Article Text

Abstract
Accumulating evidence indicates that the gut microbiome is an important regulator of body weight, glucose and lipid metabolism, and inflammatory processes, and may thereby play a key role in the aetiology of obesity, insulin resistance and type 2 diabetes. Interindividual responsiveness to specific dietary interventions may be partially determined by differences in baseline gut microbiota composition and functionality between individuals with distinct metabolic phenotypes. However, the relationship between an individual’s diet, gut microbiome and host metabolic phenotype is multidirectional and complex, yielding a challenge for practical implementation of targeted dietary guidelines. In this review, we discuss the latest research describing interactions between dietary composition, the gut microbiome and host metabolism. Furthermore, we describe how this knowledge can be integrated to develop precision-based nutritional strategies to improve bodyweight control and metabolic health in humans. Specifically, we will address that (1) insight in the role of the baseline gut microbial and metabolic phenotype in dietary intervention response may provide leads for precision-based nutritional strategies; that (2) the balance between carbohydrate and protein fermentation by the gut microbiota, as well as the site of fermentation in the colon, seems important determinants of host metabolism; and that (3) ‘big data’, including multiple omics and advanced modelling, are of undeniable importance in predicting (non-)response to dietary interventions. Clearly, detailed metabolic and microbial phenotyping in humans is necessary to better understand the link between diet, the gut microbiome and host metabolism, which is required to develop targeted dietary strategies and guidelines for different subgroups of the population.
- glucose metabolism
- nutrition
- obesity
- intestinal bacteria
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
Key messages
What is already known about this subject?
The interaction between the gut microbiome, diet and the host’s metabolism is multidirectional and complex, providing a challenge in acquiring detailed, mechanistic insights.
Gut-derived metabolites, including short-chain fatty acids (derived from fibre fermentation), may be important regulators of host metabolism. Most metabolites derived from protein fermentation seem to have detrimental effects on metabolic health.
What are new findings in this field and how could they impact future research and clinical practice?
The baseline gut microbial profile may be a predictor for an individual’s response to dietary interventions. Future research in this field should consider detailed characterisation of both microbial and metabolic phenotypes as well as their interaction.
Gut microbiome-associated effects on host metabolism may be related to fermentation products of carbohydrates and proteins. Understanding how to optimally balance proteolytic and saccharolytic fermentation and gaining insight into the importance of the site of colonic fermentation will gain insight into the interplay between diet, gut microbiome and metabolic processes.
Understanding the mechanisms of the differential responses to diet is essential to move forward in the field of precision nutrition. Although both the amount and quality of knowledge have evolved rapidly in recent years, we still only see the tip of the iceberg.
Introduction
The global rise in the prevalence of obesity is a major socioeconomic burden and is strongly associated with an increase in the incidence and prevalence of cardiometabolic diseases, including insulin resistance (IR) and type 2 diabetes mellitus (T2DM).1 2 Despite efforts to improve lifestyle choices and to increase insight in the underlying aetiology, long-term success in the prevention and treatment of obesity seems limited, as diet-induced weight loss is only maintained for approximately 25% after a 5-year follow-up.3 In recent years, advancements have been made in understanding the involvement of the gut microbiome in obesity and related cardiometabolic complications as regulator of host energy and substrate metabolism.4 5 Targeting host metabolism via the gut microbiome may therefore be a putative strategy to improve the effectiveness of lifestyle interventions to promote cardiometabolic health.
The complexity of the gut microbiome itself and its interplay with the host’s health and host energy and substrate metabolism, as well its role in obesity and T2DM, have become more evident.4 6–8 Despite an exponential increase in the number of studies investigating the impact of the gut microbiome on host metabolism over the past decade, data confirming a causal role of the gut microbiome in human metabolism are scarce and results are inconsistent.7 9 Moreover, findings of animal studies appear difficult to translate to humans due to differences in anatomy, genetics, physiology and gut microbiome–host interaction, among other factors.10 11 Throughout the 20th century, microbiome–host interactions have already gained scientific interest,12 providing the basis for human faecal transplant studies, which hint towards a causal role of the gut microbiota composition and its derived metabolites in the development of the metabolic syndrome.13 14 These studies have shown that baseline microbial composition may predict the improvement in peripheral insulin sensitivity in individuals with the metabolic syndrome, although effects were transient.14 Nevertheless, recently it has been demonstrated that strong alterations of the abundance of microbes induced by antibiotics had no significant effects on insulin sensitivity, inflammatory profile and energy/substrate metabolism in individuals with obesity and impaired glucose metabolism.15 This double-blind, randomised, placebo-controlled clinical trial indicates that modulation of microbiota composition may not have an important impact on human metabolism under all conditions, and that this impact rather depends on the individual’s characteristics such as age, habitual diet, metabolic phenotype and baseline gut microbial profile.
The composition of the gut microbiota is determined by hereditable, demographic and environmental factors, including mode of delivery at birth, age, sex, gastrointestinal transit time and use of medication. In recent studies, diet has emerged as a significant contributor to the shaping and defining of the gut microbiome.16–19 Diet-induced changes in the composition and functionality of the gut microbiota have been linked to the development of obesity and related disorders.20 Dietary fibres, in particular, have received great attention as potential mediators of the host’s metabolism via gut microbiome-related interactions,21 but conflicting findings have been reported regarding metabolic outcomes.22 23 As indicated previously, differences in baseline gut microbiota composition and functionality may determine the (lack of) response to an intervention.24 As such, it is tempting to postulate that interindividual variation in gut microbiota composition and functionality may serve as a basis for a more precision-based dietary approach in counteracting cardiometabolic diseases.
In this review, the complex interaction between dietary macronutrients, in particular dietary fibres and proteins, gut microbiome and host metabolic health, is addressed. The present review focusses on gut microbial metabolites produced in saccharolytic fermentation such as short-chain fatty acids (SCFAs) as well as proteolytic fermentation like ammonia, indolic and phenolic compounds, and branched-chain fatty acids (BCFAs) (figure 1). We will discuss the balance of saccharolytic and proteolytic fermentation as important determinant of metabolic health. It is increasingly evident that this balance may be modulated by diet in a person-specific or subgroup-specific manner.25 At first, it will be briefly addressed to what extent microbial composition and its functionality relates to the host’s metabolic phenotype, including the putative underlying mechanisms related to gut-derived saccharolytic and proteolytic metabolites. Subsequently, we will discuss the impact of dietary fibres and protein on microbial composition and functionality and insulin sensitivity, as well as to what extent response and non-response are mediated by baseline microbial composition. Overall, we will provide insight into the importance of precision nutrition strategies to modulate microbial composition and functionality and, thereby, bodyweight control and insulin sensitivity. Finally, we will address what is required to translate findings within this field into more precision nutrition-based interventions and guidelines.

Interactions between diet and saccharolytic and proteolytic fermentation in the gut and host metabolism. Fermentation of dietary fibres occurs mainly in the proximal colon and yields SCFAs that can both be used as fuel for enterocytes and can act as peripheral signalling molecules. SCFAs are involved in centrally regulating food intake and energy expenditure by effects on secretion of GLP-1 and PYY. SCFAs are beneficial regulators of interorgan crosstalk between the gut and peripheral organs like the liver and muscle. Protein fermentation mainly occurs in the distal colon and yields a more diverse range of metabolites, including BCFAs, which are associated with detrimental effects on gut and metabolic health. Green boxes indicate effects of SCFAs on metabolic processes in peripheral organs. Blue borders indicate effects in the opposite direction site direction (dotted line) or unknown direction (no line) of proteolytic fermentation products. BCFA, branched-chain fatty acid; FA, fatty acid; GLP-1, glucagon-like peptide 1; PYY, peptide YY; SCFA, short-chain fatty acid; TMAO, trimethylamine N-oxide.
Gut microbiome and host metabolic phenotype
Gut microbial composition in obesity and T2DM
The gut microbiome seems to impact the host’s metabolic phenotype through the production of different metabolites.26 Through these metabolites, the gut microbiota may affect immune function and epithelial integrity, energy and substrate metabolism, and glucose homeostasis.13 27 28 This capability of interacting with host health is dependent on the type of bacteria present and the distribution of these bacteria in the gut microbial community. Since functionality (eg, fermentation capacity) varies among bacterial species, alterations in gut microbiota composition and diversity, as observed in obesity and T2DM, may subsequently affect metabolic processes and the immune system of the host.
The core human gut microbiota mainly consists of the bacterial phyla Firmicutes, Bacteroidetes, Actinobacteria, Proteobacteria, Fusobacteria and Verrucomicrobia, of which Firmicutes and Bacteroidetes represent approximately 90% of the gut microbiota community.29 Ninety-five per cent of the Firmicutes phylum is represented by Lactobacillus, Bacillus, Clostridium, Enterococcus and Ruminococcus genera, while Bacteroidetes predominantly consists of Bacteroides, Prevotella and, to a lesser extent, Actinobacteria, which includes the Bifidobacterium genus.29 Although a core microbiota exists, its composition is highly individualised and shaped by genetics,30 sex,31 32 age,33 34 ethnicity35 and environmental factors, such as drug use28 36 and habitual diet.37 The uniqueness of the human gut microbiota profile translates into highly individualised responses to acute dietary challenges and longer-term dietary interventions.38–40 The success of an intervention might therefore be at least partially explained by baseline differences in the gut microbiome.
Interestingly, several studies demonstrated a relationship between the metabolic phenotype of an individual and the composition and functionality of the gut microbiota. Initial studies indicated an increased Firmicutes-to-Bacteroidetes ratio in humans and rodents with obesity as compared with lean individuals,41 42 but other studies have failed to observe such a difference or even reported a decreased ratio.9 43 44 More consistent, overweight and moderate obesity, IR and T2DM have been associated with compositional changes and decreased microbial richness and diversity when compared with lean, healthy individuals,41 45–47 and this has been linked to metabolic impairments. Indeed, detailed characterisation of individuals with overweight or (morbid) obesity showed that microbial gene richness is inversely correlated with fat mass, leptin levels, fasting insulin, homeostatic model assessment of insulin resistance (HOMA-IR), triglyceride levels and systemic inflammation.45 The prevalence of low microbial gene richness was found to be as high as 75% in severe obesity, compared with 23%–40% in lean or overweight/moderate obesity.4 48 In a comparison of metabolically healthy versus unhealthy individuals, alpha diversity was lower in the metabolically unhealthy group.49 Specifically, bacteria of the Oscillospira genus and Coriobacteriaceae family were associated with good metabolic health. In a study population encompassing people with normal weight and overweight/obesity, the abundance of specific genera correlated with metabolic characteristics.50 Notably, anthropometric parameters were positively associated with Collinsella aerofaciens, Dorea formicigenerans and D. longicatena, which had higher abundance in people with overweight/obesity. Moreover, different genera of Lachnospiraceae were negatively associated with body fat mass, low-density lipoprotein and total cholesterol concentrations. The involvement of individual genera or species in metabolic regulation seems to be confirmed by numerous other, mainly association studies in humans. Most compelling evidence is available for bacteria of Akkermansia genus, which is negatively associated with the risk of developing obesity and metabolic syndrome.51 52 A 3-month supplementation of Akkermansia muciniphila improved insulin sensitivity and reduced blood markers of liver dysfunction and inflammation in individuals with IR and overweight/obesity. Furthermore, another study showed significant enrichment of both A. muciniphila and Alistipes obesi in lean individuals, and of Ruminococcus gnavus in obese individuals. A 6-month calorie restriction diet increased A. muciniphila and A. obesi even more in lean individuals, while R. gnavus abundance decreased in the obese group.53 This study identified Blautia wexlerae and Bacteroides dorei as strongest predictors for weight loss when present in high abundance at baseline. Specific interventions with high-fibre diets, however, hint towards baseline Prevotella abundance as predictor for weight loss success on these diets in people with obesity.54
Additionally, patients with T2DM and metabolically compromised individuals showed an altered microbial functionality and a decreased fermentation capacity when compared with healthy individuals with, in particular, a lower abundance of butyrate-producing bacteria.13 55 56 Furthermore, the gut microbiome of insulin-resistant individuals may have increased biosynthesis potential and decreased uptake and catabolism of branched-chain amino acids (BCAAs, largely driven by Prevotella copri and B. vulgatus), which have been linked to detrimental metabolic effects.57 Taken together, there is evidence for decreased microbial gene richness and diversity in metabolically compromised individuals. Moreover, specific individual microbes may be regulators in (changes in) cardiometabolic health. Interindividual differences in gut microbiota composition and functionality may be linked to an altered responsiveness to (dietary) interventions, as will be discussed more extensively later in this review.
Saccharolytic and proteolytic fermentation, bodyweight control and insulin sensitivity
The interaction between the gut microbiota and host metabolism is multifactorial and depends on the microbial and host metabolic phenotype, which may be strongly inter-related. The production or activation of signalling molecules involved in host metabolism, the regulation of bile acid homeostasis, modification of gut permeability, the release of gut hormones, lipopolysaccharides and inflammatory markers, are among the mechanisms by which gut microbiota may influence the host cardiometabolic phenotype.58 Gut-derived metabolites may be either beneficial or detrimental to host metabolic health. In developing precision-based nutrition, the capacity of bacteria to produce certain metabolites should be considered. In this review, we focus on the balance between saccharolytic and proteolytic fermentations as a determinant of metabolic health (figure 1), as will be discussed in the following paragraphs.
Saccharolytic fermentation
The gut microbiota is capable of fermenting food components that otherwise would be unavailable to the host.59 Dietary fibres and other complex carbohydrates that cannot be broken down by enzymes in the small intestine can (partly) be fermented by bacteria in the large intestine as preferred energy source, producing microbial products such as SCFAs (predominantly acetate, propionate and butyrate).60 The main butyrate producing bacteria belong to the phylum Firmicutes, in particular Faecalibacterium prausnitzii, Clostridium leptum, Eubacterium rectale and Roseburia spp.61 The production of other SCFAs is mediated by bacteria such as Bifidobacterium that produce acetate and lactate during carbohydrate fermentation. Also, A. muciniphila species produce both propionate and acetate.61 62
Stable-isotope techniques with 13 C labelled SCFAs allow quantification of in vivo colonic production of SCFAs based on breath, urine and blood analyses.63 SCFAs are formed primarily in the colon, in which approximately 95% are subsequently absorbed.64 Butyrate is largely used as the major energy source by colonocytes, while propionate and acetate travel to the liver via the portal vein. In particular, acetate can also reach the peripheral tissues after entering the systemic circulation, inducing a diversity of metabolic and satiety-related effects.25 65 SCFAs can bind to G protein-coupled receptors (GPRs). The best studied receptors include GPR41, GPR43, GPR109a and GPR164, which are expressed in a vast array of cells, including the colonic epithelium, pancreatic β cells, immune cells and peripheral tissues, like adipose tissue.66 Effects of SCFAs on peripheral tissues include increased adipogenesis, inhibition of adipose tissue lipolysis (specifically by acetate) and attenuation of inflammation in adipocytes,65 67–69 increased lipid oxidative capacity in skeletal muscle,70 71 increased pancreatic insulin secretion and β-cell function,72 73increased insulin sensitivity and lipid oxidation in the liver74 75 and altered gut–brain interaction.76–79 These data are mainly derived from in vitro and rodent studies.
In human studies, however, data on associations between SCFAs and metabolic parameters cannot always be reproduced. For example, plasma insulin levels increased during hyperglycaemic clamps with acetate infusion independently of changes in plasma acetate in obese individuals.80 Beneficial effects of SCFAs may also be related to an increased secretion of intestinal incretins, such as glucagon-like peptide 1 (GLP-1) and peptide YY (PYY). Acute colonic administration of propionate increased postprandial plasma PYY and GLP-1 and reduced energy intake.81 Furthermore, long-term colonic propionate delivery prevented bodyweight gain, reduced abdominal obesity and intrahepatocellular lipid content, and prevented the deterioration in insulin sensitivity in overweight adults.81 In line with these findings, human in vivo data indicated that in adults with overweight or obesity, fasting lipid oxidation and resting energy expenditure increased after diet-induced changes in microbial SCFA production or direct colonic SCFA infusions.25 82–85 Although most studies indicate a beneficial role for SCFAs in energy and glucose homeostasis,65 86 87 larger human studies are needed to elucidate the complexity of underlying mechanisms.
Proteolytic fermentation
In a population consuming a Western diet low in dietary fibres, the microbiota of the distal part of the colon are more specialised in fermentation of residual peptides and proteins25 because the preferred fuel, fermentable carbohydrates, is already largely used in the proximal colon.88 Products of this proteolytic fermentation process include gaseous products such as hydrogen, methane, carbon dioxide and hydrogen sulfide; BCFAs isobutyrate, 2-methylbutyrate and isovalerate derived from fermentation of BCAAs, phenolic and indolic compounds derived from microbial fermentation of aromatic amino acids and to a lesser, unknown extent SCFAs.89 In contrast to saccharolytic fermentation products, most products derived from proteolytic fermentation are considered detrimental for host gut and metabolic health,60 89 90 although some data in animals suggest a beneficial role of indole and hydrogen sulfide in gut and peripheral tissue function.91 92
The balance between saccharolytic and proteolytic fermentation in the colon, and thus the balance between putatively beneficial and detrimental regulation of the host’s physiology, may be of interest in the development of dietary intervention strategies. Some studies showed that an increased dietary fibre intake, in particular intake of slowly fermentable fibres, reduces the production of merely detrimental proteolytic metabolites by the gut microbiota,24 93 suggesting a shift in overall fermentation balance towards the more beneficial saccharolytic fermentation.
Additionally, dietary choline and carnitine derived from, for example, red meat can be converted into trimethylamine (TMA) by microbial enzymes in the gut. This TMA reaches the liver via absorption and transportation via the portal vein. In the liver, TMA can be oxidised by hepatic flavin-containing monooxygenase into trimethylamine N-oxide (TMAO), which is strongly associated with the development of cardiovascular diseases, non-alcoholic fatty liver disease and systemic inflammation.94–96 Furthermore, in both murine and human studies, evidence is increasing that TMAO also contributes to the development of IR and T2DM.97–99 Increasing the availability of dietary fibres to the whole colonic microbiota might decrease the fermentation of dietary choline and carnitine, and therefore inhibit the production of the TMAO precursor TMA. Indeed, a rodent study demonstrated that a high amount of fermentable fibres derived from wheat bran lowered colonic TMA lyase activity.100 However, whether this can be translated to humans has to be elucidated.
Dietary macronutrients, microbial composition and metabolic health
Shaping of the adult gut microbiome is already initiated in early life, depending on factors such as exposure to the maternal microbiome, the mode of delivery and early exposure to dietary components,101 although the contribution of these factors cannot always be confirmed.102 Of the many factors influencing gut microbiota composition and function throughout all life stages, diet is key in modulating abundances of specific bacterial species and their functions.16 103 Vice versa, an individual’s response to a certain diet or dietary components may be largely influenced by features of the gut microbiota.39 This bidirectional interaction between diet and an individual’s microbiota forms a basis for understanding the concept of precision nutrition in the light of a third component: the host’s metabolic health.
Diet, macronutrients and microbial profile
In the current Western world, habitual diet composition has shifted towards a diet high in energy-dense foods, including relatively high amounts of saturated fat and simple carbohydrates, and low amounts of dietary fibre. Especially the consumption of dietary fibre, but also the macronutrient quality along with the quantity consumed can strongly influence gut microbiota composition and functionality. Population-based metagenomics analysis revealed associations between microbial composition and diversity and the habitual diet for over 60 dietary factors.These factors include energy and macronutrient intake and consumption of specific food items such as bread and soft drinks. These data confirm the importance of diet in shaping the gut microbiome.37 104
In a longitudinal monozygotic twin study, stool microbiota analysis showed that habitual intakes of energy, type of unsaturated fatty acid (FA) and soluble fibre affect microbiota composition, particularly the abundance of Bacteroides and Bifidobacterium.105 In the latter study, body mass index (BMI) did not correlate with microbiota composition. Furthermore, microbial enterotypes were strongly associated with long-term habitual diet, in particular protein and animal fat (Bacteroides) compared with carbohydrate intake (Prevotella).37 In line with this, long-term adherence to a Mediterranean diet was associated with specific taxa as well as functionality of the gut microbial profile.106 The findings of the latter study suggests that the composition of the gut microbiome is a modulator of the protective association between the Mediterranean diet and cardiometabolic disease risk.106 When comparing habitual high-fat diets to high-carbohydrate diets, microbial diversity seems to be lower with high-fat diets. Additionally, microbial diversity seems lower when comparing high (saturated)-fat diets to high carbohydrate/fibre diets.107–109 This diet-induced dysbiosis has been postulated as trigger of metabolic impairments in obesity.7 Importantly, evidence linking a high-fat, Western-type diet to the gut microbiota composition and diversity is mostly derived from observational rather than dietary intervention studies.110
Although primarily demonstrated in animal models, a limited number of human studies suggest that dietary intervention-induced alterations of microbial composition and functionality may already occur within weeks or even days after changes in dietary intake.111 In humans, modest microbial changes were found after a rigorous switch to a solely plant-based or animal-based diet.103 These rather extreme forms of dietary interventions provide insights into the mechanisms underlying diet–gut microbiome interactions and show that dietary intervention-induced microbial changes may occur very rapidly.59 In line with this, a small controlled-feeding study showed changes in microbiome composition within 24 hours of initiating a high-fat/low-fibre or low-fat/high-fibre diet, although enterotype identity remained stable throughout this 10-day study.37 These findings indicate that there is a tendency for microbial resilience in adults, which might be associated with long-term habitual dietary intake. The concept of bacterial enterotypes, however, has been questioned by several other studies due to the lack of understanding enterotype dynamics and resilience.112
Interestingly, a 1-year intervention study comparing the effects of an energy-restricted Mediterranean diet with increased physical activity to an isocaloric Mediterranean diet in overweight/obese adults showed marked differences in changes of the gut microbiota composition between the groups. Nonetheless, the tendency of microbial shift was in the same direction for both diets.113 Besides dietary macronutrient intake, other bioactive compounds may also affect gut microbiome composition and activity. Examples are dietary polyphenols,114 micronutrients, vitamins115 and probiotics116 and may contribute to the gut microbiota–host metabolism cross-talk. In the following paragraphs, we will highlight the effects of dietary macronutrient intake on the human gut microbiota and will link this to host metabolic outcomes. An overview of relevant human studies relating dietary macronutrient or dietary patterns to the gut microbiome is provided in table 1.
Effects of interventions with dietary fibre, carbohydrates and protein and changes in dietary patterns on gut microbiota and host metabolism in healthy, overweight, obese and insulin resistant individuals
Dietary fibres and metabolic health
Digestion of carbohydrates is a complex process involving specific enzymes for the type of carbohydrate ingested. The majority of digestible dietary carbohydrates are digested and absorbed in the small intestine, whereas certain, for a large part indigestible carbohydrates, including resistant starch and dietary fibres, are prone to fermentation by gut microbes with the highest content in the colon. Dietary fibres have been shown to have significant effects on the composition and functionality of the gut microbiota associated with health benefits.117–119 These dietary fibres may vary in their structural, physical and chemical properties such as water solubility, viscosity, binding and bulking ability, and fermentability.117 Highly fermentable fibres, such as β-glucan, inulin and galacto-oligosaccharides, are well defined in terms of effects on microbiota composition and production of gut metabolites,25 while insoluble fibres, although partially fermented, are mostly known for their beneficial effects on stool consistency and colonic transit time.120–122
The intake of high-fibre diets beneficially impacts the host’s health, among others by affecting glucose and lipid metabolism. Important mechanisms include the regulation of nutrient absorption123 or the production of SCFAs,65 but data on the health effects of dietary fibres are controversial. There are several explanations for the inconsistent findings in dietary fibre studies.25 65 First, in the majority of human studies, only one specific fermentable fibre was supplemented, thereby stimulating only one or a few individual (potentially beneficial) bacteria genera.22 65 The consequence of the latter can be a decreased abundance of other essential bacteria, which may result in an imbalance of the microbial ecosystem. Therefore, combining different fibres that stimulate a multitude of different bacterial genera may be important to maintain microbial richness with more pronounced (additive or synergetic) effects on immune status and metabolic health.25 Interestingly, a mechanistical study indicated that the site of fibre-derived SCFA production in the colon may be a determinant of metabolic health. The latter study showed that acute distal colonic acetate administration increased circulating acetate concentrations and increased fat oxidation, and satiety-stimulating hormone PYY, and reduced plasma tumour necrosis factor-α in men with overweight.83 In contrast to distal infusion, acetate administration in the proximal colon did not affect metabolic profile.83 Thus, targeting dietary fibre availability and SCFA formation by microbial species in the distal colon, by combining different dietary fibres and/or more complex dietary fibres, may be a promising strategy to improve immune and metabolic health.
Second, most dietary fibre intervention studies, so far, did not take baseline microbiome or metabolic phenotype into account. Characteristics of the gut microbiome at baseline may strongly interact with dietary intervention outcome. For example, it has been shown that the response of the gut microbiota to dietary fibres (resistant starch vs non-starch polysaccharides) can be predicted from the baseline microbial diversity in men with obesity.124 In the latter study, high microbial diversity was associated with lower dietary responsiveness of the microbiota,124 which may support the hypothesis that a higher diversity in gut microbes is associated with stability of the microbial ecosystem. In line with this, a high microbial gene count at baseline was related to a less pronounced response to weight loss diets as compared with individuals with a low gene count. In the low gene count group, there was an improvement in gene richness and clinical parameters, although changes in inflammatory markers were less pronounced in individuals with low gene richness.48 The basis of the non-response in the high-gene count group is not clear, as this was not associated with habitual dietary intake. Additionally, another study in individuals with obesity showed that not the baseline microbial diversity but rather the baseline abundance of Firmicutes predicted the dietary response of an individual’s microbiota.125 Together, these findings indicate that microbial diversity is not always a predictor of dietary responsiveness and imply that further research is needed to better understand the complex diet–microbiome–host metabolism interactions.
The efficacy of dietary fibre interventions may also depend on habitual diet and fibre intake. Healthy individuals with high habitual dietary fibre intake had greater changes in gut microbiota composition as a response to an inulin-type fructan prebiotic compared with people with low habitual fibre intake whose gut microbiota seemed to be more resilient to change.126 A study conducted in patients with T2DM revealed that a select group of SCFA-producing microbial strains was promoted by dietary fibres and that many other microbes, including those in proteolytic fermentation, were either diminished or unchanged, showing an overall decreased microbial gene richness.24 Faecal SCFAs increased, in particular butyrate, which was accompanied by improvements in glucose homeostasis. Thus, greater microbial gene richness might not be beneficial per se, as shown in several human fibre dietary intervention studies,24 but physiological outcomes may rather be dependent on the functionality of the microbial network.
In a study investigating the effects of a 6-week whole-grain diet on bodyweight changes, a high baseline abundance of bacteria of the Prevotella genus correlated with higher weight loss in overweight, healthy adults.54 These findings indicate a predictive capacity as effect modifier of the gut microbiota in response to specific dietary interventions. Additionally, it was found that overweight, prediabetic individuals had a reduced response with respect to changes in microbial diversity and postprandial insulin sensitivity to short-term administration of long-chain inulin combined with resistant starch as compared with lean individuals.86 In line with this, recent studies have shown that baseline gut microbial signatures may predict the change in BMI following 3 months’ long-chain inulin supplementation,127 an effect that was replicated in hum-ob mice colonised with the faecal microbiota from different individuals with obesity at baseline.
Overall, the effectiveness of prebiotic dietary fibres on metabolic health outcomes may depend on several parameters, including initial microbial composition as well as the site of microbial fermentation.
Dietary protein, microbiota composition and metabolic health
Ingested dietary proteins are first digested in the small intestine by pancreatic enzymes and peptidases from enterocytes. A significant amount of oligopeptides and amino acids is then transported to the portal bloodstream via enterocyte transporters where they are used as amino acid precursors for protein synthesis or metabolised for fuels or precursors necessary for intestinal mucosal metabolites.88 128 129 Since fermentable carbohydrates are preferentially used over proteins by most bacteria in the distal small intestine and proximal colon, most fermentation of amino acids as an energy source occurs in the distal colon, where carbohydrates are depleted.130 The percentage of ingested protein reaching the large intestine is likely also dependent on protein quality, with estimated numbers around 10%.131 Protein originating from plants have a lower digestibility due to the indigestible cell wall of the plant,132 while protein derived from animals can be more easily digested in the large intestine, indicating potential difference in functional outcomes. Metabolites resulting from proteolysis (figure 1) can be either used by colonocytes or excreted in faeces.128 Most proteolytic products are considered to have detrimental effects on gut and host metabolic health.89 Furthermore, the balance between proteolytic and saccharolytic fermentation may determine the non-response to lifestyle interventions133 and should therefore be considered in future research.134
Metabolic effects of protein intake have been well studied. Casein, a relatively slowly digestible protein derived from animal products, was the most efficient protein source preventing weight gain and accretion of adipose mass in mice on a high-fat/high-protein diet (HPD).135 Epidemiological studies also indicate that intake of dairy and vegetarian protein sources is associated with protection against obesity, whereas a high intake of meat (in particular red meat) predicts higher weight gain.136–138 Although investigated less extensively, protein intake has been shown to affect microbiota composition and functionality.139 Effects are determined by the amino acid composition and digestibility of proteins, which are influenced by the source and the amount of intake.140 High protein diets have been linked to reductions in C. coccoides, C. leptum and F. prausnitzii in rats,141 while reductions in Roseburia spp, E. rectale, C. aerofaciens, Bacteroides spp and Oscillibacter relatives have been observed in men with overweight or obesity.89 142 Of note, comparing HPDs to moderate-protein or low-protein diets in an isocaloric manner induces differences in either carbohydrate or fat content between diets. As for all isocaloric dietary macronutrient exchange studies, it is therefore difficult to determine the predominant dietary factor responsible for compositional shifts in the gut microbiota, which could be attributed to either an increase in one (macro) nutrient or a decrease in the other.
Although proteolytic metabolites are predominantly associated with adverse health effects, current evidence also suggests that proteolytic metabolites may have beneficial effects on metabolic health. For instance, several amino acid-derived compounds that can be produced only by gut bacteria (indole) or by mammalian host (tyramine, tryptamine and SCFAs) (in)directly impact satiety and gut motility in mammals via effects on GLP-1 and serotonin secretion from enteroendocrine cells.143 However, the physiological effect on the host intestinal and peripheral tissues of the majority of these compounds is still not well understood. In addition, the distinction between human and bacterial origins of many of these compounds has not been well established yet, and further in vivo studies are necessary to validate such effects.
Dietary fats and polyphenols and interaction with the gut microbiome
Dietary fats have been studied extensively in relation to diet-related metabolic diseases such as obesity, but the effects on the gut microbiota are less defined in humans, and studies often yield opposite findings.144 FAs differing in type (saturated vs monosaturated and polyunsaturated FAs), carbon chain length and the degree of saturation may have a distinct effect on the gut microbiota composition.109 Cross-sectional studies indicate that consumption of a diet rich in animal protein and fat was associated with the Bacteroides enterotype, while higher-fibre, fruit and vegetable consumption was associated with the Prevotella enterotype in healthy adults.145 Furthermore, a high intake of mainly saturated fatty acids (SFAs) has been associated with a decrease in gut microbial richness and diversity in both adults and infants.146 147 In people with overweight and obesity, SFAs were negatively associated with the genus Intestinimonas, whereas SFAs were positively associated with Roseburia, which was also highly abundant in individuals with normal body weight.50 In this study, habitual SFA intake exhibited opposite association profiles with butyrate-producing bacteria, depending on the BMI. Overall, it should be noted that dietary fat–microbiome–host physiology interactions are studied less extensively compared with dietary fibre, and that the mechanistic knowledge is largely based on animal studies. Interpretation of these findings in light of human physiology is difficult and should be investigated further.
Second, out of a wide array of dietary components, dietary polyphenols and their effect on the gut microbiome and host metabolism have been studied extensively in recent years. Polyphenols, present as phenolic compounds in mainly fruits and vegetables, are known for their beneficial effects as antioxidant, anti-inflammatory, cardioprotective, cancer preventive and neuroprotective agents.148 A 2-month epigallocatechin-3-gallate supplementation, naturally present in tea, had both an effect on bile acid metabolism and Verrucomicrobiaceae abundance in mice with obesity, under which it promoted the increase of A. muciniphila abundance.149 The latter has been associated with beneficial metabolic effects in other studies.52 150 151 Additionally, although also in an animal model, 8-week polyphenol supplementation protected from diet-induced obesity, IR and intestinal inflammation, in association with increased abundance of Akkermansia spp.114 In healthy, overweight or obese individuals, 12-week combined resveratrol and epigallocatechin-3-gallate supplementation improved both metabolic parameters and decreased Bacteroidetes in men but not in women.152 Overall, adding dietary polyphenols to a diet seems to promote both gut and metabolic health, although mechanistical insights in human studies are still required.
Towards precision nutrition: conclusions and future perspectives
Microbiome-based precision nutrition to predict metabolic health parameters like glycaemic response and variability, or used to counteract metabolic disarrangements has currently received major scientific attention. A landmark study within this field showed that, despite high interpersonal variability in postprandial glucose response, personalised diets (based on habitual diet, physical activity and gut microbiota) created with the help of a machine-learning algorithm may successfully lower blood glucose responses.39 The postprandial predictive capacity of the gut microbiome to meal challenges was again highlighted in another study, showing that an individual’s microbiome had a greater influence (explaining 7.1% of variance) than meal macronutrients (explaining 3.6% of variance) for predicting postprandial lipaemia, but not for postprandial glycaemia (explaining 6.0% and 15.4% of variance, respectively).38 In line with this, significant interpersonal differences were found in glycaemic response to different types of bread, and this glycaemic response could be predicted by baseline microbiome features.153 Of note, these studies mainly based their findings on responses to acute meal challenges and short-term interventions rather than on longer-term intervention responses. Conclusive results for the latter is still scarce, but the predictive capacity of the gut microbiome is becoming more evident, especially in studies examining the effects of fibre and faecal microbiota transplantation.13 14 40
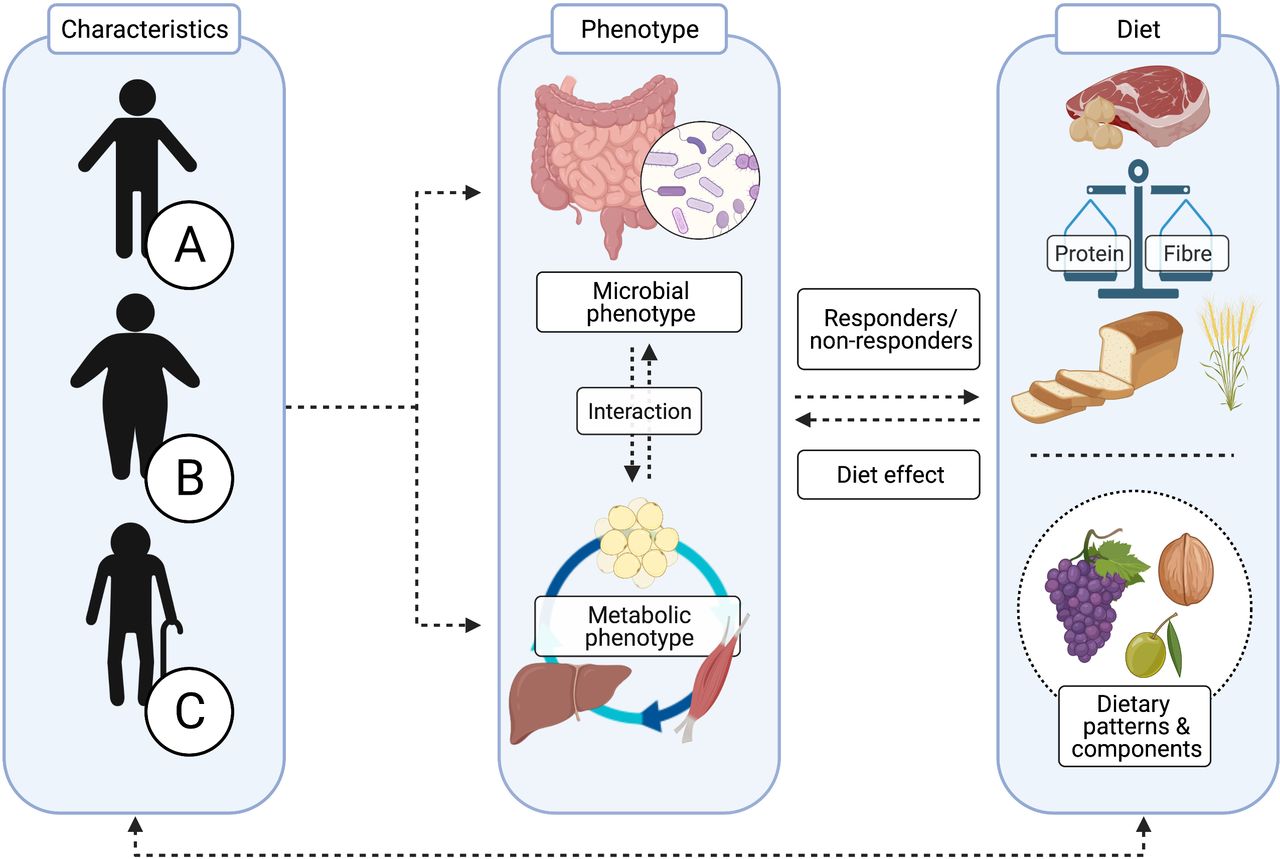
As discussed previously, there may be putative resistance of the microbiome–host metabolism axis to dietary intervention in individuals with IR, suggesting that interventions may require longer periods of time or that higher intake of functional dietary components such as dietary fibre is needed to induce beneficial effects. In this respect, the baseline microbial profile is an interesting biomarker for responsiveness to dietary interventions (eg, dietary fibre or complex proteins). Specific functional microbial groups are characterised by distinct digestive capacities for dietary components, resulting in a differential production of microbial metabolites such as SCFAs, subsequently impacting regulation of the host’s metabolism.154 Several studies identified a predictive capacity for a Prevotella enterotype to optimise weight loss in response to a high-fibre diet, while this was not found in individuals with a Bacteroidetes enterotype.40 54 155 The effectiveness of administering optimal fibre types in individuals stratified for microbial composition and functionality, degree of diversity or abundance of specific microbial strains has not been examined yet; therefore, prospective evidence is required in future dietary intervention studies. Overall, when evaluating dietary patterns and diets varying in macronutrient composition, it is important to take both quantity as well as quality of dietary components, micronutrient as well as bioactive components like polyphenols into account—within a whole-diet approach—due to differential interaction with the host’s microbial and metabolic phenotype (figure 2).

{kind=link}
{kind=link}
Applying optimal strategies in precision nutrition requires detailed characterisation of the individual. The composition and functionality of the gut microbiota are influenced by many factors, including genetics, age, sex and environmental factors such as the mode of delivery, drug use, disease, geography, physical exercise and diet.19 The multidirectional interaction between the gut microbiota, host metabolism and diet is therefore complex and highly individualised. When developing more targeted dietary strategies for individuals A, B and C, the microbial and metabolic phenotypes (eg, tissue-specific insulin resistance), as well as characteristics of dietary components (including the balance between protein and carbohydrate (CHO)/fibre intake) should be well considered.
The responsiveness to dietary interventions does not solely depend on characteristics of the gut microbiota but is rather dependent on a complex, multifactorial interaction between diet, lifestyle and environmental factors and clinical characteristics such as the metabolic phenotype (figure 2).16 156 157 It was recently shown that men and women differ in microbiota composition and that this composition is associated with insulin sensitivity in men but not in women.158 Interestingly, another study also demonstrated that 12-week combined polyphenol supplementation (using epigallocatechin-gallate (EGCG) and resveratrol) affected the gut microbiota composition in men but not in women.152 Both studies indicate a sex-specific microbial response, which should be considered when evaluating intervention responses.
In a study investigating metabolic responsiveness to specific diets in different metabolic phenotypes, the presence of either more pronounced muscle IR on more pronounced hepatic IR at baseline was a determinant of the change in disposition index (a composite marker of insulin sensitivity and insulin secretion) after 2 years of either a Mediterranean or a low-fat diet.159 Individuals with muscle IR responded better to the Mediterranean diet, while individuals with hepatic IR were more responsive to the low-fat diet.159 This study illustrates the importance of considering individual metabolic characteristics when developing precision nutrition strategies. To which extent the gut microbiome is involved in these types of dietary interventions remains to be investigated. Interestingly, the soluble fibre inulin has been shown to reduce IR in people with impaired fasting glucose but not in those with impaired glucose tolerance. The latter findings may suggest tissue specificity in the fibre–gut microbiota–host metabolism crosstalk, in view of the findings that impaired fasting glucose is strongly related to hepatic IR.160 Of note, targeting metabolically impaired individuals indeed requires both detailed characterisation of an individual’s microbial and metabolic phenotyping, as well as lifestyle factors. A previous study has shown that associations between microbial taxa and tissue-specific insulin sensitivity might be cohort-specific, suggesting alternative states of dysbiosis, depending on cohort characteristics. This further highlights the complexity of interpretation and generalisation of findings in the gut microbiome field.161
In order to translate precision-based strategies into healthcare practice or into guidelines, we need a thorough understanding of why people respond differentially to diet, whether differential responses and related phenotypes are maintained in the long term and to what extent developed algorithms are reproducible. In our view, detailed microbial and metabolic phenotyping by state-of-the art methodologies in dietary intervention studies is crucial. Obviously, in view of the complexity, we need besides that detailed information about lifestyle and environmental factors, including mode of birth, medical history, use of medication, in particular antibiotics, physical activity, psychological stress and sleep quality. This also implies that we need advanced statistical and modelling methodologies to tease out the importance of different factors. Importantly, however, we certainly must go beyond a black-box machine-learning prediction of dietary intervention responses.
Overall, we conclude that current evidence for developing optimal dietary interventions targeting bodyweight control and IR via the gut microbiota is still in its infancy and does not capture the complexity of the integration of a whole-diet approach, the microbial and the host’s metabolic phenotype (figure 2). Implementation of targeted, precision nutrition intervention strategies or dietary guidelines for individuals or subgroups in public health is still more remote and will require insight in the mechanisms involved in (non-)response to dietary intervention, implying that we need to go beyond prediction models. Detailed individual phenotyping and gaining insights into the balance between carbohydrate and protein fermentation by the gut microbiota as well as the site of fermentation in the colon are thereby key.
Data sources
A semisystematic search in PubMed was used to select relevant literature. Literature was managed using EndNote V.X8. The search strategy consisted of one or a combination of the following search terms using the “AND” and “OR”operators:
“Insulin resistance”[MeSH], “obesity”[MeSH], “gut microbiota”[MeSH], “type 2 diabetes”[MeSH], “short-chain fatty acids” [TIAB], “lps”[TIAB], “glucose metabolism”[TIAB], “”[TIAB], “impaired fasting glucose”[TIAB], “impaired glucose tolerance”[TIAB], “energy metabolism”[Mesh], “gut-brain axis”[TIAB], “PYY”[TIAB], “GLP-1”[TIAB], “peripheral insulin resistance”[TIAB], “insulin”[TIAB], “gut metabolites”[TIAB], “enterotypes”[TIAB], “gut microbiota composition”[TIAB], “dietary proteins”[MeSH], “dietary carbohydrates”[MeSH], “dietary fiber”[MeSH], “dietary fat”[MeSH], “diet”[MeSH], “dietary patterns”[MeSH]. From the identified articles, the titles and abstracts were assessed, and if considered relevant for the present review, the full text of the article was examined in detail. In addition, additional records were identified through searching reference lists. Only articles written in English were included.
Ethics statements
Patient consent for publication
Ethics approval
This study does not involve human participants.
Acknowledgments
The authors acknowledge BioRender.com as the tool for creating the figures in this article.
Footnotes
Correction notice This article has been corrected since it published Online First. The author affiliations have been updated.
Contributors KMJ wrote the manuscript; EEC, GHG and EEB conceptualised, reviewed and edited the manuscript. All authors read and approved the final version of the manuscript.
Funding The authors performed their work within a project organised by and executed under the auspices of TiFN, a public–private partnership on precompetitive research in food and nutrition (project code: 16NH04). Funding for this research was obtained from the Netherlands Organization for Scientific Research, DSM, FrieslandCampina, Danone Nutricia Research, and the Top Sector Agri & Food the Netherlands.
Competing interests None declared.
Patient and public involvement Patients and/or the public were not involved in the design, conduct, reporting or dissemination plans of this research.
Provenance and peer review Not commissioned; externally peer reviewed.