Article Text

Abstract
Objective To determine if human colonic neuromuscular functions decline with increasing age.
Design Looking for non-specific changes in neuromuscular function, a standard burst of electrical field stimulation (EFS) was used to evoke neuronally mediated (cholinergic/nitrergic) contractions/relaxations in ex vivomuscle strips of human ascending and descending colon, aged 35–91 years (macroscopically normal tissue; 239 patients undergoing cancer resection). Then, to understand mechanisms of change, numbers and phenotype of myenteric neurons (30 306 neurons stained with different markers), densities of intramuscular nerve fibres (51 patients in total) and pathways involved in functional changes were systematically investigated (by immunohistochemistry and use of pharmacological tools) in elderly (≥70 years) and adult (35–60 years) groups.
Results With increasing age, EFS was more likely to evoke muscle relaxation in ascending colon instead of contraction (linear regression: n=109, slope 0.49%±0.21%/year, 95% CI), generally uninfluenced by comorbidity or use of medications. Similar changes were absent in descending colon. In the elderly, overall numbers of myenteric and neuronal nitric oxide synthase-immunoreactive neurons and intramuscular nerve densities were unchanged in ascending and descending colon, compared with adults. In elderly ascending, not descending, colon numbers of cell bodies exhibiting choline acetyltransferase immunoreactivity increased compared with adults (5.0±0.6 vs 2.4±0.3 neurons/mm myenteric plexus, p=0.04). Cholinergically mediated contractions were smaller in elderly ascending colon compared with adults (2.1±0.4 and 4.1±1.1 g-tension/g-tissue during EFS; n=25/14; p=0.04); there were no changes in nitrergic function or in ability of the muscle to contract/relax. Similar changes were absent in descending colon.
Conclusion In ascending not descending colon, ageing impairs cholinergic function.
- human
- colon
- ageing
- axon transport
- choline acetyltransferase
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
Significance of this study
What is already known on this subject?
Studies in ageing rodents suggest loss of myenteric neurons and a decline in cholinergic function of the colon. However, not all studies agree and uncertainty exists in terms of how these findings translate to humans.
Studies using human colon have generally used small numbers of tissues with variable or undefined clinical history and heterogeneous techniques, generating inconsistent data.
Nevertheless, a decline in neuromuscular functions has been suggested to contribute to the mechanisms underlying the increased incidence of constipation among the elderly.
What are the new findings?
In a large functional study (ascending/descending colon from, respectively, 109 and 130 patients), increasing age increased the likelihood that electrical nerve stimulation caused muscle relaxation in ascending colon, instead of contraction, but no age-related changes were observed in descending colon.
The functional changes with increasing age were not explained by loss of neurons within the myenteric plexus, reduced density of intramuscular axon bundles or changes in numbers of nitrergic neurons, but were associated with an increase in choline acetyltransferase (ChAT) immuno-positive myenteric nerve cell bodies in ascending, not descending colon.
Functional changes with increasing age were explained by a decline in cholinergic function in ascending but not in descending colon (allowing an unchanged nitrergic function to exert greater inhibitory influence on cholinergic activity).
There were no regional or age-related differences in the ability of the smooth muscle to contract or relax.
How might it impact on clinical practice in the foreseeable future?
The new findings indicate that absolute or selective neuronal loss is not likely in itself to be an important aetiological factor in lower GI symptoms associated with ageing.
The detection of age-dependent changes in ascending but not descending colon (decreased cholinergic function and increased myenteric somal ChAT immunostaining) indicates that greater attention needs to be paid to region-dependent differences in how the human colon ages and in particular, the changes which occur within the cholinergic innervation.
Introduction
There is evidence that human GI functions change in the elderly (>65–70 years),1 contributing to the development of constipation with associated pain, loss of dignity, reduced quality of life, faecal impaction and incontinence.1 2 Approximately 30%–40% of people aged 65 years or older self-report constipation.3 However, other factors are important, including changes in diet, fluid intake, exercise, bowel disorders (eg, diverticulitis or bowel cancer) and use of medications that reduce GI motility (eg, opioid-based analgesics, tricyclic antidepressants, calcium-channel antagonists2 4).
In rodents, declining colonic neuromuscular functions are reported during increasing age; some5 6 (not all7) report reduced numbers of myenteric cholinergic neurons and glial cells, reduced mucosal secretory capacity and slower intestinal transit. However, caution is required when translating such data to humans. Rodents have high metabolic rates, relatively short life spans8 and GI physiology distinct from primates, with functional disparities reflected by differences in anatomy, neuronal functions, receptor pharmacology and molecular structures.9 Further complications are caused by genetic variation between different strains of laboratory rodents,10 including differences in loss of myenteric nitrergic neurons during advanced age.11 Alternative use of baboons12 does not entirely remove translational uncertainty (the colon has adapted to fruit and vegetable diets13) and regardless, these animals are not suitable for routine research. Functional studies in humans have generated contradictory findings, perhaps due to small sample sizes with unclear medical history and heterogeneous experimental approaches.14–16
This study aimed to determine if age-associated changes in neuromuscular functions can be identified using ex vivo preparations of human ascending and descending colon and then determine mechanisms of change by multiple, systematic anatomical and pharmacological approaches.
Materials and methods
Patients and tissues
After ethical approval (REC 10/H0703/71), written informed consent was obtained for use of macroscopically normal ascending and descending/sigmoid (referred to hereon as descending) colon (5–10 cm from tumour) from patients undergoing elective surgery for non-obstructing bowel cancer. Patient records were examined for ongoing medication and comorbidity; clinical investigations into bowel functions were not conducted. No patient had previous chemoradiotherapy or diagnosis of inflammatory bowel disease. Tissue was immersed into preoxygenated Krebs solution 60–120 min after surgery. When investigating mechanisms, two discontinuous age groups were studied, either side of the overall median age of patients examined: adult (35–60 years) and elderly (≥70 years).
Anatomical studies
Histopathology
Full thickness tissues, orientated to demonstrate mucosal, submucosal, muscularis and serosal layers, ~1×1 cm, were fixed and embedded in paraffin blocks prior to cutting into multiple 4 µm thick sections. Sections were stained with H&E for routine histopathological assessment. Significant inflammation was excluded by observation of crypt architecture, distortion and atrophy, lamina propria cellularity, basal plasmacytosis, active inflammation, granulomata, mucin depletion, crypt abscesses, cryptitis and ulceration.17
Myenteric neuronal quantitation
Neurons were immunostained using anti-human neuronal protein C/D (anti-HuC/D; pan-neuronal antigen for cytoplasmic RNA-binding proteins, labelling cell nucleus and perikaryon18), anti-choline acetyltransferase (anti-ChAT) and anti-neuronal nitric oxide synthase (anti-nNOS). For each, a median of seven sections were examined for each patient (so not less than 10 mm of stained myenteric plexus could be studied, usually more). Each section was cut from the same block (minimum 16 µm separation). Following antigen retrieval and visualisation, cell nuclei were counterstained with hematoxylin. Positive and negative controls were performed in colon (anti-Hu/C/D, anti-nNOS) and spinal cord (anti-ChAT), with or without primary antibodies. Stained slides were scanned and digitally visualised.
For neuronal quantitation, a line was drawn along the myenteric plexus (freehand draw function in viewing software). For ganglion counts, sections stained with anti-HuC/D were viewed at ~40× magnification. To quantify cell bodies within each ganglion, sections stained with anti-HuC/D were viewed at 60×–80× magnification. Subclasses of myenteric cell bodies were counted separately after staining with anti-ChAT and anti-nNOS. Counting of ganglia and cell bodies19 and other assessments of staining were performed by two independent observers ‘blinded’ to patient identity/age (online supplementary file 1).
Supplementary file 1
Densitometric analysis of intramuscular nerve fibre bundles
Sections were labelled using anti-protein gene product 9.5 (anti-PGP9.5) (online supplementary file 1) to examine changes in axons/nerve fibre bundles in the muscle. Stained slides were scanned, digitally visualised and two non-adjacent areas from each of three different regions investigated per section: circular muscle near myenteric plexus, circular muscle close to the mucosa (deep circular muscle) and longitudinal muscle.
Cholinergic enzyme assays
ChAT, acetylcholinesterase (AChE) and butyrylcholinesterase (BChE) expression and activity in colon muscle were measured as previously described (online supplementary file 2).
Supplementary file 2
Functional studies
Detailed methods have been described previously.20 21 In brief, after removing the mucosa, strips were cut parallel to the circular muscle (~5 mm wide, 10–15 mm long; 3–31 from each patient) and mounted in warmed tissue baths containing Krebs solution (mmol/L: NaCl 121.5, CaCl2 2.5, KH2PO4 1.2, KCl 4.7, MgSO4 1.2, NaHCO3 25, glucose 5.6, equilibrated with 5% CO2/95% O2) for measuring isometric muscle tension. After 1-hour recovery, electrical field stimulation (EFS) was applied and consistent responses obtained.
Responses to EFS were analysed qualitatively by counting numbers of tissues relaxing or contracting. Quantitative measurements were of changes in tension/gram (g) wet weight of tissue; contractions were assigned positive values and relaxations negative values (if relaxation followed contraction, assignment was to the movement of greatest magnitude). Drug-induced changes were differences between the mean of three responses before and 30 min after drug application. In determining the effects of L-NAME (Nω-nitro-L-arginine methyl ester hydrochloride) and atropine (inhibiting, respectively, nitrergic and cholinergic functions in human colon20), one strip/patient was selected from multiple strips/patient exposed to these drugs for a number of different studies (computer random number generation). Similarly, when determining the influence of age on these responses, strips were randomly selected to match the proportions of phenotype (contraction/relaxation in response to EFS) observed in each region of colon. Non-cumulative concentration– response curves for carbachol (muscarinic M1–M5 receptor agonist) and sodium nitroprusside (SNP; nitric oxide (NO) donor) were established by applying single concentrations for 15 min or until maximum effect was observed (online supplementary file 3 shows carbachol concentration–response curves).
Supplementary file 3
Drugs were freshly prepared prior to use. Carbachol, atropine, L-NAME , SNP and DEA-NO (diethylamine NONOate; diethylammonium (Z)−1-(N,N-diethylamino)diazen-1-ium-1,2-diolate; Sigma, UK), MRS2500 (P2Y1 receptor antagonist in human colon22) and tetrodotoxin (TTX; Tocris, UK) were dissolved in distilled water. The L-type calcium channel stabiliser (S)-(-)-Bay K8644 was dissolved in dimethyl sulfoxide (DMSO) (Sigma) to 10 mM. A cocktail of neurokinin (NK)1, NK2 and NK3 receptor antagonists was prepared by dissolving L732138 (Tocris) in DMSO, GR159897 (Tocris) and SB235375 (GSK) in ethanol, each to 10 mM, for use at, respectively, 1 µM, 0.1 µM and 0.1 µM bath concentrations, inhibiting tachykinergic functions in human colon.20
Statistical analysis
To test whether patient characteristics affected responses to EFS, measured initially when surveying the effects of age, contingency analyses (χ2 or Fisher’s exact test) were applied as a screening test to two groups (higher or lower than the mean percentage strip relaxation of adult ascending colon), with a conservative level of statistical significance used (p≤0.01) to allow for multiple comparisons. On the basis that only one of the 130 covariates evaluated appeared to confound results, multivariable analyses were not performed.
In mechanism-based functional experiments, EC50 and Emax values were obtained from three-parameter agonist–response curves using GraphPad Prism 7.02. Data are expressed as means±SEM. The n values represent number of patients. Differences between means were determined using analyses of variance (ANOVAs) with Sidak’s multiple comparisons test for unpaired observations. P value <0.05 represented statistical significance.
For anatomical studies, numbers of cell bodies were expressed per millimetre of myenteric plexus. Inter-rater differences (95% CI) were analysed using one-sample Student’s t-tests and proportional biases investigated using Bland-Altman plots (online supplementary file 1). For comparisons, normality testing (D’Agostino & Pearson) was performed and neurons/mm length analysed for differences in means using ANOVAs with Sidak’s multiple comparisons test (11/12 variables were normally distributed). P value<0.05 represented statistical significance.
Results
Patients
Two hundred and forty-five patients (122 women; mean age 66 (range 35–91) years) consented to provide fresh surgical colon (table 1). On H&E staining, only a minority of adult and elderly patients showed low-grade inflammation limited to the mucosa.
Patients and tissues used
Age-related changes in overall function
Before examining mechanisms of change we looked for existence of any non-specific change in neuromuscular function, using a standard burst of EFS to evoke neuronally mediated contractions/relaxations in muscle strips from large numbers of human ascending and descending colon over a wide age range (35–91 years).
Preliminary experiments
To determine the optimal frequency to study, frequency–response curves were constructed using adult (n=34) and elderly (n=58) tissues (females and males assessed together). In adult ascending/descending colon, 1–20 Hz EFS usually caused contraction often followed by after-contraction on termination of EFS (figure 1A). TTX 1 μM prevented responses to 1–5 Hz EFS, but small monophasic contractions remained at higher frequencies (figure 1B). L-NAME 300 µM prevented any relaxations and increased the amplitude of contractions during EFS. Atropine 1 µM prevented contractions during EFS, revealing small muscle relaxations, particularly in ascending colon (figure 1B). Compared with adults, ascending colon from the elderly exhibited little-or-no muscle movement during 1–2 Hz EFS and contractions during 5–20 Hz appeared smaller (respectively n=19, 32; there were no age-dependent differences in after-contractions), whereas in descending colon from the elderly, any effects of increasing age were unclear (n=12–26; online supplementary file 4).
Supplementary file 4
These small numbers of patients precluded meaningful statistical analyses. However, they supported initiation of a larger investigation in which EFS was applied at the single frequency of 5 Hz, evoking a robust, TTX (1 µM)-sensitive muscle contraction or relaxation, usually followed by a large after-contraction (figure 1D). It was confirmed that atropine 1 µM and L-NAME 300 µM prevented, respectively, contractions and relaxations during EFS, enhancing the opposing response (figure 1D, table 2). Atropine also reduced the amplitude of any after-contractions, reduced further by NK1,2,3 receptor antagonism; MRS2500 1 µM had no consistent effects (table 2).

The effects of electrical field stimulation (EFS) in circular muscle of ascending and descending colon from adult (35 to 60 years) and elderly (≥70 years) patients. Panel A shows representative trace examples illustrating responses during and after termination of EFS over a range of frequencies of stimulation. EFS was applied at 1–20 Hz and at 50 V for 10 s every 1 min. Using tissues from the adult group, panel B shows the overall contraction force (mean±SEM g tension/g wet weight of tissue) generated during EFS in both regions of colon for each frequency of stimulation in the presence of vehicle, tetrodotoxin (TTX) 1 µM, atropine 1 µM or L-NAME (Nω-nitro-L-arginine methyl ester hydrochloride) 300 µM (n=5 each). Panels C and D show responses to EFS at 5 Hz, 50 V for 10 s, repeated every 1 min. The individual trace in panel C illustrates the ability of TTX 1 µM to inhibit responses evoked by repeated EFS. In panel D, the effects of single bursts of EFS are shown before and after treatment with atropine 1 µM or L-NAME 300 µM, in two muscle strips cut from the same tissue (male, 74, descending colon). The 10 s period of EFS is indicated by the horizontal bars (note the expanded time scale relative to panel C). Atropine inhibited contractions during EFS and decreased after-contractions. L-NAME inhibited relaxations during EFS and facilitated contraction amplitudes.
Pharmacology of responses to EFS in circular muscle from human adult ascending and descending colon
Age-dependent changes in neuromuscular responses
During EFS at 5 Hz, the occurrence of contractions or relaxations sometimes varied among different muscle strips from the same colon (figure 1D). To investigate the effects of advancing age, a mean of 10 strips (3–31 from each patient, depending on tissue availability) were examined (239 patients). The numbers of strips relaxing or contracting during EFS, and contracting after EFS, were expressed as the percentage total for that patient (note: the number of strips used from each region of colon was similar and showed no age-dependent selection bias; data not shown).
Advancing age (35 to 91 years) increased the likelihood that muscle relaxation, rather than contraction, occurred during EFS in ascending colon (figure 2A,C). In descending colon, there were no age-related changes in responses evoked during EFS (figure 2B,C) and the incidence of after-contractions was unchanged in either region of colon (figure 2C).
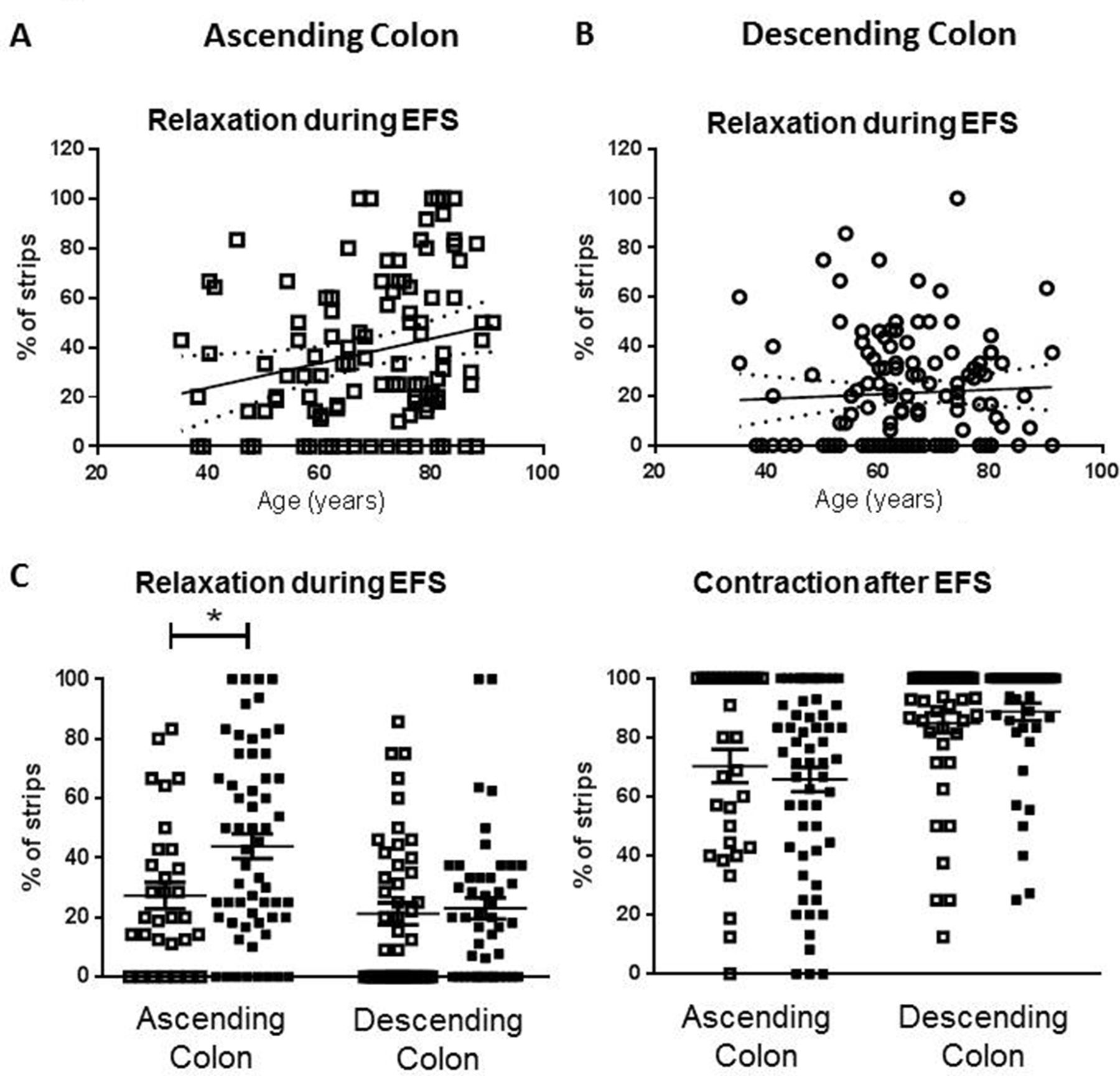
Age-dependent changes in phenotype of response to electrical field stimulation (EFS) in human ascending but not descending colon. For each patient (n=239), a mean of 10 strips (range 3–31) were examined and the number which relaxed or contracted during EFS expressed as the percentage of total tested for that patient. Panel A shows the percent values for relaxation during EFS in ascending colon from all patients. The linear regression line (95% confidence bands as dotted lines) demonstrates an increased likelihood of relaxation as age increases (slope=0.49%±0.21%/ year, n=109). Panel B shows no age-related changes in the percentage values for relaxation during EFS in descending colon (all patients: slope=0.09%±0.17%/year, n=130). Panel C shows the overall percentage of strips relaxing during EFS for each of the tissues studied and the percentage that contract after EFS, placed in the adult (35–60 years old; □) and elderly (≥70 years old; ■) age groups. For ascending colon, n=31 and 58 and for descending colon, n=45 and 46. The data are given as means (±SEM). These were analysed using analysis of variance with Sidak’s multiple comparison tests; *p<0.05. For comparison between adult ascending and descending colon contraction after EFS, p=0.06, all other comparisons between adult groups, and adult and elderly groups were not statistically significant (p>0.70).
Multivariate logistic regression showed these results were largely uninfluenced by clinical variables with known potential to affect colon functions (table 3, online supplementary file 5) although use of α-adrenoceptor antagonists appeared associated with reduced occurrence of muscle relaxation during EFS.
Supplementary file 5
Contingency analyses to identify effect of patient state and trait variables on responses evoked by 5 Hz EFS in the ascending colon (n=109)
Subsequent studies to examine mechanisms of change were conducted using two discontinuous age groups (35–60 and ≥70 years), either side of the median age.
Mechanisms: anatomical
Myenteric neuronal cell bodies staining for nNOS and ChAT
In 36 patients (8 adult, 9 elderly ascending; 9/10 adult/elderly descending), a mean of 36 mm of myenteric plexus/antibody/ patient was sampled and 30 306 myenteric neurons counted (646 sections). 18 838 neurons were stained for HuC/D (3520/3104 adult/elderly ascending, 6580/5634 descending), ChAT (5047 neurons, respectively, 827/1153 and 1711/1356) and nNOS (6421 neurons, respectively, 1627/1235 and 1743/1816). Reported changes were not influenced by interobserver differences (online supplementary file 1).
Figure 3A shows examples of labelled sections; table 4 provides a summary. In adults, there appeared to be fewer myenteric ganglia and a smaller number of nerve cell bodies (corresponding to a smaller number of ChAT-staining cells) in ascending, relative to descending colon (respectively, n=8, 9 patients); these numbers did not change with increasing age (figure 3B; respectively, n=9, 10). Compared with other antibodies, the signal-to-background differentiation for anti-ChAT was less strong. Nevertheless, in the elderly, the numbers of ChAT-immunolabelled neurons were consistently and significantly greater in ascending colon (figure 3C) but unchanged in descending colon. There were no statistically significant age-related differences in numbers of neurons expressing nNOS for either region (figure 3C). Post hoc calculation of statistical power (Stata SE v14.0) for the observed difference in ChAT immunostaining between adult and elderly ascending colon was 0.97 (expressed alternatively, only five patients would be required in each group to detect the observed difference with a power of 0.8). For the observed difference in HuC/D staining between ascending and descending colon, the post hoc power was 0.82.

Expression of myenteric neuronal markers in ascending and descending colon. Panel A shows representative staining examples from the adult (35–60 years of age) and elderly group (≥70 years) for each region of colon, using antibodies for human neuronal protein C/D (HuC/D), choline acetyltransferase (ChAT) and neuronal nitric oxide synthase (nNOS). Images were captured using NDP View version 2.3.1. Black scale bar is 250 µm. Counting was performed by two independent observers, and the values for each tissue are the mean of these counts. A ganglion was defined as a neural structure containing at least two neurons. The areas counted as neurons were between the circular and longitudinal muscle layers and represented areas of dark brown perikaryal staining in a cell that contained a nucleus (granular stain must cover the nucleus OR encircle at least 50% of circumference of the nucleus AND at least some cytoplasmic granular brown staining must be present). If the staining overlapped or appeared as a continuous area of dark brown staining, in the presence of two distinct nuclei and cell membranes, this was counted as two cell bodies; if there was ambiguity about the presence of a nucleus, the cell was not included. Panel B shows the number of ganglia and number of neuron cell bodies per millimetre of myenteric plexus (using the pan-neuronal cell body marker HuC/D) for adult (□) and elderly (■) ascending and descending (respectively o and •) colon. Panel C is arranged similarly and shows the numbers of neuron cell bodies per millimeter of myenteric plexus stained by the antibody for ChAT or nNOS. In Panels B and C, the data are expressed as means±SEM; n=8 adult ascending, 9 elderly ascending, 9 adult descending and 10 elderly descending colons for each stain. These were analysed using analysis of variance with Sidak’s multiple comparison tests; *P<0.05; **P<0.01, only where indicated. PGP9.5, protein gene product 9.5.
Summary of ganglia and cell bodies stained by antibodies for HuC/D, ChAT and nNOS per millimetre myenteric plexus in different regions of colon from patients of different age groups
Densitometric analysis of intramuscular nerve fibre bundles
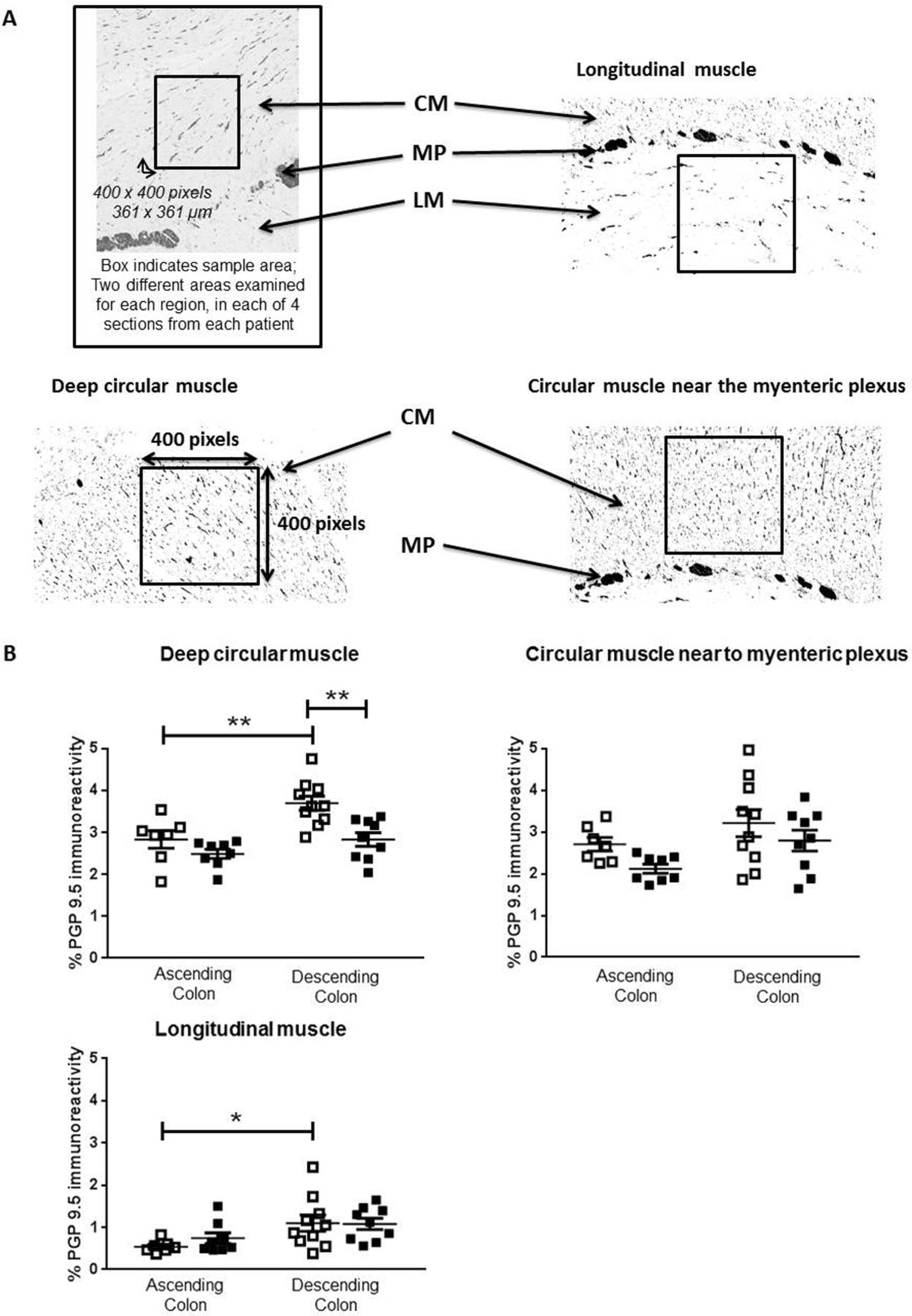
In adults, the density of anti-PGP9.5 immunostaining within muscle was generally greater in descending, compared with ascending colon (figure 4B). In ascending colon of the elderly, the density of anti-PGP9.5 staining was unchanged compared with the adults. In descending colon of the elderly, the density was reduced only in deep circular muscle (figure 4B).

Density of neuronal innervation within muscle layers. Panel A shows representative examples from protein gene product 9.5 (PGP9.5) stained paraffin-embedded sections of human colon muscle. Images were captured using NDP View version 2.3.1. Densiometric analysis was performed on filtered images converted to black and white, and the percentage density of staining measured over 400×400 pixel excerpts, as shown in the boxes, from the circular muscle (CM) near the myenteric plexus (MP), the deeper CM and the longitudinal muscle (LM), on a fixed magnification of 10×, using ImageJ. For each patient, four sections were analysed and for each region of muscle, two different fields were examined in each of the four sections by two separate assessors. Panel B shows the density of PGP9.5-positive nerve fibres in the adult (35–60 years of age; □) and elderly (≥70 years; ■) groups within ascending and descending colon, n=7 adult and n=8 elderly ascending colon and, respectively, 10 and 9 adult and elderly descending colon. Both assessors identified the same statistically significant changes or trends in the elderly (online supplementary data file 1). These data are expressed as means±SEM and were analysed using analysis of variance with Sidak’s multiple comparison test, *P<0.05; **P<0.01.
Cholinergic enzymes
In ascending colon there were no age-dependent differences in expression, specific activity or overall function of ChAT, although in descending colon ChAT function was greater in the elderly (expression unchanged), tending to increase specific activity (table 5). There were no age-dependent differences in AChE or BChE function in either region of colon (table 5).
The expression and function of choline acetyltransferase, acetylcholinesterase and butyrylcholinesterase
Mechanisms: functional
Age-dependent/region-dependent reduction in cholinergic, not nitrergic functions
In ascending colon of both age groups, L-NAME 300 µM increased the magnitude of cholinergically mediated contractions during EFS, but this was barely evident in descending colon; after-contractions were not significantly changed (figure 5 shows the magnitude of response; online supplementary file 6 shows percentage change). However, in ascending colon from the elderly, the increase in contractions was smaller compared with the adults (p<0.05; figure 5C; ascending colon from, respectively, n=14/25 adult/elderly patients; descending colon from 16/23 adult/elderly).
Supplementary file 6

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Age-dependent changes in cholinergic and nitrergic responses to electrical field stimulation (EFS) in human ascending and descending colon. EFS was applied at 5 Hz and at 50 V for 10 s every 1 min, in the absence and presence of Nω-nitro-L-arginine methyl ester hydrochloride (L-NAME) 300 µM or atropine 1 µM. Panels A and D show examples of original traces showing the effects of, respectively, L-NAME and atropine in the adult and elderly ascending colon. Similarly, panels B and E show examples of original traces for both treatments in the adult and elderly descending colon. Panels C and F show the data for each treatment in each region of adult (35–60 years of age; □) and elderly (≥70 years; ■) colon, for each tissue tested. The data are expressed as g tension/g wet weight of tissue, generated during and after termination of EFS, together with the mean±SEM contractile force. Respectively, n=14/6 and 7/4 (response generated during/after termination of EFS in the presence of L-NAME and atropine for adult ascending colon), 25/6 and 14/6 (elderly ascending colon), 16/8 and 14/8 (adult descending colon) and 23/5 and 15/5 (elderly descending colon); note that after-contractions did not always occur so their n values are smaller than for the responses measured during EFS. These were analysed using analysis of variance with Sidak’s multiple comparison tests; *P<0.05 only where indicated.
Atropine 1 µM revealed or enhanced nitrergically mediated muscle relaxation and decreased the magnitude of after-contractions (figure 5D). Comparing the age groups, there were no statistically significant differences in magnitudes of muscle relaxations or after-contractions evoked by EFS during the presence of atropine, in either region of colon (figure 5F; ascending colon from, respectively, n=6/6 adult/elderly and descending colon from 8/5 adult/elderly).
No age-dependent changes in muscle function
Advancing age did not influence contractions evoked by carbachol or Bay-K8644 in either region of colon (carbachol: respectively, n=7/7 and 8/12 ascending/descending colon from adult and elderly; Bay-K8644: n=5 each), nor relaxations evoked by SNP (table 6).
Contractions and relaxations of the muscle in adult (35–60 years of age) and elderly (≥70 years) human ascending and descending colon
Discussion
This is the first large-scale study into the effects of advancing age on both neuromuscular anatomy and functions in human colon. It identified an age-dependent decline in cholinergic, not nitrergic, function but surprisingly, only in ascending colon. Paradoxically, the numbers of myenteric neurons staining positively for anti-ChAT increased in the same region, but there was no overall change in neuron count between adult and elderly patients.
The study was needed because human colon has important differences in anatomy and functions compared with mammals in which age research is usually conducted.13 23 Further, in those age-related studies undertaken using human colon, data are inconsistent.14–16 This may be because small numbers were used, insufficient to overcome genetic differences and human variation (especially among the elderly, with longer exposure to lifestyle factors affecting bowel function). Inconsistencies are also created by studying only ascending or descending/sigmoid colon; each has different primary functions (respectively, fermentation of unabsorbed polysaccharides with water/nutrient absorption and storage of faeces prior to defection), embryological origin and molecular profile.24 Finally, to draw appropriate conclusions, analysis of changes in both anatomy and functions is desirable. Thus, the high reserve capacity of the enteric nervous system (ENS) (in animals5) means that functions of the whole organ are not represented by investigations into only structure (eg, neuron numbers) or functions of individual cell types.
The study began with a simple, large-scale assessment of the overall neuromuscular functions of ascending and descending colon, in which tissues were exposed to EFS at a frequency evoking clear neuronally mediated changes in muscle contractility. In adult colon from both regions EFS usually caused contraction, followed by ‘after-contraction’ on termination of the stimulus. The contraction during EFS was cholinergically mediated, attenuated by simultaneous inhibitory nitrergic activation, their dominance being consistent with greater numbers of myenteric neurons in human colon staining for ChAT or nNOS.25 However, in the elderly, EFS was increasingly likely to evoke muscle relaxation in ascending colon. This difference was largely uninfluenced by concurrent disease or medications, systematically analysed (unlike previous studies) as potentially confounding covariates. Thus, although it remains a possibility that other factors not measured could have influenced the data (eg, poor glycaemic control, inclusion of premenopausal/postmenopausal women among the younger group, asymptomatic diverticulosis), it seems reasonable to conclude that the data support an age-dependent change in neuromuscular functions in ascending but surprisingly not in descending colon.
Additional substudies were conducted to determine the mechanisms of change in the same cohort of patients. First, we looked for changes in numbers and phenotype of myenteric neurons. Counting of neurons requires strict application of defined criteria,18 performed blind to subject status. In this study, 30 306 myenteric neurons were counted in 36 patients; anti-HuC/D and anti-PGP9.5 assessed, respectively, myenteric ganglia/neuron cell bodies and density of nerves within the muscle. This represents the largest quantitative measurement of ENS neurons so far reported.26 The results indicated that descending colon had greater numbers of myenteric neuron cell bodies, and density of PGP9.5 staining within the muscle, compared with ascending colon (consistent with previous studies26), and increasing age did not change these numbers. However, others reported a decline in numbers of myenteric ganglia and neurons in elderly human colon27 and small intestine.28 The relatively small numbers of tissues studied and different staining techniques may explain this difference. In the largest previous study, Bernard et al 14 found a decline in numbers of myenteric neuron cell bodies in descending and sigmoid colon staining for HuC/D, ChAT and PGP9.5, but no change in numbers of cell bodies staining for nNOS. Here, the different findings may be attributed to differences in method (whole mount vs paraffin sections) and/or presentation of data. In our study, we compared two age groups. Bernard et al tested for linear trends, with most patients showing similar numbers of HuC/D-staining neurons. In spite of the decline in neuron cell bodies, these authors found no age-dependent changes in density of PGP9.5 staining of intramuscular axon bundles.
In the present study, there were relatively more nNOS-staining neurons within ascending colon (confirming others25), but no age-related changes in ascending or descending colon. These data are consistent with the suggestion that enteric nitrergic neurons do not exhibit age-related loss.6 29 In unspecified regions of colon from patients of advanced age, others report higher percentage of nitrergic neurons with no changes in numbers of calretinin-expressing neurons.16
The present study indicates that ChAT is found in ~30% of myenteric neurons within human colon (others suggest 41%–48%25; perhaps the difference reflects the quality of antibodies used and the counting criteria). A surprising finding was that in ascending, not descending colon, the numbers of nerve cell bodies detected by anti-ChAT increased in the elderly, seemingly contrasting with a generally held view that in rodents, enteric cholinergic neurons are lost with increasing age.6 Further experiments, in which ChAT, AChE and BChE were extracted from human colon, showed no consistent age-dependent differences in overall expression, specific activity or function. Similarly in rat colon, cholinesterase activities were unchanged by advancing age.30 Thus, these data, together with unchanged myenteric neuron cell bodies numbers, may be explained not by increased synthesis of ChAT, but by its increased presence within cell bodies of existing neurons. ChAT is synthesised in the soma of cholinergic neurons and transported to the terminals for ACh synthesis.31 In age-related neurodegeneration, impaired axonal transport and/or axonal dysfunction are among the earliest changes.30–33 Similarly, in mice exposed to the neurotoxic agent 5-fluorouracil, a decline in numbers of enteric ChAT-immunoreactive and other neurons was preceded by increased number of ChAT-immunoreactive neurons.34 In the present experiments, the increased detection of ChAT-immunoreactive cell bodies may therefore reflect reduced axonal transport of the enzyme away from the cell body following age-related damage to axonal transport. Experiments are required to test this hypothesis. The relatively poor signal-to-background for the ChAT antibody precludes meaningful analysis of ChAT expression by nerve terminals within the muscle, but perhaps choline uptake/ACh release could be measured during nerve stimulation in human colon.
In the first of additional functional studies, cholinergic and nitrergic functions were isolated by, respectively, L-NAME and atropine. During the presence of L-NAME, cholinergically mediated contractions evoked by EFS in ascending colon from the elderly were smaller compared with adults. A similar difference was not observed in descending colon. In the presence of atropine, nitrergically mediated relaxations were not changed with increasing age in either region of colon. These data suggest that the age-related changes in function observed in the larger functional study (see above) were due to region-dependent decline in cholinergic function, which in ascending colon enabled an intact nitrergic function to become dominant. This change was not due to reduced ability of the muscle to contract (in response to carbachol or Bay-K8644, also refuting the possibility of compensatory age-dependent changes in muscarinic receptor density35) or relax (in response to SNP). Others15 reported a greater maximal response to carbachol in sigmoid colon from females, compared with males (~60 years), with elderly females (mid-late 70s) being more sensitive; by contrast, EFS-evoked contractions and the effect of L-NAME on these responses were greater in elderly males. However, several muscle strips were used from the same patient, n values representing numbers of preparations, not patients; the potential for unintentional bias is therefore high. The present data also contrast with a reported age-dependent increase in ability of human colon muscle cells to contract in response to different ligands, including carbachol36; perhaps enzyme digestion influences how cells respond.
In summary, the size and design of the study aimed to accept, control and understand state and trait variability. The data (1) represent the most complete set of normative data for the human ENS (a recent systematic review identified marked diversity in quantification of human ENS markers among numerous small studies26), (2) refute previous observations with smaller numbers of patients, notably the existence of age-related changes in muscle function15 and in neuron numbers within the colon,16 (3) pose a new hypothesis meriting further study, namely that a region-dependent decline in cholinergic functions could be associated with increased somal ChAT staining caused by reduced axonal transport and (4) indicate that findings from laboratory animals do not always translate to humans, species-dependent variability35 37 38 being additionally hampered by gross differences in lower bowel anatomy13 and failure to consider that different bowel regions may age differently. Several questions remain unanswered. What causes the loss of cholinergic function and why only in ascending colon? Possible mechanisms include age-related intestinal inflammation,39 susceptibility to reactive oxygen species and loss of trophic support from glial cells.40 In the present study, there was no age-associated difference in gross inflammatory status, but more sensitive measurements are needed (eg, assessment of age-dependent changes in ‘proinflammatory’ status of macrophages41 and analysis of immune activity of dendritic cell subsets in different regions of human colon42). Interestingly, the high content of living bacteria in ascending colon may change in composition among the elderly, promoting low-grade inflammation (inflammaging)43 and increasing mucosal permeability to allow potentially damaging substances to penetrate into the wall of ascending colon. Precedents for age-related, region-specific reductions in cholinergic functions (and increased ChAT; see Discussion above) exist in brain of patients with mild cognitive impairment44 45 and in aged rats.46
Notably, none of the patients studied had previous clinical diagnosis of constipation (any history of self-reporting constipation was not obtained). It is, therefore, not possible to conclude that declined myenteric cholinergic function within the ascending colon necessarily leads to clinical constipation. Indeed, other degenerative changes are suggested to contribute to age-related constipation (including changes in inhibitory junction potentials within the descending colon ENS,47 enteric sensory5 and extrinsic sensory innervations48). Nevertheless, the reduced cholinergic function does support the view that the ENS reserve capacity is reduced in the elderly, increasing sensitivity to lifestyle changes affecting intestinal functions (eg, diet, exercise, medications; see the Introduction section) and raising the probability of developing constipation. A similar argument has been suggested for cholinergic function in the aged brain, in which the consequence of a gradual decline in synthesis and/or release of ACh from synaptosomes49 only becomes apparent when nerves are stressed or damaged.31
Acknowledgments
We thank other consultant colorectal surgeons, particularly Chris Chan, and pathologists at Barts Health for assisting with identification of patients for recruitment into this study and for providing tissue suitable for laboratory use.
References
Footnotes
JB and VWSK contributed equally.
Contributors JB and VWSK conducted the experiments, SE and CHK facilitated the identification, collection and governance of human tissue collection, TD-S, JEM and MJS provided methodology guidance, GJS wrote the manuscript and all authors participated in its construction and refinement, particularly CHK, VWSK, JB and MJS.
Funding JB was supported by the Bowel and Cancer research charity, the Dunhill Medical Trust (grant no: R382/1114 to GJS, CHK and MJS) and an EMBO scholarship to visit the Karolinska Institutet (to JB). VWSK was supported by the research into ageing fund, set up and managed by AgeUK (grant to GJS). AP and SE are supported by Takeda Pharmaceuticals.
Competing interests GJS is currently in receipt of funding from Takeda Pharmaceuticals, BBSRC together with GlaxoSmithKline, Benevolent and the Dunhill Foundation. He acts as an advisor to Takeda Pharmaceuticals and to Zealand Pharma.
Patient consent Not required.
Ethics approval Approved by the local ethics committee (REC 10/H0703/71).
Provenance and peer review Not commissioned; externally peer reviewed.