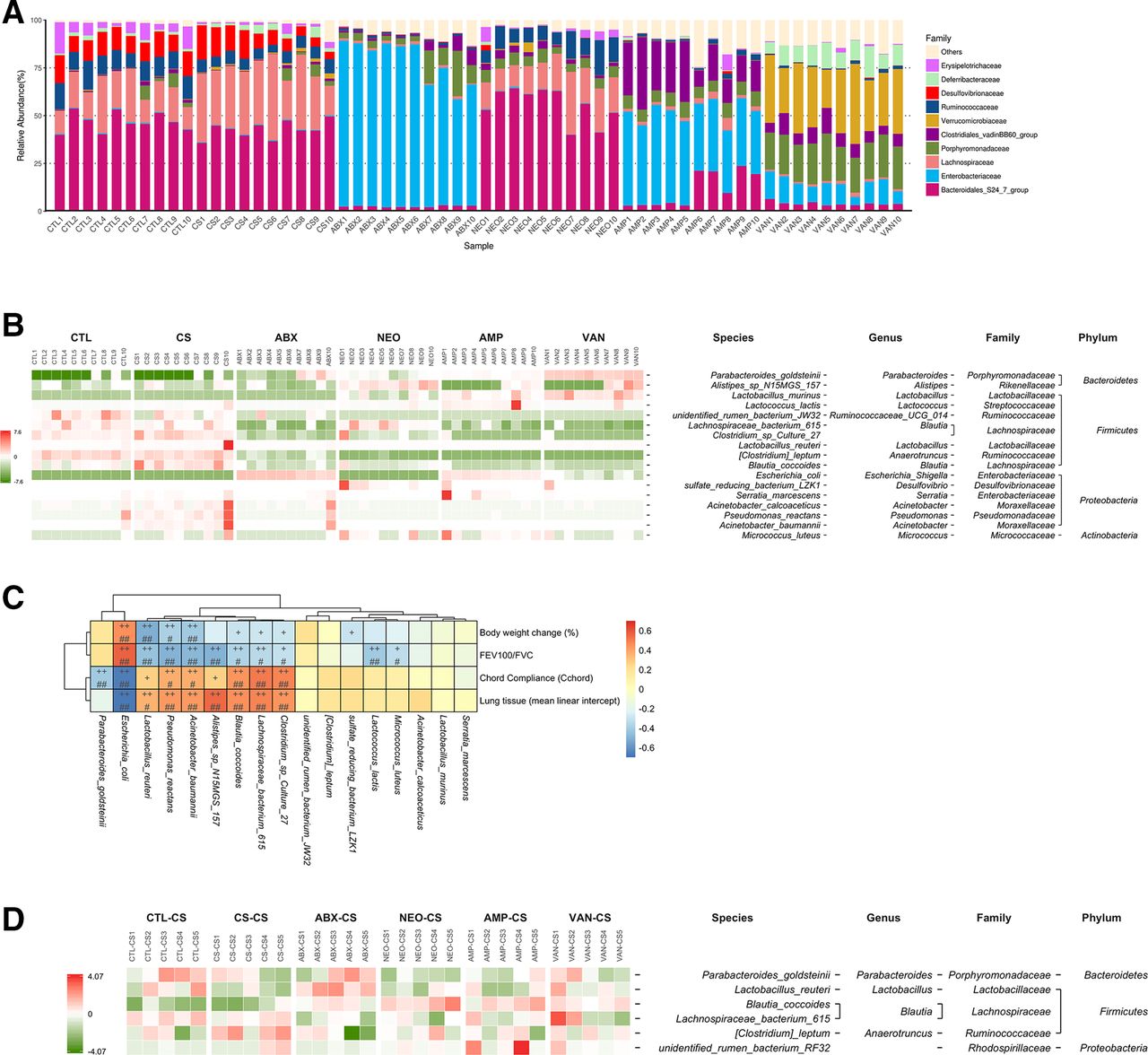
Negative association of Parabacteroides goldsteinii with COPD severity in different mice groups. The top 10 most abundant faecal microbiota bacterial families of CTL, CS, ABX, NEO, AMP and VAN treated mice groups were shown in (A). Bacterial species significantly altered by antibiotics (ABX, NEO, AMP and VAN) in comparison with the CS group (q<0.05) were shown in the heatmap (B). Spearman’s correlation analysis between the 17 identified bacterial species and COPD traits (C). The p values were corrected for multiple comparisons by using Benjamini and Hochberg (BH) method. (+p<0.05; ++p<0.01; #adjusted p<0.05; ##adjusted p<0.01). bacterial species significantly altered by FMT (CTL-CS, CS-CS, ABX-CS, NEO-CS, AMP-CS and VAN-CS) between any two groups (p<0.05) were shown in the heatmap (D). The values of the heatmap (B, D) are based on the row scaling (z-scores) of the relative abundance. ABX, antibiotics; AMP, ampicillin; CS, cigarette smoking; COPD, chronic obstructive pulmonary disease; CTL, control; FEV, forced expiratory volume; FMT, faecal microbiota transplantation; FVC, forced vital capacity; NEO, neomycin; VAN, vancomycin.
Pg MTS01 ameliorates COPD
A Pg bacterial strain MTS01 (online supplemental figure S4a) was subsequently isolated from ABX-treated faecal microbiota samples. Full-length sequencing of 16S rRNA gene of Pg MTS01 showed 99.9% identity to that of Pg JCM13446.14 Oral treatment of Pg MTS01 (2×108 colony-forming units per day per mouse) significantly increased body weight gain (by 7.4%) and ameliorated COPD phenotypes (figure 3A–J). In parallel, the expression of lung ZO-1 and Occludin mRNA-linked with the respiratory tract integrity was also increased (online supplemental figure S4b, c).

Parabacteroides goldsteinii MTS01 ameliorates COPD. Live P. goldsteinii (Pg) MTS01 at the colony-forming units of 2×108 were orally administered to CS mice to evaluate their effects on COPD development. Percentage of body weight change was measured throughout the 12-week period (A). Body weight change (B), BALF cellular composition (C), RNA expression of IL-1β and TNF-α in lung (D), histopathology in lung (E) and the mean linear intercept of lung tissue (F), and lung functions analyses including FRC (G), FVC (H), Cchord (I) and FEV100/FVC (J) were measured after 12 weeks. Except panel E, the data representing means±SD or medians±IQR (n=5–17) were analysed using the Newman-Keuls multiple comparison post hoc one-way ANOVA analysis. *p<0.05; **p<0.01, ***p<0.001. ANOVA, analysis of variance; BALF, bronchoalveolar lavage fluid; COPD, chronic obstructive pulmonary disease; CS, cigarette smoking; CTL, control; IL-1β, interleukin 1β; NS, not significant; TNF-α, tumour necrosis factor α.
CS has been shown as a risk factor of intestinal mucosal damages.18 In colon, Pg MTS01 treatment reduced expression of proinflammatory cytokines such as IL-1β and TNF-α (online supplemental figure S4d), and increased expression of ZO-1 and Occludin (online supplemental figure S4e, f) in CS mice. Pg MTS01 further restored TNF-α-induced increase of cell layer TEER permeability in a Caco-2-cells in vitro model (online supplemental figure S4g). Effects of Pg MTS01 on body weight change, histopathology in lung and liver, serum GPT and BUN levels revealed that Pg MTS01 has no adverse effect on healthy mice (online supplemental figure S4h–l).
Pg MTS01 reduces intestinal inflammation and enhances cellular ribosomal and mitochondrial activity in CS mice
Pg MTS01 administration reduced pro-inflammatory cytokines production and increased intestinal integrity in colon (online supplemental figure S4d–g). To further characterise the effects of Pg MTS01 administration on physiology of colon cells, single-cell RNA sequencing (scRNAseq) and analyses of colon tissue-derived cells of CS mice with/without Pg MTS01 treatment were conducted (figure 4A,B). Specifically, in CS compared with CTL mice, an increased expression of genes involved in fatty acid/eicosanoids metabolism, membrane microvillus formation and endopeptidase activity, while reduced expression of genes involving ribosomal biogenesis and mitochondrial activity were observed (online supplemental dataset 1 and figure S5a, b). By contrast, Pg MTS01 administration significantly upregulated genes of cellular antioxidants, redox processes, antibacterials, mitochondrial depolarisation, ribosomal activity and mucin production in CS mice (figure 4C,D and online supplemental dataset 2). Expression levels of Mptx1 (mucosal pentraxin 1), Ang4 (angiogenin, ribonuclease A family, member 4) and Muc2 (mucin 2) (online supplemental figure S5c–e), RNR1 (mitochondria encoded 12S rRNA), RNR2 (mitochondria encoded 16S rRNA), Cytb (mitochondria encoded cytochrome B), PGC-1α (peroxisome proliferator-activated receptor γ coactivator 1α), ND-5 (mitochondria encoded NADH-ubiquinone oxidoreductase chain 5 protein) and mitochondria encoded Transcription Factor A genes related to oxidative stresses and ribosomal/mitochondrial activities, were further validated by qPCR (online supplementary table S1 and figure S5f–k). By contrast, Pg MTS01 did not significantly downregulate any GO pathway in CS mice.
Supplemental material

Single cell RNA sequencing (scRNAseq) analyses of colon intestinal epithelial cells (IEC) and lung tissue of CTL, CS and CS administered with Parabacteroides goldsteinii MTS01 mice groups. For colon IEC, the results of t-SNE analysis (A) and proportion of cell type (B) determined by SingleR, heatmap of differentially expressed genes between CS and CS+Pg group (C), as well as GO enrichment pathways analysis (D) were shown. for lung tissues, the results of t-SNE analysis (E) and proportion of cell type (F) determined by SingleR, heatmap of differentially expressed genes between CS and CS+Pg group (G), as well as GO enrichment analysis (H, I) were shown. DC, dendritic cells; CS, cigarette smoking; CTL, control
In COPD, reduced ribosomal biogenesis activity is linked to mitochondria dysfunction.19 The oxygen consumption rate (OCR) and extracellular acidification rate (ECAR) linked to oxidative phosphorylation and non-mitochondria glycolysis function, respectively, were measured. Pg MTS01 increased the aberrant OCR activity, and reduced ECAR in CS mice (online supplemental figure S5l, m). Pg MTS01 may therefore reduce inflammations and restore the abrogated cellular ribosomal and mitochondrial activity in colon of CS mice.
Pg MTS01 systematically restores aberrant amino acid-metabolism in Cs mice
The phenomenon that oral administration of Pg MTS01 ameliorated COPD lung pathogenesis might be related to restoration of systematic aberrant metabolism in blood. To further characterise whether any metabolomics in mice sera were affected by Pg MTS01 treatment, non-targeted positively and negatively charged metabolite-analyses were analysed. Ingenuity canonical pathways analyses on sera metabolites indicated that Pg MTS01 administration most significantly improved aberrant amino acid-related metabolism and tRNA-amino acid charging activities in CS mice, including increased biosynthesis of citrulline and proline, degradation of phenylalanine and glycine, and turnover of arginine (online supplemental figure S6a–d). Pg MTS01 may therefore enhance amino acids metabolism related to cellular ribosomal and mitochondrial activities and urea cycle turnover (online supplemental figure S6c).
Pg MTS01 reduces lung inflammations in Cs mice
Administration of Pg MTS01 enhanced lung integrity (online supplemental figure S4b, c) and reduced inflammations in COPD lung (figure 3D). Further lung tissue single-cell RNA sequencing analyses indicated that CS also increased viable counts of B cells, macrophages as well as reduced T cells and neutrophils counts, which were reversed by Pg MTS01 administration (figure 4E,F). Furthermore, Pg MTS01 significantly restored the aberrant gene expression patterns identified in CS mice. Especially, genes involved in antigen processing and presentation via MHCII, and B cell receptor signalling pathway in B cells were reduced (figure 4G–I, online supplemental dataset 3, 4 and figure S7a, b). By contrast, genes involving antioxidant activity, eicosanoid-binding activities, fatty acids synthesis and immune responses were upregulated (figure 4G–I, online supplemental dataset 3-4 and figure S7a, b). Among these, expression of Scgb1a1 that played important functions in anti-inflammatory properties was most significantly upregulated by Pg MTS0120 21 (online supplemental figure S7c). However, Pg MTS01 did not significantly reduce sera IgG level in CS mice (online supplemental figure S7d). Briefly, Pg MTS01 reduced overinflammatory immune responses, including those related to B cells receptor signalling in lung tissues.
Pg MTS01 LPS (Pg-LPS) antagonises E. coli LPS-induced inflammations
The proinflammatory LPS has been associated with increased oxidative stress, production of proinflammatory cytokines and COPD pathogenesis.22 As shown in figure 5A,B, LPS activities measured in the sera and lung BALF were all significantly increased in CS mice compared with CTL mice. By contrast, Pg MTS01 administration significantly reduced these LPS activities detected.

LPS purified from Parabacteroides goldsteinii (Pg) MTS01 (Pg-LPS) antagonises the inflammation elicited by Escherichia coli O111.B4 LPS (EC-LPS). The proinflammatory LPS activities in BALF (A) and sera (B) among CTL, CS and CS+Pg groups detected by HEK-Blue-mTLR4 reporter cells were shown. The circular genome map of Pg MTS01 was shown in (C). The biosynthesis pathway of kdo2-lipidA in EC was shown in (D), indicating LpxL and LpxM are responsible for adding the fifth and sixth acyl chains to EC lipid A. The putative genes for Kdo2-lipid a synthesis of Pg MTS01 were identified by blast searches using the EC MG1655 lipid A biosynthesis genes as queries. The gene ID and locations as well as identities in comparison with those in EC MG1655 and Bacteroides dorei DSM17855 were indicated (E). Effects of different doses of LPS derived from EC O111.B4 (EC-LPS) or Pg MTS01 (Pg-LPS) on NF-kB activations in HEK-Blue-mTLR4 reporter cells were shown in (F). The antagonistic results of Pg-LPS on EC-LPS induced NF-kB activations in HEK-Blue-mTLR4 cells were shown in (G). The competition of Pg-LPS on FITC-EC-LPS binding on HEK-Blue-mTLR4 cells were shown in (H). Effects of EC-LPS or Pg-LPS in single or in combination administration on expression of CD86 and proliferation of B cells were analysed by flow cytometry and shown in (I) and (J), respectively. The antagonistic results of Pg-LPS on EC-LPS induced TNF-α in CD11c+ bone marrow derived dendritic cells and IL-1β in human peripheral mononuclear cells were shown in (K) and (L), respectively. Cytokine levels were expressed as relative percentage in comparison with the level induced by EC-LPS (grey BAR). The data represent means±SD analysed using the Newman-Keuls multiple comparison post hoc one-way ANOVA analysis. *p<0.05; **p<0.01; ***p<0.001; ****p<0.0001. B cell proliferation indicated by halving of CFSE, a green fluorescence dye, which is partitioned approximately equally between the progeny on division. ANOVA, analysis of variance; BALF, bronchoalveolar lavage fluid; CS, cigarette smoking; CTL, control; IL-1β, interleukin 1β; LPS, lipopolysaccharide; NS, not significant.
A BLAST analysis of whole genome sequence of Pg MTS01 (figure 5C) highlighted genes involved in lipid A synthesis resembled those identified in Bacteroides dorei (figure 5D,E) and Bacteroides thetaiotaomicron.23 Bacteroides species-derived LPS subtypes exhibited significantly lower endotoxicity and immune-stimulatory responses relative to enteric bacteria-isolated proinflammatory LPS.24 25 E. coli normally produced a lipid A molecule with six acyl chains.26 By contrast, Pg MTS01 lacked LpxM, one ortholog of the acyltransferases genes, and was expected to produce lipid A with five acyl chains (figure 5D,E).
To see whether Pg MTS01 and E. coli exhibited subtype-specific LPS immunogenicity differences, their effects on cellular activation were measured. The NF-kB activations in HEK-Blue-mTLR4 reporter cells that specifically measured the proinflammatory LPS activity were used. While LPS derived from E. coli O111.B4 (EC-LPS) started to activate NF-kB activity at the concentration of 1 ng/mL, Pg-LPS at the concentration of 1000 ng/mL still failed in activation (figure 5F). Pg-LPS further showed antagonistic effects on EC-LPS activity. Under the concentration ratio of 1:1, the NF-kB activity activated by EC-LPS was reduced up to 80% by addition of Pg-LPS (figure 5G). Further studies indicated that Pg-LPS competed with EC-LPS for TLR4 receptor binding in a dose-dependent manner (figure 5H).
Whether the phenomenon of EC-LPS and Pg-LPS interaction also occurs in other immune cells was next addressed. Expression of CD86 and proliferation of CD19+ B cells by EC-LPS were antagonised by Pg-LPS (figure 5I,J). Concordantly, overproduction of TNF-α in CD11c+ bone marrow derived DCs (figure 5K), and IL-1β in human peripheral mononuclear cells (figure 5L) by EC-LPS treatment was also dose-dependently reduced by Pg-LPS administration, respectively. No cytotoxicity revealed in cell treated with Pg-LPS (online supplemental figure S8a). Briefly, Pg-LPS competes with EC-LPS and antagonises its proinflammatory activity.
Pg-LPS ameliorates CS-induced COPD syndromes
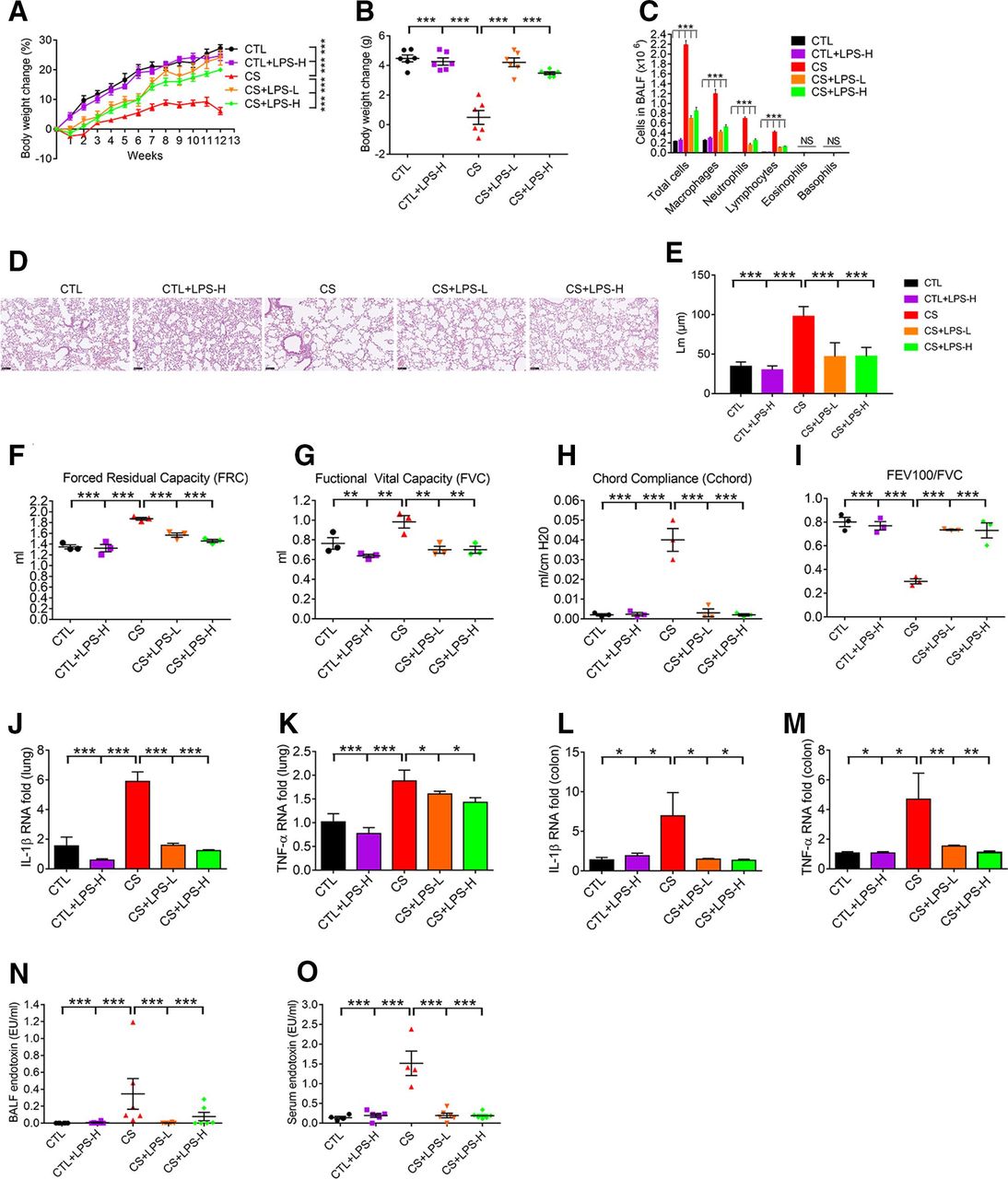
Pg-LPS was subsequently investigated to see its potentiality in ameliorating CS-induced COPD pathogenesis. As intraperitoneal (i.p.) administration of proinflammatory LPS enhanced emphysema development in COPD,27 we tested the effect of i.p. treatment of Pg-LPS in CS mice. CS mice with or without i.p. treatment of Pg-LPS (0.2 or 2 µg in 100 µL saline for each mice) twice per week were phenotyped for COPD. Contrary to the CS mice, Pg-LPS treatment significantly increased body weight gain by 18% and 13.9% in low and high doses, respectively (figure 6A,B), reduced BALF cellular-infiltration (figure 6C), and normalised lung pathology and functions (figure 6D–I). Furthermore, Pg-LPS reduced over-expression of proinflammatory cytokines such as IL-1β and TNF-α in lung tissues (figure 6J,K) and colon (figure 6L,M) in COPD mice. Concordantly, increased LPS concentration and activity in the BALF (figure 6N) and sera (figure 6O) of COPD mice were all significantly reduced by Pg-LPS. Further study indicated that control mice treated with Pg-LPS had normal liver and kidney functions (online supplemental figure S8b, c). Therefore, LPS of Pg MTS01 origin ameliorated CS-induced COPD in mice.

Pg-LPS ameliorates COPD development in CS mice. After i.p. treatment of Pg-LPS, the body weight change (A, B), BALF cellular composition (C), lung histopathology (D), the mean linear intercept of lung tissue (E), lung functions analyses including FRC (F), FVC (G), Cchord (H) and FEV100/FVC (I), mRNA expression level analysis of IL-1β and TNF-α in lung tissue (J, K) and colon (L, M), respectively, and endotoxin activity in BALF (N), and sera (O) were shown. LPS-L, and H: Pg-LPS administered at 0.2 and 2 µg/twice per week, for 3 months. Except panel D, statistical analysis was performed using the Newman-Keuls multiple comparison post hoc one-way ANOVA analysis (n=3–6). *p<0.05; **p<0.01; ***p<0.001. ANOVA, analysis of variance; BALF, bronchoalveolar lavage fluid; CS, cigarette smoking; COPD, chronic obstructive pulmonary disease; CTL, control; FEV, forced expiratory volume; FVC, forced vital capacity; IL-1β, interleukin 1β; i.p., intraperitoneal; LPS, lipopolysaccharide; TNF-α, tumour necrosis factor α.
Discussion
This study highlighted that gut microbiota dysbiosis plays a causal effect influencing the severity of CS-induced COPD pathogenesis. Therefore, the potential axis between gut microbiota composition and lung COPD can be established. A recent human study also found bacterial species differ between the faecal microbiome of COPD patients and healthy controls.28 Current evidences strongly indicated that not only respiratory tract, but also gut microbiota structure may both be involved in controlling COPD development. The composition of microbiota in BALF of CTL, CS and ABX mice groups was determined using 16S microbiota ribosomal DNA sequencing (online supplemental figure S9), indicating that CS or ABX treatment affect the composition of BALF microbiota. The ameliorative effect of ABX treatment in CS mice may also be partly due to changes of the lung microbiota composition. Based on these results, to maintain microbiota homeostasis in both colon and the respiratory tract has to be taken into consideration in designing an effective strategy for prevention/treatment of COPD.
During the process of 12 weeks’ smoke exposure and co-housing experiment, control group cohoused with CS (CO-CS) group developed aberrant phenotypes, as shown in online supplemental figure S2. Even so, we could not come to solid conclusion that CS-induced abnormal gut microbiota composition plays a causing effect on aberrant COPD-related phenotypes in control mice. Part of the reasons is that not only uptake each other’s faeces bacteria, cohoused mice may also lick each other’s skin fur and expose to cigarette-derived substances that could be adhered to the bedding and walls of the cage due to smoke exposure. Therefore, the aberrant COPD-related phenotypes observed in the control mice might only partially be due to transfer of abnormal gut microbiota bacteria from the smoke-exposed mice. Besides faeces microbiota, other factors may also participate in affecting COPD-related phenotypes. We, therefore, did not determine gut microbiota composition in this scenario. To confirm the possibility of gut microbiota-COPD axis, FMT was subsequently performed to establish the relationship.
Among the gut microbiota bacteria affected, increased abundance in Lachnospiraceae (online supplemental figure S3a–c, k) might be one of the contributing factors leading to aberrant inflammations and reduction of cellular physiological activities in colon. Consistently, increased abundance of Lachnospiriaceae was also reported in many other chronic inflammations related diseases such as inflammatory bowel disease, irritable bowel syndrome with diarrhoea, obesity and after stress.29 A recent human study also reported multiple members of the family Lachnospiraceae correlate with reduced lung function.28 On the other hand, FMT from ABX-treated mice reduced intestinal inflammation, promoted intestinal integrity, and ameliorated lung COPD pathogenesis, which was positively correlated with Pg abundance (figure 2B–D and online supplemental figure S3f, h, j). An assumption was developed that optimal depletion of potentially harmful bacteria and/or enrichment of beneficial bacteria in gut may be considered as an alternative strategy for amelioration of COPD. Subsequent studies using the isolated Pg MTS01 as the sole bacterium for oral administration validated its effect on COPD amelioration, thus attracting the investigation of the underlying mechanism by which orally-administered Pg MTS01 ameliorated COPD.
A recent study found that Pg was significantly decreased in patients of chronic kidney disease which was also a chronic inflammatory disease.30 There seemed to be not many reports related to Pg abundance. One of the reasons may be that growth of Pg was heavily and selectively affected by some sugars and polysaccharides. For example, those derived from traditional Chinese medicinal (TCM) fungi, such as Ganoderma lucidum and Hirsutella sinensis stimulated the growth (Lai et al, unpublished). Larger scale human investigation, especially those related to TCM polysaccharides administration, will be necessary to provide more in-depth information.
Previous can lead to intestinal inflammations and severe colitis,31 closely connected to impaired intestinal ribosomal-biogenesis.32 Furthermore, CS-related deregulated ribosomal biogenesis has also been linked to cellular senescence phenotypes33 and ageing, such as progeria and the Hutchinson-Gilford progeria syndrome.34 On the other hand, inhibition of mitochondrial respiratory activity was associated with pathogenesis of COPD,19 34 and might be the key mechanism of lung CS toxicity. Specifically, the mitochondrial dysfunction is linked to loss of RNA Pol I transcription, thus, compromising translational fidelity, and causing endoplasmic reticulum stress and apoptosis.34 Additionally, CS has been linked to subcutaneous fat losses that might lead to weight loss in CS mice35 (figure 1A,B), cachexia,36 atherosclerosis, alopecia, osteoporosis, and cataracts.37 Therefore, Pg MTS01 perturbation or restoration of aberrant colon ribosomal biogenesis and mitochondrial activity by CS-induced chronic inflammations may be more detrimental.
CS also systematically manifested aberrant amino acid-metabolism, including decreased protein anabolism and increased protein catabolism38 (online supplemental figure S6). Notably, Pg MTS01 systematically promoted amino acids turnovers, especially those involved in upregulated urea cycle, nitric oxide-biosynthesis and TCA cycle activities (online supplemental figure S6). The decreased metabolic turnovers of L-arginine and L-citrulline involved in mitochondrial TCA, argino-succinate shunt and urea cycle, and nitric oxide-biosynthesis activities (online supplemental figure S6) were restored by Pg MTS01 administration. While the Pg MTS01 effects on restoration of amino acids metabolism might be related to the enhanced cellular ribosomal and mitochondrial activities, more detailed molecular mechanistic studies are warranted.
Previous studies also have shown that the amount of IgG1 isotypes antibodies positively correlated with COPD pathogenesis and severity.39 Pg MTS01 administration did not significantly reduce the total IgG level in sera of COPD mice, although a weak trend was observed (online supplemental figure S7d). Effect of Pg MTS01 on other isotypes needs further investigations. At the same time, Pg MTS01 significantly reduced the expression level of scgb1a1 (online supplemental figure S7c) where bronchiolar cells derived from Scgb1a1-expressing progenitors function as major sentinel cells of the airway, responsible for controlling immune responses. Besides, concerning COPD-pathogenesis, particularly emphysema that was developed as the later stage of COPD, the role of B cells and B cell-rich lymphoid follicles have been highlighted.40 Specifically, increased IgA1-production B-cell counts were observed in lung lymphoid-follicles within distal lung parenchyma of COPD subjects.41 Apart from DCs and macrophages, B cells can internalise, process and present the antigen to responding T cells.40 Results shown in this study indicated that the over-activated B cells signalling pathways observed in lung of CS mice were significantly reduced by Pg MTS01 (figure 4G–I). Besides, expression levels of CD86 and proliferation in the B cells induced by EC-LPS were also antagonised by Pg-LPS (figure 5I,J). These results indicated the restoring effects of Pg MTS01 might be, at least in part, through modulation of B cells activities, and Pg-LPS may play some direct regulatory role on B cells. Whether the ameliorative mechanism of Pg MTS01/Pg-LPS in COPD lung may be dependent on interactions between B and T cells, and even other immune cells remains to be further determined.
Unexpectedly, scRNAseq analysis of lung of COPD animals revealed reductions in number of neutrophils and T cells relative to the control group (figure 4F), different from those obtained from cellular analysis (figure 1C). Neutrophils are central in COPD pathogenesis, with NETosis enriched in neutrophil-dominant refractory lung diseases as a cell death indicator.42 On the other hand, T cells also play important role in COPD inflammations,43 and a mean 103% increase in apoptosis in T-lymphocyte was found in COPD patients in contrast to normal population.43 Since scRNAseq mainly analysed live and intact cells, our observed phenomena might suggest that CS induced NETosis42 and apoptosis of T cells,44 leading to subsequent inflammations development. Treatment with Pg MTS01 increased the number of viable neutrophils and T cells, which may be closely related to its ameliorative effects.
The proinflammatory LPS mainly produced from Gram-negative Proteobacteria has been closely implicated in COPD development.45 Furthermore, cigarettes often contain proinflammatory LPS contaminants.46 The increased release of proinflammatory LPS in sera and BALF of CS mice may initiate and proceed with COPD development (figure 5A,B), and suggested the level of proinflammatory LPS activity is closely correlated with the inflammation severity. Pg-LPS reduced LPS activities in sera and BALF and downregulated genes involving CS-induced, and LPS-related inflammations, leading to alleviation of COPD-pathogenesis (figure 6J–O). Subsequent studies indicated that Pg-LPS did not significantly enhance cellular activation activity, and even antagonised the E. coli LPS induced activation activities (figure 5F–L). These results indicated the development of a potential new strategy on COPD treatment by using the anti-inflammatory LPS as therapeutic agents.25 The ameliorative effect of Pg-LPS was also examined in another CS COPD model of BALB/c mice which have been reported to have a very different composition of gut microbiome compared with C57BL/6 mice.47 The result revealed that Pg-LPS also ameliorates COPD development in CS BALB/c mice (online supplemental figure S10). Therefore, effects of Pg-LPS are not specific to C57BL/6 mice. While the underlying ameliorative mechanism of Pg-LPS might be through competing with EC-LPS for TLR4 receptor, further studies on modification of Pg-LPS to improve its anti-inflammatory efficacy are essential.
The general effects of hypoacylated lipid A on inhibition of immune activity have been reported before.25 Through genome comparison, the lpxM that might encode an acyltransferase during the processes of lipid A synthesis was found to be absent in Pg MTS01 genome (figure 5D,E). Therefore it was predicted that the structure of lipid A in Pg MTS01 was similar to those identified in Bacteroides spp such as Bacteroides thetaiotaomicron, Bacteroides dorei, Bacteroides fragilis, Bacteroides ovatus, Rhodobacter capsulatus and Rhodobacter sphaeroides, which is penta-acylated rather than hexa-acylated25 48 (figure 5E). These anti-inflammatory LPSs are generally characterised by showing no potent agonist activity, and even presenting antagonistic effects to E. coli derived proinflammatory LPS.23–25 For example, LPS derived from Rhodobacter demonstrated potent antagonism of proinflammatory LPS-mediated cellular activation and protected mice from proinflammatory LPS-induced death.49 50 Among these, E5564 developed form Rhodobacter sphaeroides was shown to prevent chronic airway hyperreactivity and inflammation caused by inhaled LPS. In addition, B. fragilis and B. ovatus also alleviated LPS-induced inflammation in mice,51 while B. vulgatus and B. dorei ameliorated endotoxaemia, decreased gut microbial LPS production, and suppressed proinflammatory immune responses.52 Recently, anti-inflammatory properties of weak agonistic LPS derived from Bacteroides spp were attributed to endotoxin tolerance induction via the MD-2/TLR4 receptor complex axis in intestinal lamina propria CD11c+ cells.53 Pg-LPS may, therefore, belong to a family of hypoacylated LPS that generally show anti-inflammatory functions.
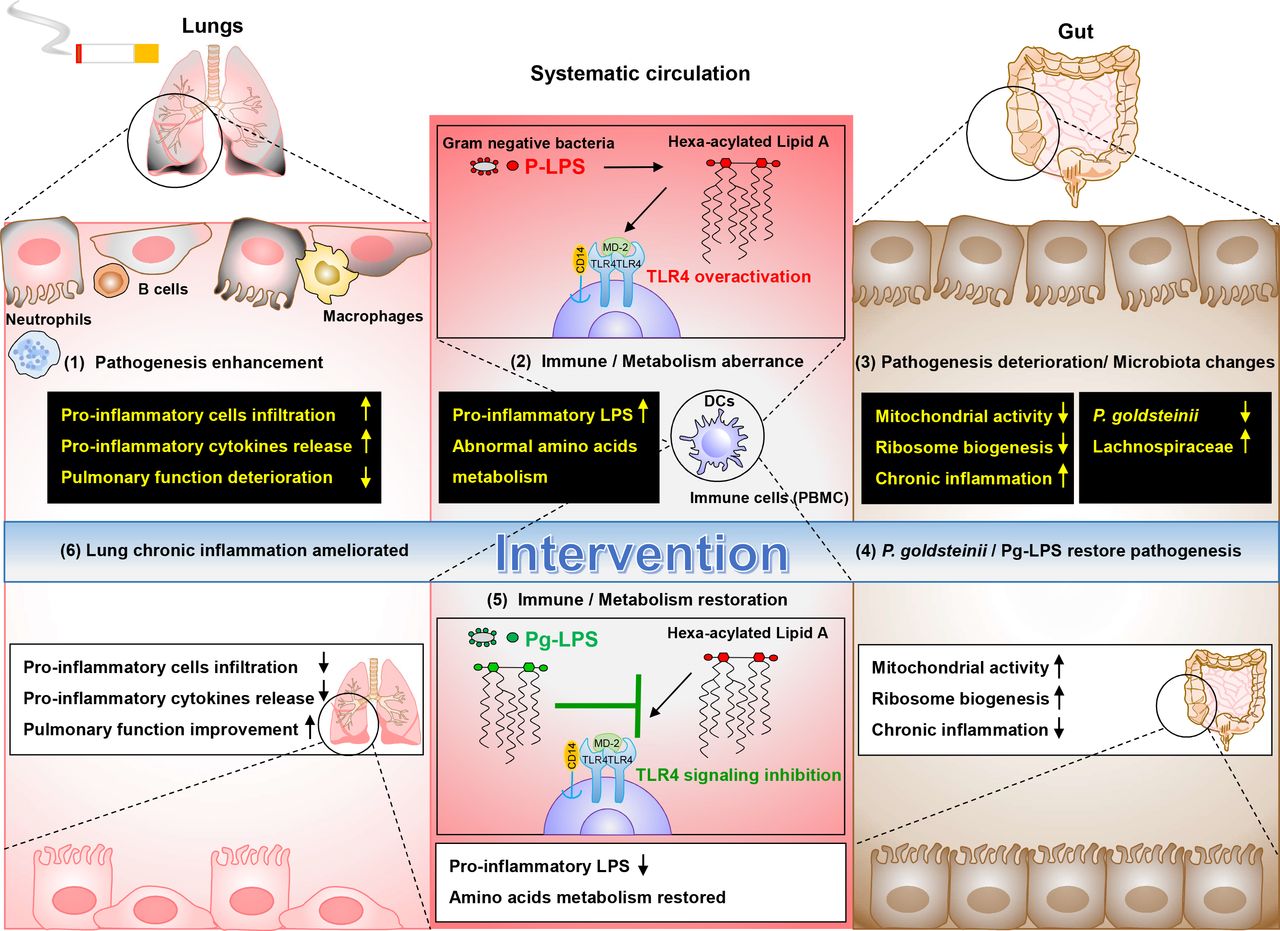
In conclusion, Pg MTS01 and Pg-LPS are potential therapeutic agents that may be developed into functional probiotics10 25 54 55 and postbiotics,25 respectively, to ameliorate COPD (figure 7).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Hypothetic working model of the ameliorative effects Parabacteroides goldsteinii (Pg) MTS01 and Pg-LPS on CS-induced COPD. In CS-induced COPD model, pathogenesis enhancement in lung such as increased infiltration of proinflammatory cells, increased release of proinflammatory cytokines and pulmonary function deterioration was observed (1). Increased pro-inflammatory LPS in systematic circulation was evident (2). These were closely related to aberrant systematic immune (TLR4 overactivation) and metabolic (amino acids metabolism disturbance) activities in the host. besides, CS also resulted in pathogenesis deterioration in colon, including reduced ribosome biogenesis, mitochondrial activities, and reduced ZO-1 and occludin production (3). Furthermore, microbiota changes were correlated with pathogenesis of COPD (abundance of Lachnospiraceae was positively correlated with disease severity, while abundance of Pg was negative correlated) (3). Interventions by administration of Pg MTS01 or its derived LPS (Pg-LPS) restored the pathogenesis in colon (4). Pg MTS01 might, at least in part, use its LPS by acting as an antagonist of TLR4 signalling pathway to restore the aberrant inflammatory and metabolic phenotypes (5). Consequently, amelioration of lung chronic inflammation, leading to reduced COPD pathogenesis was achieved (6). P-LPS, pro-inflammatory LPS. COPD, chronic obstructive pulmonary disease; CS, cigarette smoking; LPS, lipopolysaccharide; TLR4, toll-like receptor 4.
Data availability statement
Data are available on reasonable request. All data relevant to the study are included in the article or uploaded as online supplemental information. The data that support the findings of this study are available from the corresponding author, on reasonable request.
Ethics statements
Patient consent for publication
Ethics approval
Animal experiments were approved by the Institutional Animal Care and Use Protocol of Fu Jen Catholic University and were performed according to their guidelines (animal ethics approval numbers A10558 and A10849).
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Footnotes
H-CL and T-LL contributed equally.
Correction notice This article has been corrected since it published Online First. Figures 4 and 6 have been updated.
Contributors T-LL conceived the project, contributed to experimental design, performed experiments, interpreted the results, prepared the figures and wrote the manuscript; T-WC performed the scRNAseq analysis; Y-LK performed the microbiota analysis; C-JC and T-RW contributed to experimental design and performed experiments; C-CS, Y-HT and SS reviewed and edited the manuscript; H-CL and C-CL conceived and supervised the project, interpreted the results and wrote the manuscript; all authors discussed the results and approved the manuscript.
Funding This study was funded by CORPD1F0013 and CORPD1J0052 from Chang Gung Memorial Hospital, 108–2321-B-182–002, 109–2320-B-030–010, 109–2327-B-182–001 from Ministry of Science and Technology (MOST), Microbiota Research CenterCentre from Chang Gung University, and the Research CenterCentre for Emerging Viral Infections from The Featured Areas Research CenterCentre ProgramProgramme within the framework of the Higher Education Sprout Project by the Ministry of Education (MOE) in Taiwan and MOST, Taiwan (MOST109-3017-F-182–001).
Competing interests None declared.
Provenance and peer review Not commissioned; externally peer reviewed.
Supplemental material This content has been supplied by the author(s). It has not been vetted by BMJ Publishing Group Limited (BMJ) and may not have been peer-reviewed. Any opinions or recommendations discussed are solely those of the author(s) and are not endorsed by BMJ. BMJ disclaims all liability and responsibility arising from any reliance placed on the content. Where the content includes any translated material, BMJ does not warrant the accuracy and reliability of the translations (including but not limited to local regulations, clinical guidelines, terminology, drug names and drug dosages), and is not responsible for any error and/or omissions arising from translation and adaptation or otherwise.