Article Text

Abstract
Objective Although gut dysbiosis is increasingly recognised as a pathophysiological component of metabolic syndrome (MetS), the role and mode of action of specific gut microbes in metabolic health remain elusive. Previously, we identified the commensal butyrogenic Anaerobutyricum soehngenii to be associated with improved insulin sensitivity in subjects with MetS. In this proof-of-concept study, we investigated the potential therapeutic effects of A. soehngenii L2-7 on systemic metabolic responses and duodenal transcriptome profiles in individuals with MetS.
Design In this randomised double-blind placebo-controlled cross-over study, 12 male subjects with MetS received duodenal infusions of A. soehngenii/ placebo and underwent duodenal biopsies, mixed meal tests (6 hours postinfusion) and 24-hour continuous glucose monitoring.
Results A. soehngenii treatment provoked a markedly increased postprandial excursion of the insulinotropic hormone glucagon-like peptide 1 (GLP-1) and an elevation of plasma secondary bile acids, which were positively associated with GLP-1 levels. Moreover, A. soehngenii treatment robustly shaped the duodenal expression of 73 genes, with the highest fold induction in the expression of regenerating islet-protein 1B (REG1B)-encoding gene. Strikingly, duodenal REG1B expression positively correlated with GLP-1 levels and negatively correlated with peripheral glucose variability, which was significantly diminished in the 24 hours following A. soehngenii intake. Mechanistically, Reg1B expression is induced upon sensing butyrate or bacterial peptidoglycan. Importantly, A. soehngenii duodenal administration was safe and well tolerated.
Conclusions A single dose of A. soehngenii improves peripheral glycaemic control within 24 hours; it specifically stimulates intestinal GLP-1 production and REG1B expression. Further studies are needed to delineate the specific pathways involved in REG1B induction and function in insulin sensitivity.
Trial registration number NTR-NL6630.
- diabetes mellitus
- intestinal bacteria
- duodenal mucosa
- gut hormones
- gene expression
Data availability statement
All data relevant to the study are included in the article or uploaded as supplementary information.
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
Significance of this study
What is already known on this subject?
Intestinal microbiota shapes host metabolic fitness and gut dysbiosis is associated with metabolic disorders.
Intestinal Anaerobutyricum soehngenii levels associate with enhanced insulin sensitivity in subjects with metabolic syndrome (MetS).
Oral A. soehngenii L2-7 supplementation improves insulin resistance in diabetic obese db/db mice.
Significance of this study
What are the new findings?
A single-dose of duodenal infusion containing A. soehngenii L2-7 is sufficient to improve peripheral glucose metabolism in human MetS.
A. soehngenii L2-7 stimulates the secretion of the incretin hormone GLP-1, which is reported to act on both insulin secretion and sensitivity.
Treatment with A. soehngenii specifically upregulates duodenal expression of regenerating islet-protein (REG)1A/B.
Duodenal REG1B expression associates with improved glycaemic control and GLP-1 levels.
Delivery of A. soehngenii into small intestine is shown, for the first time, to be safe and well tolerated.
How might it impact on clinical practice in the foreseeable future?
This study provides new insights into how intestinal microbes can affect host metabolism via duodenal signals.
Duodenal engraftment by multiple A. soehngenii administrations may be a novel treatment against insulin resistance in obesity and type 2 diabetes.
Introduction
Along with lifestyle, diet and visceral obesity, the gut microbiota constitutes an important modulator of metabolic heath.1 Accumulating evidence has now established that intestinal microbiota–diet–host interactions shape host metabolic homeostasis2–4; particularly, an unbalanced gut microbiome is increasingly recognised as an important risk factor for metabolic disorders, such as obesity, insulin resistance and type 2 diabetes (T2D).3–5 In the pursuit of novel therapeutic insight for the rising cardiometabolic disease burden, the development of culture-independent approaches using high-throughput sequencing has tremendously advanced our knowledge of the microbial signatures of obesity and T2D.6 General traits of obesogenic microbiota include a decline in faecal microbial community diversity, constriction of species richness and deprivation in short-chain fatty acid (SCFA)-producing microbes.7 8 Nonetheless, the exact mechanisms by which specific bacterial strains regulate metabolic functions and influence the pathophysiology of metabolic disorders in humans are still poorly understood.
In an attempt to move from association to causality, we previously conducted repetitive faecal microbiota transplantations (FMTs) in humans to gain insights into the microbiome-derived effects on glucose and lipid metabolism. We proved that transfer of healthy microbiota from lean donors into patients with metabolic syndrome (MetS) improved their peripheral insulin sensitivity3 4 and discovered that the latter was associated with increased relative abundance of Anaerobutyricum spp, including Anaerobutyricum soehngenii, in the small intestine (SI) following lean donor FMTs.3
A. soehngenii (formerly classified Eubacterium hallii) strain L2-7 is an anaerobic Gram-positive, catalase-negative bacterium belonging to the Lachnospiraceae family of the phylum Firmicutes.9 This strain is capable of converting sugars as well as lactate and acetate into the SCFA butyrate,10 which was shown to exert beneficial effects on glucose metabolism in obese mice and lean humans,11 12 thus underscoring a potential therapeutic benefit of intestinal A. soehngenii. We previously described the efficacy of oral A. soehngenii L2-7 supplementation in improving insulin resistance and energy expenditure in diabetic and obese db/db mice.13 Moreover, in a phase I/II safety and dose-finding trial, we showed that daily oral intake of A. soehngenii L2-7 for 4 weeks is safe and well tolerated and disclosed a positive correlation between faecal A. soehngenii L2-7 abundance and whole-body glucose rate of disposal.14 However, a major disadvantage of oral administration of bacterial strains is the loss of viability due to contact with oxygen and stomach acid. Therefore, in the present study, duodenal tube infusion was chosen to bypass this issue and, hence, maximise the therapeutic potential by delivering viable bacteria directly into the SI, the first anatomical site with a central role in glucosensing, regulation of peripheral insulin sensitivity/secretion and glucose homeostasis.15
In this regard, within the intestinal milieu, SI enteroendocrine cells act as ‘chemo sensors’ of diet-derived and microbiota-derived metabolites, such as butyrate, and can regulate host glucose metabolism by secreting a variety of hormones, such as the incretin glucagon-like peptide-1 (GLP-1), which in turn enhances both insulin secretion and sensitivity.15–17 However, the impact of single bacterial strains on the human enteroendocrine system remains elusive.
We therefore performed a randomised double-blind placebo-controlled cross-over trial to determine the localised and systemic effects of a single duodenal infusion of A. soehngenii L2-7 in male subjects with MetS. Our primary objective was to characterise the immediate changes induced by A. soehngenii L2-7 in the SI transcriptomic profile (6 hours postinfusion). Secondary objectives consisted in investigating the effects of A. soehngenii L2-7 on circulating (postprandial) incretins, faecal SCFA rates as well as gut microbiota composition.
Results
Baseline characteristics and safety parameters
We included 15 Caucasian treatment-naïve men with MetS. During the trial, three subjects were excluded (two subjects refrained from participation after the screening due to personal reasons, and one subject was excluded because of antibiotic use), and thus 12 subjects were left for primary endpoint analyses. Baseline characteristics are presented in table 1. Participants were randomised to receive either 10% glycerol infusion (placebo, n=6) or 1011 cells of A. soehngenii L2-7 in 10% glycerol (treatment, n=6) as first intervention, and switch to treatment/placebo 4 weeks later (figure 1). Both infusions were well tolerated and no (severe) adverse events occurred during the entire study. Safety laboratory parameters (inflammatory, kidney and liver parameters) were all stable during the study. Energy and macronutrient intake did not differ in the week after A. soehngenii L2-7 or placebo administration (online supplemental table S1). We also observed no differences in body weight, blood pressure, glucose, insulin, homeostatic model assessment of insulin resistance (HOMA-IR) or cholesterol levels between placebo and A. soehngenii L2-7 treatments (online supplemental table S1).
Supplemental material

Study overview. Schematic representation of the study design showing the time points of interventions and of biological samplings: all 12 subjects received placebo (10% glycerol in PBS) or treatment (Anaerobutyricum soehngenii L2-7) at week 0 or 4 (time of intervention cross-over). FSL, FreeStyle Libre System; SCFA, short-chain fatty acid.
Baseline characteristics at screening
Single-dose of A. soehngenii L2-7 has no impact on microbiota richness/diversity nor fecal SCFA
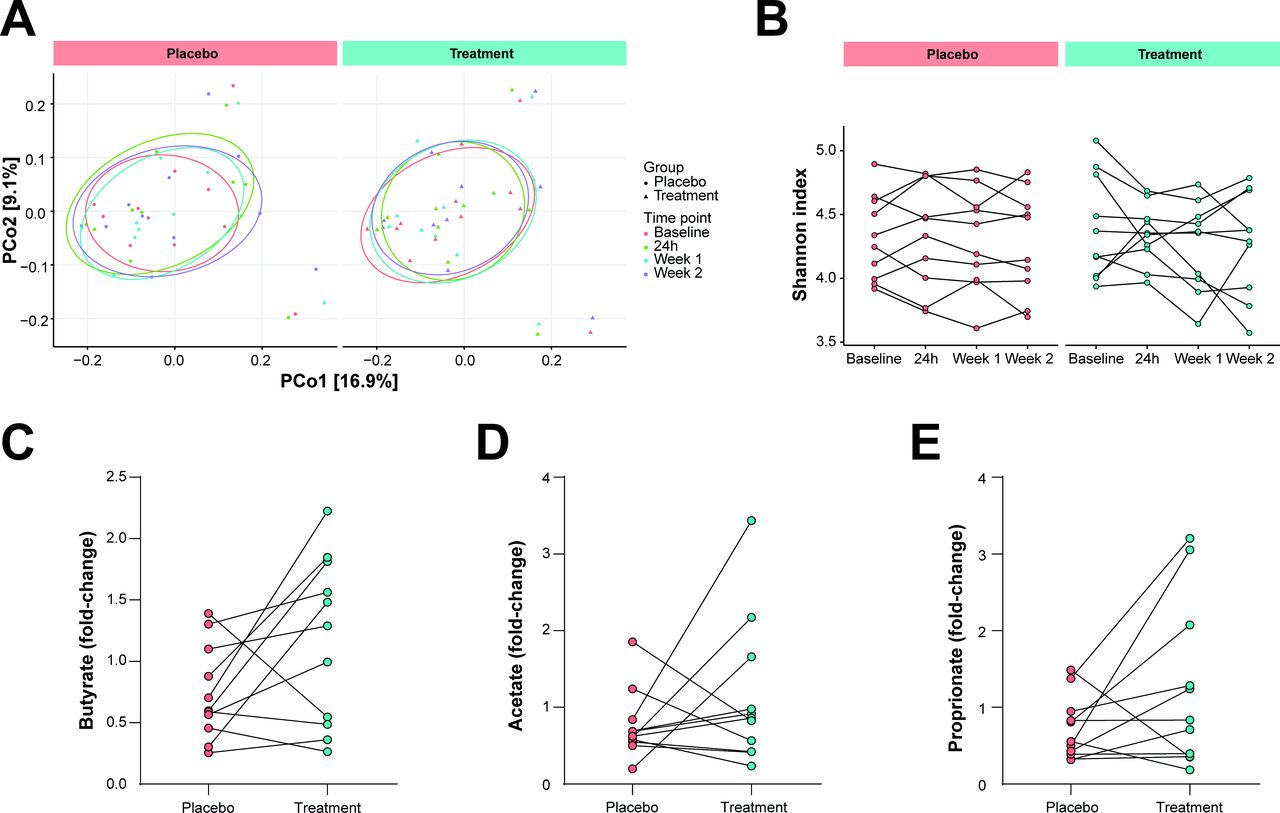
To discern the impact of A. soehngenii L2-7 infusion on gut microbial communities, 16S rRNA gene amplicon sequencing was performed using faecal DNA extracted from stool samples collected at baseline, 1 day, and 1 and 2 weeks after interventions. A single infusion of A. soehngenii L2-7 did not affect gut microbiota composition or alpha-diversity (Shannon index) either in the short term (24 hours) or long term (2 weeks) after intervention (figure 2A,B). Similarly, the abundance of A. soehngenii L2-7 in faecal samples, assessed by qPCR, was not durably altered over time (online supplemental figure S1). Notably, these data exclude microbiota-mediated carry-over effects at the time of cross-over (4 weeks after first intervention). Duodenal A. soehngenii L2-7 levels were below minimal detection rates, indicating that administered A. soehngenii L2-7 is not colonising the SI but rather transiting through the intestinal tract.
Supplemental material

Microbiota composition and faecal SCFA. (A) Principal coordinate analysis plot on the unweighted UniFrac distances showing the clusters of 16S rRNA sequences. (B) Alpha diversity (Shannon Index) in faecal microbiota composition in stool samples collected at baseline, 1 day, and 1 and 2 weeks after placebo/treatment intervention. (C) Faecal levels of butyrate (p=0.06), (D) acetate and (E) propionate shown as fold change of concentrations (nmol/mg dried faeces weight) obtained 1 day after intervention versus baseline. SCFA, short-chain fatty acid.
Given the capacity of A. soehngenii to produce butyrate (from sugars and lactate/acetate) and propionate (from 1,2-propanediol),10 18 19 SCFAs (butyrate, acetate and propionate) were measured in faecal samples taken at baseline and 24 hours after duodenal infusion. Surprisingly, glycerol placebo intervention significantly decreased the levels of butyrate and acetate (p=0.02 and p=0.01, respectively), whereas faecal SCFA remained stable on A. soehngenii L2-7 delivery (online supplemental figure S2A–C), indicating that the vehicle glycerol-containing solution inhibits SCFA production. However, when comparing the intervention-induced changes in SCFA concentrations within the first 24 hours, no significant differences were found between placebo and treatment groups (figure 2C–E). Nonetheless, fold-change values of butyrate tended to be higher following A. soehngenii L2-7 feeding (p=0.06, figure 2C).
Supplemental material
A. soehngenii L2-7 intake increases postprandial GLP-1 response and reduces the extent of glucose variability
To establish the immediate metabolic effects of A. soehngenii L2-7 intake, a standardised mixed meal test (MMT) was performed in all participants 6 hours postinterventions and the excursion of incretins, glucose, insulin and triglycerides was followed over 120 min. A significant increase in postprandial plasma GLP-1 levels was observed upon A. soehngenii L2-7 treatment (figure 3A,B; p=0.021). In line with this, during the first 24 hours, glucose excursions, determined as median absolute deviations (MADs) of continuous glucose measurements by FreeStyle Libre technology, were significantly diminished after A. soehngenii L2-7 intake compared with placebo infusion (figure 3C, p=0.045). In contrast, postprandial circulating levels of gastric inhibitory polypeptide (GIP), glucose, insulin and triglycerides were comparable between the two interventions (online supplemental figure S3A–D). Given the higher faecal butyrate levels 24 hours post-A. soehngenii infusion, we assessed the concentrations of plasma SCFA at the end of the MMT; however, no significant differences in butyrate, acetate or propionate levels were observed between placebo and treatment intervention-groups (online supplemental figure S4A–C).
Supplemental material
Supplemental material

GLP-1, glucose and BA metabolism. (A) Plasma GLP-1 levels (pmol/L) at 0, 20, 30 and 120 min during MMT. (B) Plasma GLP-1 levels during MMT as total AUC. (C) MAD of continuous glucose measurements over the first 24 hours after placebo/treatment intervention. (D) Total secondary BA plasma levels during MMT, shown as the sum of AUCs of TOMCA, TUDCA, TDCA, TUDCA, TLCA, GHDCA, GDCA, GUDCA, GlcA, omcA, DCA, UDCA, LCA, HDCA, MuroCA and IsoUDCA. (E) Correlation heatmap showing the Spearman’s r rank correlation coefficients and statistically significant correlations. *P≤0.05, **P≤0.01, ***P≤0.001 between BA and GLP-1 levels 6 hours postinfusion (6 hours) or during the MMT (AUC). MMT, mixed meal test; AUC, area under the curve; MAD, median absolute deviation; BA, bile acid; GLP-1, glucagon-like peptide 1; GDCA, glycodeoxycholic acid; TDCA, taurodeoxycholic acid; TLCA, taurolithocholic acid; isoUDCA; iso-ursodeoxycholic acid.
Since the genome of A. soehngenii L2-7 strain includes genes encoding a bile acid (BA) sodium symporter (EHLA_2286) and BA hydrolases (EHLA_1602 and EHLA_2245),20 21 we investigated the effects of A. soehngenii L2-7 on secondary BA. Although borderline significant (p=0.06), A. soehngenii L2-7 infusion augmented the postprandial excursions of secondary BA (figure 3D). Notably, the postprandial levels of the secondary BA taurodeoxycholic acid (TDCA), taurolithocholic acid (TLCA) and glycodeoxycholic acid (GDCA) positively correlated with the GLP-1 postprandial excursion rate, whereas iso-ursodeoxycholic acid (isoUDCA) rates were associated with GLP-1 concentrations at 6 hours postinfusion (figure 3E).
In light of the positive effects of A. soehngenii L2-7 on GLP-1 and BA plasma concentrations, we subsequently questioned whether the observed systemic outcomes were linked to differential expression of the butyrate and BA receptors in SI. Indeed, both butyrate and BA may act as GLP-1 secretagogues on intestinal L cells by binding the transmembrane receptors G protein-coupled receptor 43 (GPR43) and Takeda G protein-coupled receptor 5 (TGR5), respectively,22 23 whereas activation of the nuclear BA farnesoid X receptor (FXR) inhibits GLP-1 secretion.24 25 Gene expression of GPR43, TGR5, FXR and of the FXR target genes OSTa and FGF19 26 27 was analysed in duodenal biopsies taken at 6 hours postinfusion (figure 4A–E). Gene expression of GPR43, TGR5 and FGF19 was comparable between placebo and treatment (median fold change equal to 1.3, 1.1 and 0.9, respectively) (figure 4A,B,E). Instead, upon A. soehngenii L2-7 intake, the transcript levels of OSTa were significantly decreased, while FXR expression tended to be lower (median fold change equal to 0.8 and 0.86, respectively) (figure 4C,D), hinting to a reduced FXR activity.

Duodenal gene expression. Gene expression measured by quantitative in duodenal biopsies of (A) GPR43, (B) TGR5, (C) FXR, (D) OSTalpha and (E) FGF19. Data showing the relative gene expression (to placebo) using the 2–∆∆Ct method.
A. soehngenii L2-7 significantly impacts duodenal gene expression with a remarkable upregulation of regenerating islet-protein (REG)1B expression
To obtain an unbiased and in-depth snapshot of the intestinal transcriptome upon placebo/treatment infusions, we employed RNA sequencing (RNAseq) technology using RNA isolated from duodenum biopsies. The RNAseq data set analysis shows that a single dose of A. soehngenii L2-7 is sufficient to substantially change the transcriptomic profile of duodenal mucosa as early as 6 hours after intake. Indeed, using the digital gene expression (DGE) analysis pipelines Sleuth, EdgeR and DESeq2, we found respectively 380, 323 and 217 genes significantly upregulated or downregulated by A. soehngenii L2-7 intake (figure 5A). Only the genes with significant adjusted p values in all three statistical packages were retained, resulting in a total of 73 differentially expressed genes between placebo and treatment (figure 5B). Among these genes, REG1B, LCN2 and SLC6A14, showed an upregulation above two log2(fold-change) after A. soehngenii treatment, whereas DISP2 was the most downregulated gene (−1 log2(fold-change)) as compared with placebo (figure 5C). Overall, the most remarkable effect was the A. soehngenii-induced expression of REG1B, which encodes for the regenerating islet-derived 1 beta protein (figure 5C,D). Originally discovered in pancreatic calculi, Reg family members 1–4 are small secreted proteins that have been reported to promote proliferation, β-cell mass expansion and exert antidiabetogenic activities.28–30 We, therefore, further studied the gene and protein expression of Reg1B in SI biopsies after placebo/treatment interventions. By quantitative PCR (qPCR), we validated the RNAseq findings on REG1B expression and found that the expression of the closely related REG1A gene was also strongly upregulated in response to A. soehngenii treatment (figure 5E,F and online supplemental figure S5A), whereas the transcript levels of REG3A, REG3G and REG4 were not significantly upregulated by A. soehngenii L2-7 infusion (online supplemental figure S5B–D). We next questioned whether the treatment-mediated intestinal alterations are linked to systemic responses. Strikingly, an inverse correlation was found between the duodenal expression of the most upregulated genes REG1B, LCN2 and SLC6A14 and systemic glucose variability (MAD), monitored continuously in the first 24 hours (figure 5G). In addition, duodenal REG1B levels were positively associated with the expression rates of LCN2 and SLC6A14, and most importantly with the plasma concentrations of GLP1, underscoring systemic favourable effects of this single-bacterial strain intervention (figure 5G,H).
Supplemental material

RNAseq (duodenal gene expression). (A) RNAseq data sets analysed by three DGE technologies: Sleuth, EdgeR and DESeq2 were identified. Venn diagram showing the numbers of genes significantly upregulated or downregulated in either one of the pipelines. (B) Heatmap of the 73 top DE genes between placebo and treatment, identified by all three DGE technologies. (C) MA (ratio intensity) plot visualising the gene expression ratios (fold changes treatment vs placebo, Y axis) and the mean expression intensity (average RNAseq counts per gene, X axis) of DE genes. (D) RNAseq read counts of REG1B gene, duodenal expression. (E) Gene expression measured by qPCR in duodenal biopsies of REG1B and (F) REG1A, shown as fold change versus placebo by 2−ΔΔCt data analysis. (G) Correlation heatmap showing Spearman’s r rank correlation coefficients and statistically significant correlations. *P≤0.05, **P≤0.01, ***P≤0.001 between duodenal gene expression of REG1B, LCN2, SLC6A14 and median absolute deviation of continuous glucose measurements (MAD) over the first 24 hours after placebo/treatment intervention. (H) Spearman’s correlation between GLP-1 plasma concentrations 6 hours postinfusion and duodenal gene expression of REG1B (assessed by quantitative PCR). DE, differentially expressed; DGE, digital gene expression; GLP-1, glucagon-like peptide 1; RNAseq, RNA sequencing.
Subsequently, we investigated the intestinal expression and localisation of Reg1B and Reg1A proteins by western blotting and immunohistochemistry (IHC) in small intestinal tissues. At protein level, the mean expression of Reg1B and Reg1A within the duodenal mucosa trended toward higher expression after treatment, although not significant compared with placebo (figure 6A–C and online supplemental figure S5E–G). This inconsistency with gene expression rates is likely due to the fact that Reg proteins are secreted molecules. Immunostaining for Reg1B (performed with two different antibodies) showed a strong expression at the base of the crypts, where also stem cells and Paneth cells are located, and a milder staining throughout the villi and in the area delimiting vacuolated cells (figure 6D and online supplemental figure S5H). Comparing the IHC staining for Reg1B and Reg1A, at the same antibody concentrations, revealed that both proteins localize at the duodenal crypt bases, with Reg1B being more prominently expressed both in the crypt and villi compartments (online supplemental figure S5H). Since Reg1A and Reg1B belong to the same protein family and are highly similar proteins of almost equal size, we excluded cross-reactivity of anti-Reg1A/B antibodies by using recombinant human Reg1B and Reg1A proteins in western blotting assays, as performed by Zheng et al.31 The antibodies directed against Reg1B did not recognise Reg1A and vice versa (online supplemental figure S5I,K).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Reg1B expression in duodenum. (A) Western blot image of duodenal lysates blotted with antibodies against Reg1B and β-actin. (B) Quantification of Reg1B expression level in duodenal biopsies, Reg1B expression normalised to β-actin (loading control). (C) Reg1B protein expression shown as fold change treatment versus placebo. (D) Immunohistochemical staining of Reg1B in duodenal biopsies. (E) Sequential immunostaining of duodenal biopsies for Reg1B (red), lysozyme (yellow, Paneth cells) and mucins (deep blue, Goblet cells). Images shown at ×10 or ×40 magnification, as indicated. Arrow indicates one of the Reg1B+lysozyme colocalisation point. (F) Quantification of Reg1B expression level in Caco-2 cells, Reg1B expression normalised to β-actin (loading control); Reg1B expression shown as fold-change treatment versus placebo, but: butyrate 1 mM; MDP: 1 µg/mL. (G) Western blot images of Caco-2 cell lysates blotted with antibodies against Reg1B and β-actin. MDP, muramyl dipeptide.
To further unravel the nature of the cell types expressing REG1B within crypt-villus units, we performed a triple-staining to visualize Reg1B (red), lysozyme (yellow, Paneth cell marker), and mucins (Alcian blue dye, Goblet cell marker) (figure 6E). As in the single staining, the Reg1B red staining is prominently found at the crypt base, colocalised with the immunostaining (brown) of lysozyme, and it is adjacent to blue-dyed mucin-positive cells. Although further investigations are needed to understand the nature of these transcriptional changes, our findings indicate that Reg1B is expressed in intestinal villi, especially at the crypt-base, and might be produced and secreted by Paneth cells in response to A. soehngenii L2-7 transit. In this regard, the low concentrations of plasma Reg1B, assessed 8 hours postduodenal infusion (at the end of the MMT) (online supplemental figure S5J), suggest that Reg1B is mainly secreted into the (small) intestinal lumen, thereby acting in a paracrine manner on intestinal cells. To understand how A. soehngenii L2-7 may regulate Reg1B expression, we further exposed Caco-2 cells for 6 hours to either butyrate or muramyl dipeptide, the bioactive bacterial peptidoglycan motif, and found a marked upregulation of Reg1B by both stimulations (figure 6F,G). In line with this, direct exposure to heat-inactivated A. soehngenii L2-7 bacteria upregulates Reg1B expression (online supplemental figure S5L).
Importantly, no carry-over effects were observed between weeks 0 and 4, independently of the intervention order, as shown in online supplemental figure S6 for baseline faecal levels of A. soehngenii, faecal butyrate, plasma GLP1 excursions and glucose MAD in the first 24 hours postinfusion (online supplemental figure S6A–D).
Supplemental material
Discussion
In this pioneer randomised cross-over phase II trial, we demonstrate the early and wide impact of a single duodenal infusion of A. soehngenii L2-7 on the duodenal transcriptomic profile, and moreover, we identify the metabolic parameters being influenced (for up to 24 hours) by a single dose of live A. soehngenii L2-7 in treatment-naïve subjects with MetS. Indeed, administration of A. soehngenii L2-7 (vs placebo) resulted in an altered small intestinal gene expression signature and, most prominently, in the upregulation of REG1B. Moreover, A. soehngenii L2-7 infusion induced higher postprandial plasma bile salt and GLP-1 levels as well as lower glucose variability (MAD) within 24 hours after infusion of A. soehngenii L2-7. Although further investigations are warranted, these data, combined with our previous studies,13 14 suggest that A. soehngenii L2-7 improves human glucose metabolism in human MetS, likely by shaping BA metabolism and augmenting intestinal GLP-1 production.
We previously showed that, in subjects with metabolic syndrome, oral intake of A. soehngenii L2-7 for 4 weeks increased plasma primary and, particularly, secondary bile acids,14 which are known to be formed by commensal microbes, such as those belonging to the Ruminococcaceae and Lachnospiraceae.32 33 Moreover, in db/db mice, 4 week oral A. soehngenii treatment alleviated insulin resistance and modified bile salt metabolism in conjunction with augmenting the expression of genes involved in BA metabolism/transport in SI, including suppression of duodenal Fxr and Ost-alpha expression.13 Remarkably, we here disclose that a single duodenal infusion of A. soehngeniiis sufficient to increase secondary BA, of which TDCA, TLCA, GDCA, and isoUDCA associate with increase GLP-1 levels. These findings are in line with previous studies showing that conjugated BA, including TDCA, TLCA, GDCA and UDCA, promote GLP-1 release by intestinal L cells.34–38 Notably, the genome of A. soehngenii harbours a sodium symporter gene (EHLA_2286) as well as two bile salt hydrolase (BSH)-encoding genes (locus tags EHLA_1602 and EHLA_2245).20 Besides, transcriptome analysis of a simplified microbiota community (harbouring the A. soehngenii strain) engrafted in murine guts confirmed that both bsh genes are functionally expressed in the intestinal tract.21 These findings together with our observations of increased secondary BA on A. soehngenii feeding (this study,14) point to an active role of this strain in the formation of secondary BA, which may act as GLP-1 secretagogues by binding the TGR5 receptor.23 39 40 In addition, A. soehngenii decreased the duodenal expression of the FXR-target OSTalpha,26 consistent with a de-activation of FXR signalling observed in A. soehngenii-treated db/db mice.13 Notably, FXR suppresses GLP-1 secretion by enteroendocrine L cells, whereas its inhibition improves metabolic control.24 25 41 Thus, the diminished FXR activation may also account for more GLP-1 availability and, according to the study of Ducastel et al, more L cells’ responsiveness to butyrate/GPR43 signaling.25 Indeed, we observed a significant effect of A. soehngenii infusion on postprandial levels of GLP-1, but not GIP, suggesting that this strain sensitises L cells to secrete more GLP-1, likely in response to its own metabolites, such as butyrate and secondary (hydrophobic) BA. This hypothesis is supported by animal studies showing that gut microbiota has a rapid and pronounced effect on L cell and GLP-1 content, predominantly in the small intestine, where there is direct contact between enteroendocrine cells and mucosal microbiota.15 42 43 Moreover, A. soehngenii is a known producer of the SCFA butyrate,10 which by binding its receptor GPR43 expressed on L cells stimulates GLP-1 secretion.22 Overall, the A. soehngenii-mediated increase in plasma GLP-1, whether through butyrate/BA signalling and/or FXR inhibition, may justify the observed reduction in glucose MAD after bacteria intake. This improvement in glycaemic variability may be the result of the insulin sensitising effects of butyrate as well as GLP-1.11 ,16 ,17 In addition, we cannot exclude that A. soehngenii ameliorates the glycaemic control through alternative (less obvious) mechanisms: for example, by contributing to the generation of microbiota-derived neurotransmitters, in forms of proteins/peptides or gases, which can act locally on gut muscle relaxation or enteric neuron activation as well as distally on the brain influencing appetite, behavior and peripheral glucose homeostasis.44 Furthermore, a recent study disclosed that the beneficial action of Akkermansia muciniphila on the systemic metabolic profile are accounted by a bacterial protein, named P9, of the peptidase S41A family that stimulates, in vivo, thermogenesis and GLP-1 secretion.45 Curiously, A. soehngenii has the genetic capacity to express the peptidase S41 family, underscoring that multiple processes may dictate the observed metabolic benefits of A. soehngenii, beyond BA/butyrate production (eg, via production of bioactive molecules or possibly via Reg1B induction).
Although the fold-change comparison of faecal butyrate rates showed that butyrate tended to be higher after A. soehngenii-treatment, herewith, we also found a negative effect of placebo infusion on butyrate and acetate concentrations; an effect most likely due to side-effects of glycerol on SCFA production, as earlier described.46 Nonetheless, SCFA levels remained stable after A. soehngenii L2-7 administration suggesting that an increased SCFA production by A. soehngenii L2-7 counterbalances the reduction in SCFA caused by glycerol infusion and can hence better modulate intestinal GLP-1 production. In contrast to the changes seen in faecal butyrate, the plasma levels of butyrate were largely unaffected 8 hours after A. soehngenii-treatment. This might be due to the fact that microbially-produced butyrate is the primary energy source for colonocytes and hepatic lipid and glucose production1 rendering more difficult to detect disparities in its circulating peripheral plasma levels. Notwithstanding, we cannot exclude that variations in circulating SCFA concentrations become evident at a later time-point (eg, 24 hours, as for the faecal SCFA levels).
Being at the interface with intestinal microbiota, duodenal mucosa cells underwent a robust transcriptional reprogramming upon encountering A. soehngenii L2-7. The most differentially expressed genes after bacteria administration encode for proteins involved in metabolite transport, cholesterol metabolism or cytokine signalling. Nevertheless, the three most upregulated genes REG1B, LCN2 and SLC6A14 were found to negatively correlate with glucose MAD rates, hinting to a protective function in glycaemic control. In line, rodent studies demonstrate that increased LCN2 expression promotes glucose tolerance, insulin sensitivity and controls appetite47, whereas LCN2 deficiency worsens insulin resistance.48 Similarly, SLC6A14 deficiency in high-fat diet-fed mice worsens adiposity and MetS. Accordingly, the obesity-linked single-nucleotide polymorphism (SNP) in SLC6A14 has been shown to reduce SLC6A14 expression.49
The most prominent changes were seen in the expression of REG1B, which, along with REG1A, was markedly upregulated after A. soehngenii L2-7 administration. They are both members of the REG gene family, which was first discovered in pancreas, being expressed mainly by exocrine acinar cells and, upon cellular damage, in the islets of Langerhans.50 Animal studies using Reg knock-out, overexpression or administration of recombinant REG proteins showed that Reg proteins elicit mitogenic effects on β-cells and protection against diabetes.28 51 52 However, genomic SNPs in the REG1B were not associated with T2D53 suggesting that tissue specific expression is more relevant. Similarly to the pancreas, intestinal Reg protein expression has been linked to proliferation being enhanced in inflamed and neoplastic conditions.54–56 By immunostaining, we found that Reg1B and Reg1A are localised at the base of small-intestinal crypts with Reg1B being more prominently expressed by Paneth cells as compared with its expression (at a lower extent) in enterocytes; this is corroborated by a previous study describing REG1A, REG1B, and REG3 gene expression in Paneth cells.56 Notably, Paneth cells guard Lgr5+ stem cells in the crypt bases through production of signalling and bactericidal molecules. Indeed, Paneth cells have been reported to directly sense indigenous microbes via Toll-like receptor engagement and, thus, limit mucosa penetration by commensals by secreting antimicrobial products, which include Reg3β and Reg3γ proteins.57 In line with this, we disclose here that in enterocytes, Reg1B expression is triggered by peptidoglycan and butyrate. Although further investigations are needed, we can argue that intestinal epithelial and Paneth cells sense the administered bacteria via GPR and innate immune receptors, resulting in the induction of REG1 expression. The regenerating activities of Reg proteins on pancreatic cells seem to act in an auto/paracrine manner50 52; hence, it is likely that the duodenally secreted Reg1A/B acts locally, possibly inducing progenitor or L-cell hyperplasia. In support of a ‘local’ effect of Reg1B on intestinal L cells, duodenal REG1B levels were significantly associated with GLP-1 concentrations, and Reg1B concentrations were markedly lower in plasma samples than in duodenal tissues. Indeed, Reg1B was non-detectable in plasma samples of six patients 8 hours after placebo intake and in two samples following A. soehngenii L2-7 infusion. At protein levels, we did not find statistically significant differences in Reg1B or Reg1A expression in duodenum between placebo and treatment interventions. We therefore feel that this is likely due to their secretion into the intestinal lumen and hence loss of Reg proteins within the duodenal mucosa.58
As expected and importantly for the cross-over nature of our study design, the single duodenal bacteria infusion did not affect faecal microbiota composition and Shannon microbial diversity in a major manner, therefore excluding treatment-induced carry-over effects. Accordingly, the order of interventions did not impact the levels of faecal butyrate, postprandial GLP-1 responses nor the 24-hour glucose variability. Also, the lack of fluctuations in faecal A. soehngenii L2-7 abundancy over time was anticipated as it is unlikely that a single-dose of 1011 bacteria results in colonic colonisation. Notwithstanding, these findings implicate that the herein reported effects of A. soehngenii are triggered solely by its transit throughout the gut, and they would be greatly enhanced with a full bacteria engraftment of the SI/colon.
Limitations
Some limitations of this study need to be acknowledged. We administered the bacterial strain only once, as performed by van Baarlen et al 59 with a Lactobacillus strain; nonetheless, multiple infusions could permit bacteria colonisation of the gut and, consequently, elicit more prominent and lasting metabolic responses. Herein, by using a nosoduodenal infusion, we limited the deleterious effects of stomach acid and oxygen exposure on the viability of A. soehngenii L2-7, thus optimising the clinical potential of A. soehngenii. Nevertheless, future studies will have to demonstrate whether multiple bacterial administrations via duodenal-tube infusions or enteric-coated capsules will result in more pronounced effects.60
Conclusions
To our knowledge, this is the first study to administer a single strain of a strict anaerobe directly into the duodenum to maximally preserve viability bypassing the stomach. Single duodenal infusion of A. soehngenii L2-7 resulted in a significantly altered expression of small intestinal genes with the most prominent effect on REG1B, which was found to be associated with increased GLP-1 levels and improved peripheral glycaemic control and to be strongly expressed at the base of the intestinal crypts within Paneth cells. Moreover, the infusion of A. soehngenii L2-7 rapidly triggers favourable changes in metabolic parameters: it significantly enhances postprandial GLP-1 response (6 hours after intake) and ameliorates blood glucose variability (MAD, first 24 hours). Although A. soehngenii-derived bioactive metabolites and the incretin system may drive the improvement in glycaemic control and the insulin-sensitising effects of this strain, further studies are warranted to elucidate the mechanisms underlying the beneficial effects of A. soehngenii.
Materials and methods
Patient recruitment and involvement
Twelve Caucasian male subjects (age 21–69 years) with a body mass index between 30 and 43 kg/m2 were recruited by local newspaper advertisements (period of recruitment and follow-up: December 2017–February 2019). In order to be included in the trial, all subjects had to be treatment-naïve and suffer from MetS, determined by the presence of ≥3 criteria out of the five following criteria: fasting plasma glucose ≥5.6 mmol/L, triglycerides ≥1.7 mmol/L, waist circumference ≥102 cm, high-density lipoprotein cholesterol ≤1.04 mmol/L and blood pressure ≥130/85 mm Hg.61 Also, HOMA-IR (>2.5) was included as an extra screening marker of insulin resistance. Exclusion criteria included a history of cardiovascular event, cholecystectomy, overt untreated gastrointestinal disease or abnormal bowel habits, liver enzymes>2.5 fold higher than the upper limit of normal range, smoking, alcohol abuse and use of proton pump inhibitors or antibiotics in the past 3 months. Only men were included in the study to avoid confounding effects on insulin sensitivity due to changes in female hormone concentrations in (postmenopausal) women.62 Study participants were requested not to alter their physical exercise and dietary patterns after inclusion. The study was registered at the Dutch Trial Register.
Study design
This was a randomised double-blind placebo-controlled cross-over phase II study. All subjects (n=12) received both treatment (1011 A. soehngenii L2-7 cells, dosage based on our previous study14) and placebo (10% glycerol-phosphate-buffered saline, PBS), with a washout period of 4 weeks in between, as depicted in figure 1. The order of administration was randomised in a 1:1 fashion using computerised randomisation and was double-blinded (to patients and doctor in charge). After overnight fasting, a duodenal tube was placed using the electromagnetic-guided system Cortrak. The treatment arm received 10 mL of 10:90 glycerol:PBS solution containing A. soehngenii L2-7 (NCBI taxonomy ID 105843)9 at a concentration of 1010 cells/mL (total of 1011 cells) infused distally to the papilla of Vater, whereas the placebo arm underwent the same intervention, receiving only 10 mL of vehicle solution (10% glycerol in PBS). Six hours later, a gastroduodenoscopy was performed and duodenal biopsies were taken around the same location as the duodenal infusion and either stored in paraffin for histology or snap-frozen in liquid nitrogen and then stored at −80°C. After the gastroduodenoscopy, a 2-hour MMT was conducted as previously described.4 Subjects received an intravenous catheter in a distal arm vein over which baseline blood samples were drawn; hereafter, subjects immediately ingested a liquid meal solution (Nutridrink; Nutricia Advanced Medical Nutrition, Amsterdam, Netherlands) containing 600 kcal (35% fat, 49% carbohydrates and 16% proteins) and, for the subsequent 2 hours, blood samples were drawn for postprandial glucose, insulin, triglyceride, GIP and GLP-1 excursions (measured by standard clinical diagnostic methods). Subjects received a continuous glucose monitor (CGM, FreeStyle Libre System, Abbott USA) for 24 hours. Finally, subjects were asked to keep an online nutritional diary to monitor food intake during these days after the intervention (https://mijn.voedingscentrum.nl/nl/eetmeter/) and to collect faecal samples at several time-points (see figure 1). Four weeks after the first visit, the complete study cycle was repeated, switching intervention arms for each patient.
Data availability statement
All data relevant to the study are included in the article or uploaded as supplementary information.
Ethics statements
Patient consent for publication
Ethics approval
The study was approved by the local institutional review board of the Amsterdam University Medical Center in Amsterdam, the Netherlands, and conducted in accordance with the Declaration of Helsinki. Patients were not involved in the design and conduct of this research, although they were thoroughly informed about the procedures and goals of the study on recruitment visits. All participants signed a written informed consent.
Acknowledgments
We acknowledge Dr S Shetty and Dr H Heilig (Microbiology Lab, Wageningen University, The Netherlands), respectively, for helpful suggestions and DNA isolations from biopsies for Anaerobutyricum soehngenii L2-7 detection.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Footnotes
Contributors AK, AKG, ES, WMDV, MN and ER designed the study. AK, KW, JW, SM, HH, MW, SA, SH, HL, EMK, BH, JH, JJGHMB, MS, FB, ER, P-OB, JvS, MB, DMB, CMS and SLH performed the research. AK, ER, AP and EL performed the statistical analysis. AK, WMDV, AKG, MN and ER drafted the paper. All authors critically reviewed and approved the manuscript. ER acts as guarantor for this study and publication.
Funding MN is supported by a ZONMW VICI grant 2020 (number 09150182010020). WMDV is supported by the SIAM Gravitation Grant (024.002.002) and the 2008 Spinoza Award of The Netherlands Organization for Scientific Research. KW, MB, JvS, DMB and ER are appointed on a CAMIT grant 2018 (to MN and FB). The study reported here was additionally supported by Le Ducq consortium grant 17CVD01 to SLH and MN (on which HJH and CMS are appointed). MN and AGK are supported by a Dutch Heart Foundation CVON IN CONTROL-2 consortium grant.
Competing interests MN is in the scientific board of Kaleido Biosciences, Boston USA. WMDV is founder and in the board of A-mansia, Belgium. FB is in the scientific board of Metabogen AB, Sweden. MN and WMDV are founders and Scientific Advisory Board members of Caelus Pharmaceuticals, the Netherlands. SLH is a paid consultant for P&G and coinventor on pending and issued patents held by the Cleveland Clinic, and is eligible for receiving payments for inventions or discoveries related to cardiovascular diagnostics or therapeutics from Cleveland HeartLab, Quest Diagnostics and P&G.
Provenance and peer review Not commissioned; externally peer reviewed.
Supplemental material This content has been supplied by the author(s). It has not been vetted by BMJ Publishing Group Limited (BMJ) and may not have been peer-reviewed. Any opinions or recommendations discussed are solely those of the author(s) and are not endorsed by BMJ. BMJ disclaims all liability and responsibility arising from any reliance placed on the content. Where the content includes any translated material, BMJ does not warrant the accuracy and reliability of the translations (including but not limited to local regulations, clinical guidelines, terminology, drug names and drug dosages), and is not responsible for any error and/or omissions arising from translation and adaptation or otherwise.