Article Text

Abstract
Objective Micro-RNAs (miRNAs) play a crucial role in controlling intestinal epithelial barrier function partly by modulating the expression of tight junction (TJ) proteins. We have previously shown differential messenger RNA (mRNA) expression correlated with ultrastructural abnormalities of the epithelial barrier in patients with diarrhoea-predominant IBS (IBS-D). However, the participation of miRNAs in these differential mRNA-associated findings remains to be established. Our aims were (1) to identify miRNAs differentially expressed in the small bowel mucosa of patients with IBS-D and (2) to explore putative target genes specifically involved in epithelial barrier function that are controlled by specific dysregulated IBS-D miRNAs.
Design Healthy controls and patients meeting Rome III IBS-D criteria were studied. Intestinal tissue samples were analysed to identify potential candidates by: (a) miRNA-mRNA profiling; (b) miRNA-mRNA pairing analysis to assess the co-expression profile of miRNA-mRNA pairs; (c) pathway analysis and upstream regulator identification; (d) miRNA and target mRNA validation. Candidate miRNA-mRNA pairs were functionally assessed in intestinal epithelial cells.
Results IBS-D samples showed distinct miRNA and mRNA profiles compared with healthy controls. TJ signalling was associated with the IBS-D transcriptional profile. Further validation of selected genes showed consistent upregulation in 75% of genes involved in epithelial barrier function. Bioinformatic analysis of putative miRNA binding sites identified hsa-miR-125b-5p and hsa-miR-16 as regulating expression of the TJ genes CGN (cingulin) and CLDN2 (claudin-2), respectively. Consistently, protein expression of CGN and CLDN2 was upregulated in IBS-D, while the respective targeting miRNAs were downregulated. In addition, bowel dysfunction, perceived stress and depression and number of mast cells correlated with the expression of hsa-miR-125b-5p and hsa-miR-16 and their respective target proteins.
Conclusions Modulation of the intestinal epithelial barrier function in IBS-D involves both transcriptional and post-transcriptional mechanisms. These molecular mechanisms include miRNAs as master regulators in controlling the expression of TJ proteins and are associated with major clinical symptoms.
- INTESTINAL BARRIER FUNCTION
- GENE EXPRESSION
- IRRITABLE BOWEL SYNDROME
- MOLECULAR BIOLOGY
- RNA EXPRESSION
This is an Open Access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/
Statistics from Altmetric.com
Significance of this study
What is already known on this subject?
Differential messenger RNA (mRNA) signatures correlate with ultrastructural abnormalities in the intestinal epithelial barrier in patients with diarrhoea-predominant IBS (IBS-D).
A distinctive micro-RNA (miRNA) expression profile has been identified both in the intestinal mucosa and in the peripheral blood of patients with IBS-D.
However, combined high-throughput analysis of the expression of miRNA and mRNA profiles in the jejunal mucosa has not been performed.
What are the new findings?
We provide evidence of miRNA-dependent modulation of tight junction-specific proteins by using an integrative approach combining both mRNA expression data from RNA sequencing analysis and miRNA expression profiles in the jejunum of patients with IBS-D.
Expression of hsa-miR-125b-5p and hsa-miR-16 is downregulated while their respective target proteins, cingulin and claudin-2 are upregulated.
Functional analysis of the role of hsa-miR-125b-5p and hsa-miR-16 identified these miRNAs as modulators of barrier function.
These molecular alterations correlated with bowel dysfunction, perceived stress and depression and with number of mucosal mast cells.
How might it impact on clinical practice in the foreseeable future?
Defining the complex array of interactions of miRNAs with apical junctional proteins will pave the way for potential diagnostic and therapeutic interventions that would reinforce the intestinal barrier function, consequently preventing or ameliorating inflammatory reactions.
Introduction
IBS has traditionally been considered as a functional GI disorder defined by clinical manifestations lacking specific and sensitive biological markers. However, recent cumulative evidence highlights the plausible organic origin of IBS symptoms. Particularly, distorted mucosal barrier ultrastructure,1–4 immune activation and low-grade inflammation5–9 have been implicated in IBS. Latest studies have shown differential expression of genes related to alterations in immune system signalling pathways and intestinal barrier function.10–12 In addition, our group has lately described differential mucosal humoral activation and transcriptional signatures correlated with ultrastructural abnormalities in the epithelial barrier associated with mast cell activation and major clinical manifestations in patients with diarrhoea-predominant IBS (IBS-D).1 ,13 ,14
Previous studies have mainly focused on the expression of protein-coding genes and their correlation to clinical outcomes. As a matter of fact, <3% of the human genome encode proteins,15 consequently, the majority of the transcribed human genome represents non-coding RNAs.16 ,17 Among them, micro-RNAs (miRNAs) are particularly relevant for intestinal inflammatory disorders as recent research reported on their impact in the regulation of immune and inflammatory responses.18 ,19 To date, their role in IBS has been addressed only in a few studies. The first evidence for a miRNA-regulated expression of serotonin (5-HT) receptor genes in IBS20 ,21 was reported as association of IBS-D with gene variants residing in regulatory regions causing disturbed regulation by hsa-miR-510 and hsa-miR-16 family. The respective variants seem to impair miRNA binding to the target region, thereby reducing translational repression and increasing protein expression. More recent studies have reported distinctive miRNA profiles in IBS-D analysing the intestinal mucosa22 and peripheral blood.23 The role of miRNA-driven expression regulation in the gut is underlined by a study investigating the relevance of miRNAs during development, differentiation and function of the intestinal epithelium in a knockout mouse of the pre-miRNA processing enzyme Dicer1.24 Of note, Dicer1-deficient animals showed significant changes in miRNA profiles correlating with a remarkable disorganisation of the gut epithelium, impairment of the intestinal barrier and intestinal inflammation.24
Therefore, we hypothesised that miRNA-driven dysregulation of intestinal immune system activation and epithelial barrier function is involved in the pathophysiology of IBS-D. Our specific aims were to identify miRNAs differentially expressed in the small bowel of patients with IBS-D compared with healthy controls (HC) and to identify target proteins specifically involved in intestinal epithelial barrier function. We applied the following multistage strategy: (1) identification of differentially expressed miRNAs and their target transcripts in patients with IBS-D versus HC by RNA sequencing (RNAseq) and miRNA profiling; (2) ascertainment of regulatory networks, biological functions and upstream regulators associated with the potential targets identified in the previous approach by pathway analysis and (3) investigation of the functional relevance of specific miRNA's modulation of endogenous protein expression and epithelial barrier function in intestinal epithelial cell lines.
Methods
Participants
Newly diagnosed patients meeting Rome III IBS-D criteria25 and HC were prospectively recruited from the gastroenterology outpatient clinic (see also online supplementary methods section). HC were recruited from the general population by public advertising. Prior to entering the study, a complete medical history and physical examination were carried out in both patients and controls. All subjects completed structured clinical questionnaires (to characterise digestive symptoms in patients and to verify the lack of symptoms in HC) and underwent allergy tests to rule out food allergy (see below). Healthy subjects reporting abdominal symptoms were excluded from the study. In addition, past episodes of infectious gastroenteritis and GI comorbidities were reasonably excluded by performing a broad biochemical and serologic profile including antitransglutaminase antibodies, upper and lower fibre optic and small bowel capsule endoscopy, abdominal sonography and barium studies, when considered pertinent. The study protocol was approved by the Ethics Committee at the Hospital Vall d'Hebron (PR(AG)159/2011). Written informed consent was obtained from each participant.
supplementary data
Using daily questionnaires over a 10-day period, the following parameters were recorded: (a) severity of abdominal pain by a 100-point visual analogue scale; (b) frequency of abdominal pain (number of days with pain); (c) stool frequency (day with maximum number of bowel movements) and (d) stool consistency assessed by the Bristol stool form score.26 Background stress and depression levels were evaluated using the validated Spanish versions of the Modified Social Readjustment Scale of Holmes-Rahe,27 by the Perceived Stress Scale of Cohen28 and by the Beck's Inventory for Depression.29
Skin prick tests (SPT) were performed with 22 common food allergens (Laboratorios Leti, Barcelona, Spain), using histamine and saline as positive and negative controls, respectively. Positivity was defined by skin weal reaction (diameter >3 mm) to at least one allergen. Candidates with either positivity to foodstuff by SPT or clinical history consistent with food allergy (digestive and/or extradigestive symptoms associated with exposure to certain food components) were excluded.
Experimental design and procedures
To investigate the role of differentially expressed miRNAs and messenger RNAs (mRNAs) in the aetiopathology of IBS, total RNA isolated from gut biopsies of IBS-D and HC was subjected to next-generation sequencing. Tissue samples were obtained as follows: a single mucosal biopsy per participant was obtained from the proximal jejunum, 5–10 cm distal to the Treitz's angle, using a Watson's capsule as described previously.5 Tissue samples were immediately split into two similar pieces with a sterile scalpel. One fragment was fixed in formalin and embedded in paraffin for further microscopic examination assessing inflammation by routine procedures and mast cell numbers by CD117 staining. The remaining fragment was placed in RNAse-free tubes containing 500 μL of RNA Later Solution (Life Technologies) and stored at −80°C until processed for RNA and protein isolation.
Eighty-five subjects were initially recruited. Five samples from the healthy control group were excluded due to abnormal results (>40 intraepithelial lymphocytes) on routine histology. After RNA isolation, 11 subjects were also excluded due to insufficient RNA quality (RNA integrity number (RIN) <5). Therefore, a total of 69 subjects (43 patients with IBS-D and 26 HC) were finally included in the study. The following analyses were subsequently performed in blindly selected subgroups of subjects (see online supplementary figure S1 and table S1). The selection of subjects for the mRNA-miRNA discovery cohort was made only among those samples showing a RIN value >8. The selection of samples for the other analyses was then made based on RNA/protein availability. Detailed description of procedures is given in the online supplementary methods section.
mRNA and miRNA profiling
Total RNA was isolated from jejunal biopsies of a discovery cohort comprising eight patients with IBS-D and eight HC. Then, isolated RNA was analysed as follows:
mRNA expression was assessed by RNAseq on an Illumina Hi-Seq 2000.
miRNA expression was assessed by two different technologies: nCounter analysis system (Human_v2_miRNA expression assay kit, from miRBase v14.0) and Agilent microarrays (Agilent's Unrestricted_Human_miRNA_V16.0).
Pathway analysis
To identify relevant biological pathways implicating those genes differentially expressed, we applied ingenuity pathway analysis (IPA) methodology (IPA Software, Ingenuity Systems, http://www.ingenuity.com). IPA integrates selected omics data sets (genomics, transcriptomics, miRNAomics, proteomics) with mining techniques to predict functional connections and their interpretation in the context of protein networks that comprise protein-protein interactions and related biological functions and canonical signalling pathways. Two types of analysis were performed in our profiling data (see online supplementary figure S2):
mRNA and miRNA core analysis to identify the biological functions of the differentially expressed miRNA-target genes followed by identification of putative upstream regulators.
miRNA target filter plus miRNA-mRNA pairing analysis in order to identify coregulated miRNA-target mRNA pairs.
Validation of profiling results
Quantitative PCR (qPCR) was used to validate miRNA profiling results and to assess expression of additional miRNAs identified in subsequent pathway analysis in samples from 14 IBS-D versus 18 HC.
A customised nCounter Gene Expression CodeSet was used in order to validate results obtained by RNAseq in additional jejunal RNA samples from 31 IBS-D versus 17 HC. This CodeSet included oligonucleotide probes for detection of genes involved in epithelial barrier function (see online supplementary table S2).
Further validation of results at the protein level was performed by western blot analysis in 25 patients with IBS-D versus 15 HC. Antibodies used can be found in online supplementary table S3.
In addition, molecular changes were correlated with major clinical and histopathological features of patients.
Functional analysis
The functional interaction between specific miRNA candidates and their respective putative target genes was assessed by overexpression and inhibition of candidate miRNAs in a human intestinal epithelial cell line, colo320. Candidate miRNAs and their respective mutants harbouring disrupted seed sequences were cloned into pEP-miR expression vectors and transfected into colo320 cells. Detailed information on the cloning strategy can be found in online supplementary table S4. Endogenous levels of putative target proteins were measured by In-Cell Western (ICW) analysis (see online supplementary table S3).
The impact of candidate miRNAs on epithelial barrier function was assessed using the T84 cell line as a well-established model for human intestinal epithelia cells. This human colon cell line is able to polarise and establish fully functional tight junctions (TJs) in vitro when seeded on transwell inserts.30 Due to the low efficiency and short-term effect of the transient transfection of plasmids, we used a lentivirus-mediated expression system and subsequently selected T84 cells, where hsa-miR-125b-5p and hsa-miR-16 are either stably overexpressed or downregulated by respective miRNA-sponges (molecules loaded with a multitude of respective complementary miRNA binding sites, acting as competitive inhibitors of miRNAs).31 The created stable T84 cells were then seeded on collagen-coated transwells and the permeability of the epithelial layer was assessed by measuring transepithelial electrical resistance (TEER) every day for 7 days. Polarisation and formation of TJs was also controlled by immunostaining of the established TJ protein marker zonula occludens 1 (ZO1). After overexpression or downregulation of the candidate miRNAs, endogenous levels of putative target proteins were measured by western blot analysis. Additionally, the structure of TJs was also assessed by counting the number of nuclei surrounded by the belt of ZO1.
Statistical analysis
Statistical analysis of miRNA profiling data
miRNA expression values were normalised to take into account possible batch effects and intersample variability. The median absolute deviation (MAD) of expression values over all samples was calculated for each miRNA. miRNAs showing the 5% lowest MADs, and miRNAs with missing values for some samples were excluded from further analyses. Wilcoxon two-sided rank tests were applied to identify miRNAs with a differential expression in the two groups of samples. miRNAs with corresponding p values <0.05 were followed-up (see online supplementary material). Biostatistical analyses were conducted using R V.2.15.2 (R Development Core Team).
Statistical analysis for mRNA profiling data
RNAseq reads were aligned to the reference genome of Homo sapiens version GRCh37 (ENSEMBL based) obtained from iGenomes (https://ccb.jhu.edu/software/tophat/igenomes.html). No prior filtering of sequencing data was applied since anomalous reads were automatically discarded by the alignment programme. Total reads per sample ranged from 18 to 43.5 million. Only mRNAs with mean normalised read counts above 100 were investigated. The differential expression analysis was then performed using the Bioconductor package DESeq2.32 Normalisation was performed using the package defaults by computing sample-specific size factors to control for batch effects and other biases present in the data. Empirical distribution functions for the normalised counts showed no systematic differences across samples. Gene level count tables were obtained using the count script of the HTSeq python library. Raw p values from a negative binomial distribution were calculated by DESeq2 and they were used as input to fdrtool in order to compute q-values (false discovery rates (FDRs)). Genes with a FDR <0.1 were considered differentially expressed (see online supplementary material).
Statistical analysis for qPCR, western blot, ICW and immunofluorescence data
Two-tailed parametric or non-parametric tests were used as appropriate (unpaired Student's t-test, Mann-Whitney U test, two-way analysis of variance followed by Bonferroni post-tests) using GraphPad Prism 5.0 software. Relationships between clinical variables and gene expression were assessed by Spearman's correlation rho. Data are summarised by mean±SD, unless otherwise stated. Adjustment for multiplicity was carried out using objective-specific methods. FDRs were calculated to identify the list of genes/miRNAs used as input for pathway analyses using the Benjamini and Hochberg method.33 Bonferroni-adjusted probability values were calculated to account for multiple comparisons in one-miRNA experiments.
Results
Study population
We found no differences in age, gender proportion or body mass index between patients and controls (table 1). Patients showed significantly higher frequency of dyspepsia and scores of perceived stress and depression than HC (table 1). No differences in disease severity between discovery and validation cohorts was found (p=0.42). Furthermore, subgroups did not differ in clinical characteristics of applied biological assessments (p>0.23 for all) (see online supplementary table S5).
Clinical and demographic characteristics of participants
Specific genes are differentially expressed in the jejunal mucosa of patients with IBS-D
On average, ∼29 million reads were obtained for each sample and, the average mapping rate to the human genome was ∼80%. A total of ∼10 million reads were excluded for further analysis due to different reasons (not showing unique alignments, alignment was ambiguous or not overlapping any gene features) leading to ∼14 million reads that were used as the basis for counting. The counts were then normalised and filtered rendering ∼15 000 mRNAs. Differential expression analysis was then performed by DESeq2, which identified 3806 mRNAs differentially expressed (mean normalised read counts >100; FDR <0.05) in IBS-D versus HC. IPA analysis identified a number of canonical signalling pathways key for intestinal homeostasis as playing an important role in IBS-D (table 2).
Canonical signalling pathways associated with IBS-D gene expression profile
As a proof of principle, and to validate earlier findings of impaired barrier function, genes related to the TJ signalling pathway were selected and subsequently analysed by the nCounter technology in a larger patient cohort (IBS-D, n=37; HC, n=17). Similar results were obtained by both techniques (table 3). In addition, no differences in the expression of any of these genes were found comparing patients with and without functional dyspepsia (see online supplementary table S6).
nCounter validation of RNAseq data
The jejunal mucosa of patients with IBS-D shows a distinctive miRNA expression signature
Data from miRNA profiling was used for network analysis by IPA. The most significant network identified was composed of eight miRNAs differentially expressed in IBS-D samples compared with HC (FDR <0.05) linked to their respective target proteins (figure 1). In this network, hsa-miR-125b-5p was identified playing a central role by targeting proteins involved in pathways related to the epithelial barrier function like apoptosis of epithelial cells, TJ and actin-cytoskeleton signalling pathways (figure 1). These functions were also highlighted by RNAseq data (table 2).

Relationships between differentially expressed micro-RNAs (miRNAs) and their putative targets and related canonical pathways and biological functions (Fx). The list of differentially expressed miRNAs in IBS-D, compared with healthy controls, linked to their approved nomenclature (http://www.mirbase.org) and fold-change was uploaded into the ingenuity pathway analysis (IPA) application. Target filter analysis was performed by IPA to get target genes and interactions. Node (target gene/miRNA) and edge (relationship) symbols are described in the figure. The intensity of the node colour indicates the degree of upregulation (red) or down regulation (green). Genes in uncoloured nodes were not identified as differentially expressed in our study and were integrated into the computationally generated networks on the basis of the evidence stored in the IPA knowledge base indicating relevance for this network.
miRNA-mRNA pairing analysis identified intestinal epithelial-related functions and humoral immune response as potentially dysregulated in the jejunal mucosa of patients with IBS-D
In order to narrow down the list of relevant distinctive molecular targets playing a role in IBS-D, target filter analysis and miRNA-mRNA pairing was performed combining both differentially expressed mRNAs and miRNAs. This analysis identified 1393 mRNAs that were experimentally observed or predicted with a moderate/high score to be targets of differentially expressed miRNAs in IBS-D. The top molecular functions associated with the target genes included apoptosis of epithelial cells and several functions related to the architecture and disorganisation of apical junctional complexes and the humoral immune response (table 4). These results confirm our previous findings from different sample cohorts.1 ,13 ,14
Molecular functions associated with the target genes after miRNA-mRNA pairing analysis
Moreover, to further validate the biological relevance of our results, IPA was used to identify particular miRNAs that may explain the observed mRNA differential expression in patients with IBS-D, so called by IPA as upstream regulators. In this analysis, hsa-miR-125b-5p and hsa-miR-16-5p were identified as the most significantly deregulated miRNAs predicted to be inhibited based on the expression pattern of their targets (table 5). Downregulation of both miRNAs in IBS-D samples was then validated by qPCR (figure 2A).
Upstream regulators associated with mRNA targets

Expression analysis of candidate micro-RNAs (miRNAs) and their potential target proteins in IBS-D versus healthy controls (HC) jejunal samples. (A) Quantitative PCR validation of differentially expressed miRNAs in patients with IBS-D. To obtain the fold-change value for each sample the ratio between target miRNA and the average of reference genes was calculated for each sample and then normalised to the average of the healthy group. Groups were compared using the non-parametric Mann-Whitney U test. **p<0.01. (B) miRNA binding sites in the 3′-untranslated region (3′-UTR) of the barrier function-related genes cingulin (CGN) and claudin-2 (CLDN2) identified by TargetScan. (C) Claudin-2 and cingulin protein expression in the jejunal mucosa. Protein expression was measured by western blot in patients with IBS-D and healthy control subjects. Protein fold-change was calculated for each sample with reference to the average of the target protein to GAPDH ratio of the healthy control group. Comparisons were performed by the Mann-Whitney U test (p values shown).
CGN and CLDN2 are targets of hsa-miR-125b-5p and hsa-miR-16
Based on the previous results, we decided to further follow-up hsa-miR-125b-5p and hsa-miR-16 and identify which candidate mRNAs involved in epithelial barrier function were being targeted by these two miRNAs. Therefore, we performed bioinformatics analysis of potential miRNA binding sites by miRWalk34 (http://www.umm.uni-heidelberg.de/apps/zmf/mirwalk/index.html) and identified two putative miRNA binding sites in the 3′-untranslated region (3′-UTR) of the TJ protein encoding genes CGN (cingulin) and CLDN2 (claudin-2) for hsa-miR-125b and hsa-miR-16, respectively (figure 2B, see online supplementary tables S7 and S8). As a first step towards validating the putative miRNA-based regulation of CGN and CLDN2, we analysed their protein expression levels in the jejunal mucosa of patients with IBS-D compared with HC by western blot analysis. In concordance with downregulation of hsa-miR-125b-5p and hsa-miR-16 (figure 2A), CGN and CLDN2 protein levels were found to be upregulated in IBS-D (figure 2C). In addition, no differences in the expression of either hsa-miR-125b-5p and hsa-miR-16 or their target proteins, CGN and CLDN2 were found comparing patients with and without functional dyspepsia (see online supplementary table S6).
We next investigated the putative regulation of CGN and CLDN2 in vitro after overexpression and subsequent inhibition of hsa-miR-125b-5p and hsa-miR-16. For that purpose, we selected an established human epithelial cell line, colo320, which shows endogenous expression of CGN and CLDN2 on protein level (figure 3A). In order to confirm miRNA-target site interaction, the candidate hsa-miR-125b-5p or hsa-miR-16 were transfected into colo320 cells. In addition, both miRNAs were mutated in order to impair their binding ability to the target site (figure 3B). Endogenous levels of CGN and CLDN2 were subsequently quantified and compared with that of mock-treated cells by ICW analysis. Protein levels of both putative target genes was reduced by respective candidate miRNAs and rescued in case of particular miRNA mutants (figure 3C). To rule out unspecific events, we assessed changes in CGN and CLDN2 expression in relation to hsa-miR-510, a miRNA not predicted to target these proteins. No significant effect on protein levels was observed for hsa-miR-510, confirming no interaction with the putative targets (figure 3C).

hsa-miR-125b-5p and hsa-miR-16 target cingulin and claudin-2, respectively. (A) Endogenous claudin-2 and cingulin protein expression in epithelial cell lines. Protein expression was measured by western blot in colo320 and HEK293 cells; a jejunum sample was used as a positive control. (B) Wild-type and mutant sequences of the pEP-miR vectors used. (C) In-Cell Western (ICW) was applied to measure endogenous protein expression after cotransfecting colo320 cells with either wild-type or mutant pEP-miR vectors and the pEP-miR null vector as a control miR. A pEGFP-C1 construct was cotransfected as a transfection and normalisation control. Quantification of endogenous proteins was measured and analysed according to the Odyssey Infrared Imaging System of LI-COR. (D) ICW was applied to measure endogenous protein expression after cotransfecting colo320 cells with either wild-type or mutant pEP-miR expressing vectors and specific mirVANA inhibitors (50 nM for miR-125b inhibitor and 150 nM for miR-16 inhibitor) or negative controls. pEP-miR-510 was used as a control miRNA. A pEGFP-C1 construct was cotransfected as a transfection and normalisation control. Quantification of endogenous proteins was measured and analysed according to the Odyssey Infrared Imaging System of LI-COR. Graphs represent results from six independent experiments. Data are expressed as mean±SD. Comparisons were performed by two-way analysis of variance followed by Bonferroni post-tests. *p<0.05; **p<0.01; ***p<0.001. ctrl, control.
To further validate miRNA-target gene interactions, colo320 cells were analysed with candidate miRNAs and their mutant forms (figure 3B) in addition to specific miRNA inhibitors or a negative inhibitor control. Endogenous protein levels of both, CGN and CLDN2, were reduced in cells cotransfected with the negative inhibitor control, while the mutant miRNAs blocked this effect (figure 3D). On the other hand, endogenous expression of CGN and CLDN2 were upregulated in case of specific miRNA inhibitors (figure 3D).
hsa-miR-125b-5p and hsa-miR-16 are involved in the impairment of TJ structure and epithelial barrier function by modulating CGN and CLDN2 expression
The functional effect of candidate miRNAs on cellular permeability was assessed using the T84 cell line as a model for human intestinal epithelial cells. First, miRNA-driven modulation of CGN and CLDN2 was confirmed in T84 cells. Stable overexpression of hsa-miR-125b-5p and hsa-miR-16 led to increased expression for both miRNAs, while inhibition by miRNA sponges led to downregulation of endogenous hsa-miR-125b-5p and hsa-miR-16 expression (figure 4A). Endogenous levels of both target proteins, CGN and CLDN2, were reduced in T84 cells overexpressing the respective candidate miRNA (figure 4B). In concordance, cells transfected with the specific sponges showed increased levels of CGN and CLDN2 (figure 4B). No changes in ZO1 expression were detected (figure 4B).

Epithelial barrier function is impaired by hsa-miR-16 and hsa-miR-125b-5p in T84 cells. T84 cells were created that stably expressed hsa-miR-16 or hsa-miR-125b-5p overexpression or downregulation constructs (sponges). (A) Stable cells lines were evaluated for miRNA expression by qPCR. Values are normalised to control cells. Graphs represent results from three independent experiments. Data are expressed as mean±SD. Comparisons were performed by the Mann-Whitney U test versus control (Ctrl) miR cells. *p<0.05. (B) Stable cell lines were evaluated for cingulin (CGN), claudin-2 (CLDN2) and zonula occludens 1 (ZO1) expression by western blot. Actin was used as a loading control. Representative image of a triplicate experiment is shown. (C and D) Stable cell lines were seeded onto transwell inserts and epithelial barrier function was monitored over time by measuring transepithelial electrical resistance after overexpression (C) or downregulation (D). Graphs represent results from three independent experiments. Data are expressed as mean±SD. Comparisons were performed by two-way analysis of variance followed by Bonferroni post-tests. ***p<0.001.
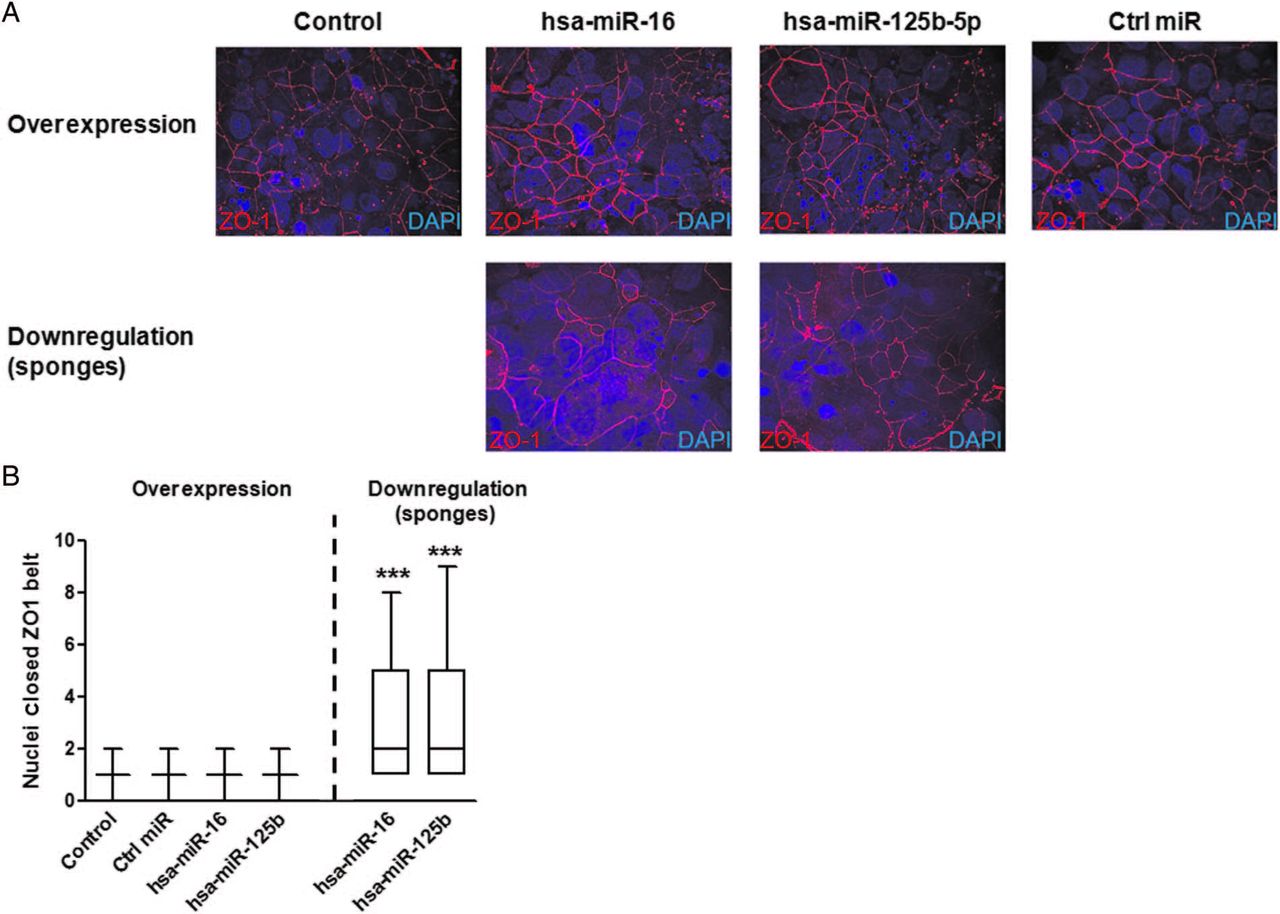
Finally, in order to assess whether modulation of CGN and CLDN2 by these two miRNAs had an influence on epithelial permeability, TEER was measured in T84 cells every day during 7 days. Downregulation of the respective miRNAs led to increased permeability in each case (decreased TEER) in T84 cells, while particular miRNA overexpression caused the opposite effect (figure 4C). In addition, influence on TJ structure was assessed by analysing ZO1 distribution in polarised T84 cells (7 days postseeding). While overexpression of hsa-miR-125b-5p and hsa-miR-16 did not affect the characteristically continuous belt-like pattern of ZO1 staining around the apical membrane of T84 cells, inhibition induced a highly disorganised ZO1 staining pattern for each miRNA (figure 5A). Disruption of TJ structure was confirmed by increased number of cell nuclei contained in complete ZO1 belts after inhibition of both miRNAs indicating a decrease in ZO1 belt structures/impaired barrier (figure 5B).

Tight junctions structure is impaired by hsa-miR-16 and hsa-miR-125b-5p. T84 cells were created that stably expressed hsa-miR-16 or hsa-miR-125b-5p overexpression or downregulation constructs. Cells were seeded onto transwell inserts and allowed to polarise for 7 days. (A) 7 days postseeding cells were fixed and stained for the tight junction protein zonula occludens 1 (ZO1) (red) and cell nuclei were stained for 4′,6-diamidino-2-phenylindole (DAPI) (blue). (B). Nuclei per complete ZO1 were quantified. Graphs represent results from three independent experiments. Fifty fields per sample were counted. Comparisons were performed by the Mann-Whitney U test versus control cells. ***p<0.001.
Major clinical and histopathological features of patients with IBS-D correlate with hsa-miR-125b-5p and hsa-miR-16 and their target genes, CGN and CLDN2
To assess the potential clinical relevance of our findings, we applied the Spearman's correlation rho to pooled data of patients and HC. Both miRNAs studied negatively correlated with bowel habits (see online supplementary table S9), while hsa-miR-125b-5p also correlated with the frequency of abdominal pain (see online supplementary table S9). Conversely, CGN and CLDN2 protein expression positively correlated with bowel habits. On the other hand, chronic stress levels measured by the Holmes-Rahe scale did not correlate with the expression levels of these two miRNAs or their target proteins (see online supplementary table S10). Yet, we found a negative correlation between the expression of miRNAs and perceived stress and depression, measured by the Cohen and the Becks scales, respectively (see online supplementary table S10).
Moreover, patients with IBS-D showed increased mast cell numbers (IBS-D: 26.46±12.68; HC: 17.80±9.52 CD117+ cells/hpf; p=0.0007). Downregulation of hsa-miR-125b-5p and hsa-miR-16 negatively correlated with mast cell numbers (figure 6A); while CGN and CLDN2 protein expression positively correlated with numbers of mast cells (figure 6B).

Statistical correlations. (A) Correlation between mast cell numbers and claudin-2 and cingulin protein expression in the jejunal mucosa of IBS-D and healthy controls. Spearman's correlation rho was calculated in the pooled dataset of healthy controls (n=15) and patients with IBS-D (n=23). (B) Correlation between mast cell numbers and hsa-miR-16 and hsa-miR-125b-5p expression in the jejunal mucosa of IBS-D and healthy controls. Spearman's correlation rho was calculated in the pooled dataset of healthy controls (n=18) and patients with IBS-D (n=14). CD117+/hpf, number of mast cells per high power field, mast cells were measured by staining with CD117.
Discussion
In this study, we provide evidence that the modulation of the intestinal epithelial barrier function in IBS-D involves both transcriptional and post-transcriptional mechanisms, including miRNAs hsa-miR-125b-5p and hsa-miR-16 as master regulators in controlling the expression of specific TJ proteins. Moreover, expression of both miRNAs and their target proteins correlate with major symptoms and mast cell hyperplasia, supporting the involvement of these cells in the impairment of intestinal epithelial barrier function as a central molecular mechanism in this disease. The data gained in this study, together with previous reports of other groups and our own research line contribute to further delineate the current view of IBS-D origin in which the organic basis of this disorder is substantiated by a heterogeneous network of immunological responses, involving hyperactivation of mast cells, T and B lymphocytes and impaired epithelial apical junctional structure and its association with intestinal barrier dysfunction and cardinal symptoms (figure 7).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Model summarising alterations in different cellular, molecular and structural components playing a crucial role in disturbing intestinal homeostasis in IBS-D. New findings in this study are highlighted with a red square. Transmission electron microscopy images are from our own picture database. Cartoons used in this figure were taken from Servier Medical Art (http://www.servier.com/Powerpoint-image-bank).
The essential role of miRNAs in the modulation of protein expression in the intestinal epithelium has been firmly established.24 ,35–37 In line with this, altered miRNA profiles have been reported in intestinal diseases including IBS.22 ,23 ,38 ,39 Now, we show additional evidence on deregulation of miRNA expression in the jejunum of patients with IBS-D. In fact, recently a comprehensive study assessed miRNA profiles in different organs of the human body and revealed different expression profiles of miRNAs in small versus large intestine.40 Of note, none of the previously reported miRNAs22 ,38 ,39 were confirmed in our study, probably due to gut region-specific differences. This represents the major drawback of comparisons of data gained in different studies on various regions of the gut owing to distinct functions of the proximal and distal gut. Consequently, differential expression of the same individual molecules (ie, genes, proteins or miRNAs) should not be necessarily expected along the whole intestine. This issue is well reflected by the discrepancies observed between different studies in IBS.6 ,41 Nonetheless, it is remarkable that molecular changes observed in earlier studies hit similar pathways leading to impaired epithelial barrier function and increased intestinal permeability. Another factor contributing to discrepancies between published data is represented by the application of different methodology and data analysis pipelines. Currently, no gold standard for miRNA profiling exists. Due to their short sequences (22–25 nucleotides) and high degree of homology between miRNA family members, miRNA detection is technically challenging. A recent comparative study on established miRNA profiling platforms revealed variation in their reproducibility, sensitivity and accuracy.42 To overcome these limitations, we combined the profiling data from two of the best performing miRNA platforms42 and validated our findings applying a third method (qPCR). In addition, this is the first study on IBS integrating miRNA and mRNA profiling data. This paired analysis led to identification of a set of miRNA targets differentially expressed in the jejunum of patients with IBS-D. Notably, 65% of the pathways associated with the RNAseq data obtained in this analysis were also found in our previous microarray study,13 including those related to the modulation of epithelial barrier function through TJs and caveolar-mediated endocytosis. The discrepancy between datasets may be attributed to the different methods used: microarrays which are restricted to detection of specific isoforms versus RNA sequencing that detects the whole transcriptome and more accurately fetches the repertoire of gene isoforms. The data obtained here, therefore, reinforces and extends our results using a different profiling technique on a different set of patients.1 ,13 ,14 We decided to focus on candidates related to the modulation of apical junctional complexes (ie, TJ, caveolar-mediated endocytosis and actin cytoskeleton signalling) to keep in line with our major research hypothesis and because other studies reporting on miRNA regulation in IBS also showed differentially regulated candidates involved in the epithelial barrier function.22 ,38 In addition, other functions and pathways were identified in our analysis with higher significance, like epithelial cell death and apoptosis categories which, in turn, may also relate to dysfunctional epithelial barrier associated with deregulation of miRNA activity in the gut.24 Candidates belonging to these functions and how they are modulated by miRNAs in IBS-D, indeed, merit further assessment in the future.
The two most promising miRNA candidates, hsa-miR-125b-5p and hsa-miR-16, were chosen based on IPA analysis of miRNA and mRNA profiles. The 3′-UTR of genes from the TJ signalling pathway were systematically scanned in order to find binding sites for these two miRNAs. Results of the most comprehensive miRNA search tool miRWalk showed two putative binding sites in CGN for hsa-miR-125b-5p and in CLDN2 for hsa-miR-16 in four and five out of five prediction tools, respectively. CGN localises in the cytoplasmic surface of TJs of intestinal epithelial cells43 and interacts with the actomyosin cytoskeleton and ZO proteins.44 Cingulin does not seem to be required for the basic structure and canonical function of TJ.45–47 However, it contributes to modulating gene expression of TJ proteins during epithelial differentiation45 through a yet not completely understood mechanism involving RHOA activity46 and GATA4 expression.48 In addition, CGN is a predicted target for HNF4A, a transcription factor regarded as key regulator of intestinal differentiation.49 Therefore, the activity of CGN in modulating TJ dynamics must be viewed in the context of a wider signalling network that is likely to be differentially modulated under different physiological and pathological conditions. CGN protein overexpression and, consistently, hsa-miR-125b-5p downregulation was confirmed in IBS-D samples. Fine-tuning the activity of CGN and its functional network by miRNAs and its implications for intestinal barrier dysfunction related to intestinal inflammation and, particularly, IBS-D remains to be investigated. Remarkably, the claudin family is responsible for modulating passage through the paracellular route and alterations in expression and distribution have been associated with several intestinal diseases.50 Recent reports have shown the implication of claudin deregulation in epithelial barrier function in IBS.1 ,38 ,51 ,52 Our own previous data showed increased claudin-2 protein levels in the jejunum of patients with IBS-D as part of the molecular mechanism that may account for disrupted TJ ultrastructure and increased permeability.1 Now we confirmed the upregulation of claudin-2 in a different set of patients with IBS-D and validate it as a real target for hsa-miR-16 which, consistently, is downregulated in IBS-D samples. In addition, we used a cell model to mimic the situation described in patients with IBS-D (downregulation of hsa-miR-16/hsa-miR-125b-5p and upregulation of CLDN2/CGN) showing the functional disruption of the epithelial barrier as a consequence and, therefore, providing evidence on the impact of miRNA modulation of specific TJ proteins in the increased intestinal permeability that has been consistently described in patients with IBS-D in earlier studies.53
Interestingly, mast cell numbers correlated positively with CGN and CLDN2 protein expression and negatively with their respective targeting miRNAs. On the other hand, miRNA alterations correlated negatively with bowel habits and with the acute stress response and depression levels. The negative correlations found in our study indicate that those patients showing less expression levels of the respective miRNA and consequently, increased target protein expression and disturbed barrier, are suffering from increased mast cell counts and higher levels of stress and depression. We and others have previously reported that stress can deeply affect permeability via mast cell activation.54 ,55 Remarkably, several miRNAs, including hsa-miR-125b-5p, have been found to be highly expressed in exosomes from mast cell lines.56 As exosomes are well known to participate in cell-to-cell communication delivering their contents to target cells, it could be speculated that mast cell-mediated modulation of epithelial permeability could be driven by direct delivery of high amounts of hsa-miR-125b-5p (and other miRNAs) to epithelial cells in homeostatic conditions. Disruption of homeostasis would then trigger downregulation of hsa-miR-125b expression. Studies aimed at deciphering this interesting connection would be key to further determine the role of miRNAs in mucosal inflammation and barrier disruption. However, whether miRNA deregulation, mast cell hyperactivity and impaired barrier function are the cause or the consequence will still remain to be determined. In addition, evidence collected to date has shown that miRNA expression is altered in patients suffering from depression and anxiety and in animal models of early life, restraint and unpredictable chronic mild stress.57–60 Of note, hsa-miR-16 has been reported to regulate other important genes of the serotonergic system, for example, the serotonin reuptake transporter gene SLC6A4, a key regulator of serotonin availability in the nervous system and gut epithelium.58 In the central nervous system, hsa-miR-16 had been reported to mediate depression behaviours through regulation of SLC6A458; furthermore, a mechanistic connection to depression through neurogenesis pathways was observed61 and hsa-miR-16 has recently been linked to anxiety.62 Taken together, our results suggest that miRNA-driven modulation of TJ proteins may cause epithelial barrier dysfunction in the jejunum of patients with IBS-D that could explain, at least in part, the bowel dysfunction characteristic of this disease. However, caution is advised when interpreting the functional consequences of these data as correlations obtained are only modest and they do not imply causation and other mechanisms can be playing a role to explain our observations. In addition, data involving miRNAs in the modulation of mast cell biology are still scarce; therefore, the precise mechanism linking mast cells, miRNA deregulation and development of symptoms needs further exploration.
We would like to acknowledge that this study was conceived as an exploratory study and, therefore, no prior power calculation was performed, yet sample size was in the same range of similar reports12 ,22 ,38 ,39. In addition, although we isolated RNA, miRNA and protein from the same sample, we could not get enough material to perform all different analyses in all participants. Therefore, the number of samples for some of the experiments was relatively small. However, we followed a very exhaustive and stringent phenotyping protocol for both patients and controls which provided us with a highly homogeneous cohort. In addition, a major strength of our study is that for functional validation of results we analysed endogenous levels of the target proteins instead of using artificial reporter systems that could bias the results.
In summary, we show specific miRNA regulation of TJ proteins associated with major clinical symptoms in IBS-D. This study together with previous results, come to reinforce our line of research where organicity is evidenced in IBS-D, mainly affecting gut epithelial barrier integrity. Defining the complex array of interactions and modulations of miRNAs with apical junctional proteins will pave the way for potential diagnostic and therapeutic interventions that would reinforce the intestinal epithelial barrier function, potentially restabilising intestinal homeostasis.
Acknowledgments
We would like to thank all patients and healthy volunteers for their kind support in participating in this study and the supporting staff at each site. Thanks to Milagros Gallart, Montse Casellas and Carmen Alastrue for expert technical assistance; Anna Aparici, Maria Teresa Casaus and Purificación Rodríguez for their invaluable assistance in the performance of jejunal biopsies. Special thanks to Dr Vladimir Benes and the staff from the EMBL Gene Core Facility in Heidelberg for their excellent technical and scientific support in sample processing and RNA sequencing analysis. We also thank Dr Ramesh Pillai for fruitful discussions throughout the project and Dr Cristina Frias for assistance in RNA isolation.
References
Footnotes
CM, BKR-J, MV, BN and JS contributed equally.
Contributors CM designed the project, performed the research and wrote the paper; BKR-J isolated RNA and protein from all samples and was involved in all research procedures; BL, CA-C, MP, FA and JS were in charge of recruiting patients/controls and collected the biopsies; MLS and SB created T84 stable cell lines and performed overexpression/inhibition analysis and measurements of TEER; BK, MG, WH, RGS and JL were in charge of all statistical analysis regarding RNA sequencing and micro-RNA profiling; RR performed nCounter experiments; IdT was in charge of routinely screening biopsies to exclude signs of inflammation and stained and counted mast cells; GR contributed to essential tools and reagents; BN, MV and JS contributed to the design of the project, supervised all stages of the research and wrote the paper. AMG-s AND ES-R collaborated in the processing of tissue samples and data collection. All authors revised and approved the final version of the manuscript.
Funding Supported in part by Fondo de Investigación Sanitaria and CIBERehd, Instituto de Salud Carlos III, Subdirección General de Investigación Sanitaria, Ministerio de Economía y Competitividad: CM08/00229 (BL); CM10/00155 (MP); EII2011-0035 and CD15/00010 (BKR-J); PI12/00314 (CA-C); CP10/00502 and PI13/00935 (MV); PI08/0940, PI11/00716 and PI14/00994 (JS); Ministerio de Educación, Dirección General de Investigación: SAF 2009-07416 (FA); Agència de Gestió d'Ajuts Universitaris i de Recerca, de la Generalitat de Catalunya: 2009 SGR 219 (FA), 2011-BP/A00099 and 2011-BP-A2/00002 (CM); The Rome Foundation Award 2013 (MV); Centro de Investigación Biomédica en Red de Enfermedades Hepáticas y Digestivas: CB06/04/0021 (FA); Chica and Heinz Schaller Foundation and Deutsche Forschungsgemeinschaft (DFG) in SFB1129 (Project 14) (SB); Brigitte-Schlieben Lange Program from the state of Baden Württemberg, Germany and the Dual Career Support from CellNetworks, Heidelberg, Germany (MS) and the University Hospital Heidelberg (BN, GR). This manuscript results in part from collaboration and network activities promoted under the frame of the international network Genes in IBS Research Network Europe, which has been funded by the COST programme (BM1106, http://www.GENIEUR.eu) and is currently supported by the European Society of Neurogastroenterology and Motility (http://www.ESNM.eu).
Competing interests None declared.
Ethics approval Vall d'Hebron Ethics Committee.
Provenance and peer review Not commissioned; externally peer reviewed.