Article Text

Abstract
Objective The microbiome directly affects the balance of pro-inflammatory and anti-inflammatory responses in the gut. As microbes thrive on dietary substrates, the question arises whether we can nourish an anti-inflammatory gut ecosystem. We aim to unravel interactions between diet, gut microbiota and their functional ability to induce intestinal inflammation.
Design We investigated the relation between 173 dietary factors and the microbiome of 1425 individuals spanning four cohorts: Crohn’s disease, ulcerative colitis, irritable bowel syndrome and the general population. Shotgun metagenomic sequencing was performed to profile gut microbial composition and function. Dietary intake was assessed through food frequency questionnaires. We performed unsupervised clustering to identify dietary patterns and microbial clusters. Associations between diet and microbial features were explored per cohort, followed by a meta-analysis and heterogeneity estimation.
Results We identified 38 associations between dietary patterns and microbial clusters. Moreover, 61 individual foods and nutrients were associated with 61 species and 249 metabolic pathways in the meta-analysis across healthy individuals and patients with IBS, Crohn’s disease and UC (false discovery rate<0.05). Processed foods and animal-derived foods were consistently associated with higher abundances of Firmicutes, Ruminococcus species of the Blautia genus and endotoxin synthesis pathways. The opposite was found for plant foods and fish, which were positively associated with short-chain fatty acid-producing commensals and pathways of nutrient metabolism.
Conclusion We identified dietary patterns that consistently correlate with groups of bacteria with shared functional roles in both, health and disease. Moreover, specific foods and nutrients were associated with species known to infer mucosal protection and anti-inflammatory effects. We propose microbial mechanisms through which the diet affects inflammatory responses in the gut as a rationale for future intervention studies.
- diet
- intestinal microbiology
- meta-analysis
- inflammatory bowel disease
- irritable bowel syndrome
Data availability statement
All relevant data supporting the key findings of this study are available within the article and the supplementary files. Raw metagenomic sequencing reads and extended phenotypic data are available from the European Genome-phenome Archive data repository: 1000 IBD cohort [EGAD00001004194] and LifeLines Deep cohort [EGAD00001001991]. Codes used for generating the microbial profiles are publicly available at:[https://github.com/WeersmaLabIBD/Microbiome/blob/master/Protocol_metagenomic_pipeline.md]. All statistical analysis scripts are written in R and can be found here: https://github.com/WeersmaLabIBD/Microbiome/blob/master/Diet_Microbiome.md.
This is an open access article distributed in accordance with the Creative Commons Attribution 4.0 Unported (CC BY 4.0) license, which permits others to copy, redistribute, remix, transform and build upon this work for any purpose, provided the original work is properly cited, a link to the licence is given, and indication of whether changes were made. See: https://creativecommons.org/licenses/by/4.0/.
Statistics from Altmetric.com
Significance of this study
What is already known on this subject?
Western diet and low-grade intestinal inflammation are implicated in a growing number of immune-mediated inflammatory diseases.
Diet quantity, content and timing play a major role in shaping gut microbial composition and function.
Dysbiosis, shifts in metabolites and translocation of microbial products contribute to immune activation.
Research has been focused on anti-inflammatory properties of isolated compounds, with limited efficacy.
What are the new findings?
Diet-gut microbiome associations are consistent across patients with intestinal disease (Crohn’s disease, UC, IBS) and the general population.
Higher intake of animal foods, processed foods, alcohol and sugar, corresponds to a microbial environment that is characteristic of inflammation, and is associated with higher levels of intestinal inflammatory markers.
Plant-based foods are linked to short-chain fatty acid (SCFA)-producers, microbial metabolism of polysaccharides and a lower abundance of pathobionts.
Significance of this study
How might it impact on clinical practice in the foreseeable future?
Modulation of gut microbiota through diets enriched in vegetables, legumes, grains, nuts and fish and a higher intake of plant over animal foods, has a potential to prevent intestinal inflammatory processes at the core of many chronic diseases.
Whole food-based dietary patterns could increase the anti-inflammatory capacity of nutrients through synergistic effects on the gut microbiome.
Sources of n-3 PUFAs (omega-3 polyunsaturated fatty acids) and polyphenols may be used to potentiate the abundance of SCFA-producers.
Replacement of animal protein by plant protein has a potential to reduce intestinal inflammatory processes by targeting microbial pathways involved.
Introduction
The gut microbiome directly affects the balance of pro-inflammatory and anti-inflammatory responses in the intestine. Microbial competition for nutrients plays a key role in controlling this balance.1 Inflammatory bowel disease (IBD) is the archetypical disease in which the homoeostasis between the gut microbiota and the intestinal immune system is lost. Beyond the local immune responses, the gut microbiota also affect systemic immune components and are implicated in a growing number of immune-mediated inflammatory diseases (IMIDs), ranging from diabetes to arthritis and systemic lupus erythematosus.2 Gut dysbiosis and associated inflammation have also been implicated in cancer and cardiometabolic disorders.3 4 Epidemiological studies uncovered several dietary factors associated with the onset of these diseases. However, the mechanisms underlying this relationship remain largely unknown.
As microbes rely on dietary substrates in the intestine, the gut microbiome is often proposed as a mediator through which foods exert their pro-inflammatory and anti-inflammatory effects. For example, animal experiments demonstrated that foods containing high levels of saturated fats,5 dietary heme,6 sugar,7 salt8 and low levels of fibre1 induce inflammation and autoimmunity through microbial mechanisms such as induction of T-helper 17 (TH17) cells. Other studies in mice and humans implicated that ingredients added during food processing including dietary emulsifiers,9 antimicrobial additives10 and artificial sweeteners,11 promote gut permeability and intestinal inflammation through an increase in mucolytic bacteria and endotoxins. In contrast, a high intake of tryptophan12 and fibre13 generally leads to immune states associated with colonic health.
The knowledge on pro-inflammatory and anti-inflammatory capacities of single compounds is increasing through functional experiments. However, there is still limited understanding of how whole foods and dietary patterns impact the gut microbiota and the host and if these impacts are different in the healthy versus the inflamed intestine. In contrast to very few food-based interventions, there have been numerous clinical trials of single nutrients. While it is easier to intervene with a pill rather than with dietary change, these trials do not acknowledge interactions of nutrients within their food matrix, which may explain the contradictory and limited effects seen.14 Understanding the synergies found in whole foods in the context of dietary patterns may result in more effective nutrition research and policy.
Long-term dietary interventions may be most suited for the modulation of the gut microbiota. Although extreme short-term dietary changes may still derange the gut microbiota,15 16 there is a tendency for microbial resilience in adults that correlates with long-term habitual diet,17–19 providing a constant source of dietary substrates and continuously shaping the gut ecosystem.
In this study we aimed to investigate the complex relationship between habitual diet, gut microbiota and intestinal inflammation in humans. To do so, we associated 173 dietary factors, moving from dietary patterns to specific foods and macronutrients, with the gut microbiome composition and function of 1425 individuals across four cohorts: Crohn’s disease, ulcerative colitis, irritable bowel syndrome and the general population. Analyses were performed in each cohort, followed by a meta-analysis to explore replicability of diet-microbiome associations in different disease contexts. We propose pro-inflammatory and anti-inflammatory mechanisms through which specific foods and dietary patterns could affect inflammatory responses in the gut as a rational basis for designing dietary interventions.
Materials and methods
Study design and cohort description
We associated the diet to the gut microbiome composition and function of 1425 individuals from the general population and patients with intestinal diseases, using two independent cohorts from the northern Netherlands. Cohort 1 consists of 331 patients with IBD from the 1000IBD cohort of the University Medical Center Groningen (UMCG).20 Patients were diagnosed by their treating physician based on accepted radiological, endoscopic and histopathological evaluation and were classified as either Crohn’s disease or ulcerative colitis. Cohort 2 consists of 1094 individuals from the Dutch general-population-based cohort LifeLines DEEP.21 The LifeLines DEEP cohort comprises 223 individuals with irritable bowel syndrome (IBS) according to a symptom questionnaire based on Rome-III criteria. Due to known differences in the gut microbiome composition and dietary preferences between healthy individuals and patients with IBD and IBS, participants were divided into four sub-cohorts, namely Crohn’s disease (CD, n=205), ulcerative colitis (UC, n=126), IBS (n=223) and healthy controls (HC, n=871) (table 1).22–25 To explore consistency and heterogeneity across these different contexts, all analyses were performed separately per cohort, followed by a meta-analysis.
Cohort characteristics
Supplemental material
Fecal sample collection and processing
For each participant one stool sample was collected. Participants were asked to collect and immediately freeze their sample at home. Samples were then picked up, transported on dry ice and stored at –80°C. All samples were processed according to the same pipeline in one laboratory (UMCG, Groningen). Intestinal inflammatory markers were measured in the same sample. The protocol for faecal sample collection and profiling of gut microbiota was previously published and is summarised below.22 26
DNA extraction and sequencing
DNA extraction was performed using the AllPrep DNA/RNA Mini kit (Qiagen; cat. #80204) combined with mechanical lysis. Metagenomic shotgun sequencing was performed at the Broad Institute (Cambridge, Massachusetts, USA) using Illumina HiSeq platform as previously described.22 26 Low-quality reads were filtered out at the sequencing facility. KneadData toolkit (V.0.5.1) was used to trim the raw metagenomic reads to PHRED quality 30 and to remove Illumina adapters.
Metagenomic profiling and filtering of samples
Reads aligning to the human genome (GRCh37/Hg19) were removed using KneadData integrated Bowtie2 tool (V.2.3.4.1), and the quality of processed metagenomes was examined using the FastQC toolkit (V.0.11.7). Functional profiles were calculated using HUMAnN2 (V.0.10.0). The taxonomic composition was evaluated using MetaPhlAn2 (V.2.2). Microbes and microbial functions that were present in less than 10% of samples and microbes with a relative abundance lower than 0.01% were not included in subsequent analyses. Samples with a sequencing depth below 10 million reads were removed. After filtering, 1425 samples remained for the analyses. Arcsine square-root transformations for taxonomic abundances and logarithmic transformation for pathways were used as normalisation methods. A Grubbs’ test was conducted to remove outliers. A link to the pipeline and analyses scripts is provided under the data availability. The statistical tests and terminology are further described in online supplemental table 1.
Supplemental material
Dietary assessment and processing of questionnaires
Dietary intake was assessed using a semi-quantitative Food Frequency Questionnaire (FFQ) that was collected concordantly with the faecal sample. The FFQ was designed and validated by the division of Human Nutrition of Wageningen University, using standardised methods.27 It assesses how often a food item was consumed over the previous month on a 7-item scale, along with the usual amount taken. The average daily nutrient intake was calculated by multiplying frequencies of consumption by portion size and nutrient content per gram as indicated in the Dutch Food Composition database (NEVO). Specific food items were aggregated into 25 food groups in grams per day, for example, a group of dairy composed of 21 single products such as yoghurt, buttermilk and milk21 (online supplemental table 2). We performed energy adjustment of the dietary intake using the nutrient density method.28
Descriptive statistics
χ2 tests for categorical variables and Wilcoxon-Mann-Whitney test (WMW test) for continuous data were performed to calculate statistically significant differences between cohorts. Differences in age, sex, body mass index (BMI) and sequencing depth were tested between each group and all other groups. Differences in food intake and medication use were tested between each group and HCs. The gut microbiota composition of each cohort has been described previously.22 26
Identification of dietary patterns and microbial clusters
Similar to microbiome data, food intake data is often zero-inflated with internal correlations of features, implying that individuals seldom eat unique foods but often consume meals with conventional food combinations.29 In order to identify stable dietary patterns, we performed unsupervised hierarchical clustering of the dietary intake data (in the units of gram per day) based on squared Euclidean distances. Subsequently, clustering was performed on microbial pathways using squared Euclidean distances, and on species abundance using Bray-Curtis dissimilarity as between-sample metrics. To ensure that the existence of clusters was stable and not dependent on just one set of parameters, different clustering heights and distance metrices were tested and clustering was performed in each cohort separately. A dendrogram (hierarchical tree) was visualised and clusters were defined by cutting branches off the dendrogram. As best cut, a height of 53 for the food tree, and a height of 0.8 for the species tree was identified. As the identification of clusters was stable across cohorts, clustering could ultimately be performed on the joint data set. Centroids were calculated for each participant as the mean consumption or mean abundance of all variables within a cluster.
Association analyses per cohort
Next, we explored associations between dietary intake and microbial and pathway abundance in each cohort. For each food item or nutrient, we constructed a multivariate linear model of the food consumption adjusted for caloric intake, versus the relative abundance of taxa and pathways. Age, sex and sequencing read depth were added as covariates, represented as:
Lm: Microbial feature (taxa/pathway)~intercept + Food/nutrient + Age + Sex + Seq.depth + Cohort (HC/IBS/CD/UC)
The same model was used to test associations between dietary patterns and microbial clusters using the centroids (means) of each cluster. Since clustering was performed on the unadjusted food intake in grams per day, caloric intake (kcal) was added as a covariate:
Lm: Microbial cluster (species/pathways)~intercept + Food cluster + Age + Sex + Seq.depth + Total kcal + Cohort (HC/IBS/CD/UC)
Subsequently, dietary patterns were correlated to chromogranin A (CgA) and faecal calprotectin (Fcal) as surrogate markers for intestinal inflammation, using the same approach.
Lm: Inflammatory maker (Fcal/CgA)~intercept + Food cluster + Age + Sex + Seq.depth + Total kcal + Cohort (HC/IBS/CD/UC)
All analyses were adjusted for multiple testing using the Benjamini-Hochberg method as implemented in the p.adjust function in R. A false discovery rate (FDR) of <0.05 was defined as the significance cut-off.
We also explored the influence of metabolic factors, choosing phenotypes that were available in our cohort, including BMI, smoking status, hypertension (indirectly defined by the use of antihypertensives), diabetes (by the use of antidiabetics) and hyperlipidaemia (by the use of statins). The statistics thereof are provided per food-microbiome association in the online supplemental tables.
Supplemental material
Supplemental material
Supplemental material
Supplemental material
Supplemental material
Supplemental material
Supplemental material
Cross-disease meta-analysis
Next, we combined the results obtained per cohort in a meta-analysis framework in order to explore diet-microbiome relations that were significant and consistent across all cohorts. The inverse-variance method was used to calculate combined meta z-scores and corresponding meta p values. Multiple testing correction was performed per food item or food cluster for all tested taxa and pathways.
A Cochran’s Q test was conducted to measure heterogeneity between cohorts using the function metagen (meta R package (V.4.8–4)). An FDR of <0.05 and a heterogeneity p value of >0.05 in the meta-analysis were considered significant.
Results
Descriptive statistics
Patients with CD and individuals with IBS were younger than patients with UC and HC (mean (SD): CD 40.6 (14.2), IBS 41.4 (12.2), UC 46.65 (14.83) and HC 45.5 years of age (13.5)) (table 1). The average caloric intake was lower in IBS compared with HCs, reflecting a greater proportion of women among patients with IBS in line with the recognised 2:1 female-to-male ratio (IBS 1797.6 (476) vs HC 1976.2 kcal/day (621.3); IBS 83% vs HC 52% female). Individuals with IBS consumed less bread, potatoes, cheese, spreads and yoghurt drinks than HCs, which was reflected by a lower protein and plant protein intake (protein: IBS 68.1 (16.1) vs HC 74.8 g/day (21.3); plant protein: IBS 27.3 (7.7) vs HC 30.8 g/day (10.5)). Protein and vegetable intake was also lower in CD compared with healthy controls (protein: CD 67.1 (24.1) vs HC 74.8 g/day (21.3); group vegetables: CD 98.1 (76.3) vs HC 109.4 g/day (64.5)). Lower protein and fibre intakes in CD and IBS have been demonstrated before.24 25Patients with CD consumed more soft drinks than HCs as previously shown,24 whereas alcohol intake was higher in HCs. Cohort characteristics and differences in average food group and macronutrient intakes are given in table 1. Full descriptive statistics of all single food items are provided in online supplemental table 2. Based on this data, we corrected our analyses for age, sex, sequencing read depth and energy intake and performed analyses separately per cohort, followed by a meta-analysis and a Cochran’s Q test.
Results of the cluster analyses
Unsupervised clustering identifies common dietary patterns and microbial clusters
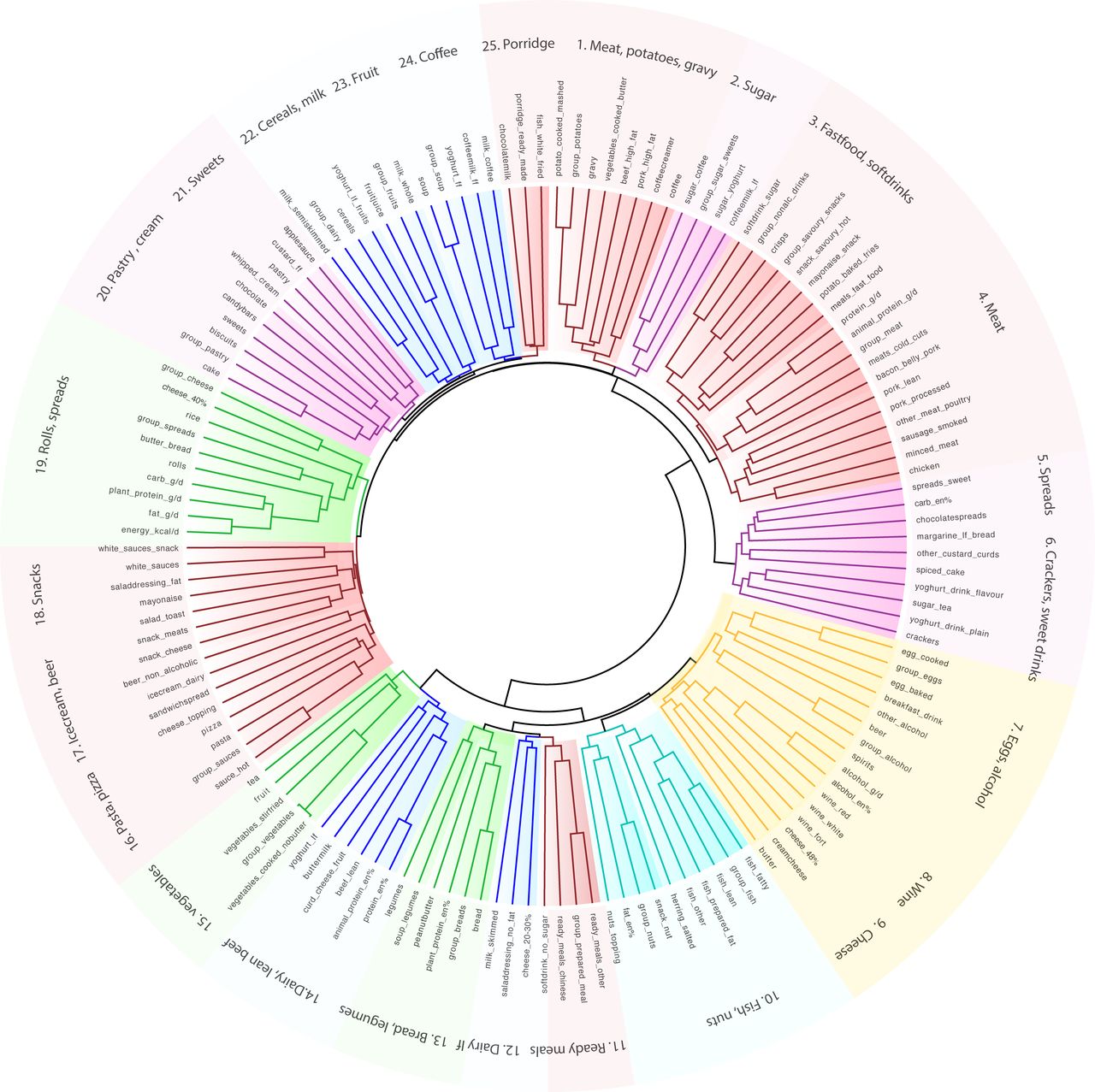
Unsupervised hierarchical cluster analyses, irrespective of disease status, identified 25 clusters of common food pairings (figure 1). For example, cereals clustered with dairy and meat clustered with potatoes and gravy. French fries, meat, savoury snacks, mayonnaise and soft drinks formed a typical ‘fast food’ cluster.

Unsupervised dietary cluster analysis reveals common food patterns. Cladogram showing clustering of the dietary intake into 25 patterns. Food frequency questionnaires were used to assess the diet of 1425 individuals comprising healthy controls (n=871), individuals with irritable bowel syndrome (n=223), Crohn’s disease (n=205) and ulcerative colitis (n=126). Unsupervised hierarchical clustering was performed using squared Euclidean distances.
The same analysis performed on taxonomical and functional abundances, identified 29 clusters of species with similar functions and 31 clusters of pathways of similar classes. Clusters are numbered sequentially throughout the manuscript and tables. For example, we found a cluster of commensal obligate anaerobes capable of short-chain fatty acid (SCFA) production (S1: Bifidobacterium, Eubacterium, Dorea, Ruminococcus, Coprococcus, Subdoligranulum, and Faecalibacterium spp). Another cluster was formed by Escherichia coli, Parabacteroides and Bacteroides fragilis. Bifidobacterium dentium clustered with various Streptococcus species that are dominant in the oral microbiome. Other clusters were composed of pathways associated with growth and survival of facultative anaerobes such as E. coli that have been linked to intestinal inflammation (P2: aerobic respiration, synthesis of lipopolysaccharides (LPS), heme, enterobacterial common antigen, menaquinol; P9: synthesis of enterobactin, O-antigen, quinones). The composition of clusters can be found in online supplemental table 3.
Results of the meta-analysis
Meta-analysis shows similar signals across intestinal disease cohorts and controls
The meta-analysis identified significant associations between 13 dietary patterns, 24 microbial groups and markers of intestinal inflammation that were consistent across the four cohorts (FDR<0.05, p-Cochran’s-Q>0.05, online supplemental tables 4–6).
Moreover, there were 393 associations between 123 unique microbial taxa and 61 food items in the meta-analysis of individual taxa and foods (FDR<0.05, p-Cochran’s-Q>0.05, online supplemental table 7). Strikingly, 280 out of 393 results had the same direction (beta-coefficient, coef) in all cohorts, suggesting shared signals across different intestinal diseases (CD, UC and IBS) and healthy individuals. Moreover, including BMI and use of anti-diabetics, anti-hypertensives and statins as additional covariates in the model, replicated 82.2% of the results, demonstrating the robustness of the meta-analysis approach.
Plant protein, carbohydrates and red wine accounted for most associations with microbial taxa (23, 23 and 20 associations), followed by fish, nuts and animal-derived protein (17, 16 and 15 associations). Species most affected by the diet were Lactobacillus sakei (13), Roseburia hominis GCF_000225345 (12), Faecalibacterium prausnitzii (11), Bifidobacterium adolescentis (11) and Ruminococcus obeum (11 associations). In the same analysis, 282 pathways were related to 41 food items and nutrients. Tea, sugar used in tea, potatoes and sauces accounted for most associations with metabolic functions (48, 186, 62 and 155 associations) while they had no significant impact on microbial taxa (online supplemental table 8).
Lastly, the per-cohort analysis also showed disease-specific results that were not significant in the meta-analysis and mainly concerned species that are enriched, such as Sutterella wadsworthensis and Bilophila, or depleted in CD, UC or IBS, such as Bifidobacterium adolescentis (online supplemental table 7).
Clusters of breads, legumes, fish and nuts show a consistent negative association with several pro-inflammatory pathways
A food cluster comprising breads and legumes, and a cluster of fish and nuts, were negatively associated with groups of pathways involved in the synthesis of growth factors, endotoxins and cell wall components (P2, P9, P22, online supplemental table 4, figure 2A). Moreover, we observed a negative association between the fish and nuts cluster and pathways for the synthesis of L-tyrosine, L-phenylalanine, terpenoids, quinones and fatty acids, a profile predictive of E. coli (P25, P24, P20, P5, figure 2A). The bread and legumes cluster was associated with a lower abundance of an E. coli, Bacteroides fragilis and Parabacteroides cluster (S13: FDR=0.015, coef=−0.066, online supplemental table 5, figure 2B). Conversely, the cluster of breads and legumes, and the cluster of fish and nuts, were associated with a higher abundance of pathways involved in the synthesis of acetate and the urea cycle for detoxification of ammonium (P4, figure 2A).

Consistent associations of dietary patterns with clusters of pathways (A) and species (B) in the cross-disease meta-analysis. Forest plot showing consistent results between dietary patterns and microbial clusters in a cross-disease meta-analysis of 1425 individuals spanning four cohorts (FDRMeta<0.05, p-Cochran’s-Q>0.05). Dots indicate pooled results of the meta-analysis; black lines indicate CIs. Dot size indicates the significance of the association (FDR-corrected p value). X-axis represents coefficients. Unsupervised hierarchical clustering was performed on dietary intake, species and pathway abundance, using squared Euclidean and Bray-Curtis distance. In each cohort, a multivariate linear model of food clusters versus microbial clusters was constructed, adding age, sex, sequencing depth and caloric intake as covariates. An inverse-variance meta-analysis was conducted on results obtained per cohort, followed by multiple testing correction and a Cochran’s Q test. AA, amino acid; ECA, enterobacterial common antigen; FA, fatty acid; FDR, false discovery rate; ferment, fermentation; LPS,lipopolysaccharides; spp, species.
Consumption of nuts, oily fish, fruit, vegetables and cereals is linked to a higher abundance of SCFA-producers
Also individually, these food items were related to several commensals capable of SCFA production (figure 3). For example, Faecalibacterium prausnitzii abundance was positively associated with consumption of fruits (FDR=0.005, coef=0.1), red wine (FDR=0.0003, coef=0.441) and oily fish (FDR=0.037, coef=1.695), but showed a negative association with high-sugar foods (soft drinks: FDR=0.028, coef= -0.131; sweets: FDR=0.039, coef= -0.669) (figure 3A). Roseburia hominis abundance was positively associated with nuts (FDR=3.80×10–05, coef=0.629), oily fish (FDR=0.0002, coef=1.057), vegetables (FDR=0.007, coef=0.079), legumes (FDR=0.029, coef=0.402), cereals (FDR=0.014, coef=0.485) and plant protein (FDR=1.17×10–05, coef=3.567) (figure 3B). These bacteria are known to have anti-inflammatory effects and provide protection of the intestinal mucosa through fermentation of fibre and pectins to acetate and butyrate.13 Details of each taxon and pathway, including statistics for each cohort, as well as the meta-analysis, are provided in online supplemental tables 7 and 8.

Dietary factors associated with Faecalibacterium prausnitzii (A) and Roseburia (B) relative abundance in the meta-analysis. Heatmap showing significant and consistent results of the cross-disease meta-analysis between individual foods and relative abundance of (A) Faecalibacterium prausnitzii and (B) Roseburia sp (FDR<0.05, p-Cochran’s-Q>0.05). Dietary intake was assessed by Food Frequency Questionnaires. Energy adjustment was performed by the nutrient density method. For each food item, we constructed a multivariate linear model of the food intake versus taxa and pathways, adding age, sex and sequencing depth as covariates. Association analyses were performed per cohort, followed by an inverse-variance meta-analysis, multiple testing correction and a Cochran’s Q test. carb; carbohydrates; CD, Crohn’s disease; en-%, energy-per cent; FDR, false discovery rate; g/d, gram per day; IBS, irritable bowel syndrome; nut_d, nuts added to dinner; sp, species; UC, ulcerative colitis. Red, positive association; blue, negative association. Colour density indicates significance of the association (FDR-corrected p value).
Red wine is associated with a higher abundance of several acetate and butyrate producers but with a lower Bifidobacterium abundance
A cluster of acetate and butyrate producing species (S1) was positively associated with a cluster of different types of wine (FDR=0.002, coef=0.036, figure 2B). Specifically, red wine was linked to higher abundances of Faecalibacterium prausnitzii, Eubacterium hallii, Ruminococcus obeum, Ruminococcus lactaris, Anaerostipes hadrus and Alistipes putredinis (all FDR<0.05, p-Cochran’s-Q>0.05, online supplemental table 7). Conversely, red wine intake showed a negative association with Bifidobacterium abundance, a SCFA-synthesising commensal (FDR=0.007, coef=−0.933).
Alcohol and sugar intake is associated with a higher abundance of quinone synthesis pathways
Consumption of spirits (pure grain-based alcohol) was associated with a higher abundance of quinone synthesis pathways, that we previously reported to be enriched in IBD (PWY-5840, PWY-5850, PWY-5860, PWY-5862, online supplemental table 8),22 although after correcting for metformin use, this was not nominally significant anymore (FDR=0.094). Moreover, we observed a negative association of a pyruvate to propanoate fermentation pathway with total alcohol intake in energy-% (P108-PWY, FDR=0.0103, coef=−0.067). In contrast to alcohol and sugar, plant protein intake was negatively associated with quinone synthesis (PWY-5862, PWY-5896, online supplemental table 8, figure 4A).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Microbial metabolic pathways (A) and taxa (B) associated with plant protein intake in the meta-analysis. Heatmap showing significant and consistent results of the cross-disease meta-analysis between plant protein intake and the relative abundance of (A) metabolic pathways and (B) taxonomical abundance of the gut microbiome (FDR<0.05, p-Cochran’s-Q>0.05). Dietary intake was assessed by Food Frequency Questionnaires. For each food item, we constructed a multivariate linear model of the food intake versus taxa and pathways, adding age, sex and sequencing depth as covariates. Association analyses were performed per cohort, followed by an inverse-variance meta-analysis, multiple testing correction and a Cochran’s Q test. CD, Crohn’s disease; FDR, false discovery rate; IBS, irritable bowel syndrome; UC, ulcerative colitis. Red, positive association; blue, negative association. Colour density indicates significance of the association (FDR-corrected p value).
Coffee intake is associated with a higher Oscillibacter abundance
Coffee consumption was significantly associated with Oscillibacter abundance (FDR=8.37×10–05, coef=0.022), heterolactic fermentation and various glycolytic pathways (PWY-6969, FUC-RHAMCAT-PWY, P122-PWY, online supplemental tables 7 and 8).
Lactic acid bacteria and fermentation to butanediol are consistently associated with consumption of fermented dairy
Consumption of fermented dairy like buttermilk and yoghurt showed strong associations with lactic bacteria, as previously shown,26 as well as with the fermentation of pyruvate to butanediol (P125-PWY) and peptidoglycan synthesis (PWY-6471) (online supplemental tables 7 and 8).
Plant-based food consumption is associated with higher synthesis and conversion of essential nutrients by the gut microbiota
Total intake of plant-derived protein was positively associated with pathways involved in the synthesis of SCFA (P108-PWY, P162-PWY), thiamin (PWY-6897), biotin (PWY-5005), flavin (PWY-6168), vitamin B6 (PWY0-845; PYRIDOXSYN-PWY) and L-ornithine (ARGININE-SYN4-PWY), and the degradation of sugar derivates (PWY-6531, HEXITOLDEGSUPER-PWY) (figure 4A). Participants consuming higher amounts of potatoes showed a higher abundance of starch degrading pathways (PWY-6731: FDR=0.038, coef=0.003).
Plant-derived and animal-derived foods and nutrients show inverse taxonomical associations
Already at the higher taxonomic levels we observed opposite relations of animal-based and plant-based foods and nutrients. While total intake of animal protein and fat was associated with a higher Firmicutes abundance, a negative association was found for plant protein and carbohydrate intake (FDR=1.30×10-05, coef=3.646; FDR=0.042, coef=2.936; FDR=0.003, coef=−6.081; FDR=4.67×10-07, coef=−1.735, respectively). Firmicutes-dominated communities have been observed in omnivores of the general population.19 While plant protein and bread intake were consistently linked to a higher Bifidobacterium abundance (FDR=0.049, coef=4.982; FDR=0.004, coef=0.815, figure 4B), total fat and animal protein intake, cheese and fish were associated with a lower Bifidobacterium abundance (animal protein: FDR=1.30×10–05, coef=−4.113), except for Bifidobacterium dentium. Bifidobacterium dentium, a dominant species of the upper gastrointestinal (GI) tract, showed a positive association with consumption of meat, animal protein and butter (FDR=0.001; FDR=0.048, FDR=1.91×10–05).
Furthermore, we observed higher abundances of Erysipelotrichaceae, Ruminococcus species of the Blautia genus and Streptococcus species with animal protein, while the opposite direction was found for plant protein intake (online supplemental table 7, figure 4B).
Fast food consumption is associated with higher abundances of Blautia, Lachnospiraceae bacteria and Clostridium bolteae
We observed significant positive associations between the consumption of fast food and savoury snacks and the abundance of Blautia, Lachnospiraceae bacteria and Clostridium bolteae in line with previous reports.30 A fast food cluster consisting of meats, french fries, mayonnaise and soft drinks, showed a positive association with a cluster of Clostridium bolteae, Coprobacillus and Lachnospiraceae bacteria (1_4_56FAA and 2_1_58FAA) (FDR=0.040, coef=0.057, figure 2B). Moreover, the cluster of fast food showed a positive association with a Ruminococcus gnavus and Lachnospiraceae bacterium 1_1_57FAA cluster in IBS and HCs, which was not significant in CD and UC in the meta-analysis (FDRHC=4.99×10–5, FDRIBS=3.22×10–5, FDRCD=0.490, FDRUC=0.761, FDRmeta=4.69×10–6, coefmeta=0.128, p-Cochran’s-Q=0.02). This finding reflects a difference in the effect size, with Ruminococcus gnavus being already enriched in CD and UC as compared with controls.31 Moreover, fast food meals and savoury snacks were positively associated with Parabacteroides johnsonii, Lactobacillus sakei, Lachnospiraceae bacterium 1_1_57FAA and the Ruminococcus genus across all cohorts (online supplemental table 7).
Per cohort analysis also reveals disease-specific results
While the primary purpose of this study was to perform a meta-analysis across different conditions, heterogeneity tests and linear models performed separately per cohort also revealed disease-specific results for species that are enriched in IBD or IBS compared with controls22 23 (online supplemental tables 4–8). We observed a positive association of the abundance of bile tolerant bacteria such as Sutterella wadsworthensis, Bilophila, Bacteroides and Alistipes spp. with the consumption of fast food or ready meals in CD, UC and IBS. This association was not statistically significant in the HC-group in which these taxa have a lower abundance, but showed the same directionality (coefficient) (online supplemental tables 7). In IBS, consumption of buttermilk, bread and cereals was associated with a lower abundance of hydrogen producing Dorea spp and with a higher Bifidobacterium abundance (all FDRIBS<0.05). In UC, Methanobacter smithii abundance was positively associated with whole milk, butter, sauces, sweets and alcoholic drinks (all FDRUC<0.05). In CD, Bacteroides vulgatus abundance was associated with milk, animal protein and fat intake (FDRCD=0.002, coef =1.053; FDRCD=0.009, coef =4.561, FDRCD=0.01, coef =2.078).
Dietary patterns are associated with intestinal inflammatory markers
Lastly, we observed significant positive associations of Fcal with the cluster comprised of fast food (FDR=4.14×10–4, coef=0.242) and a cluster comprised of high-fat meat, potatoes and gravy (FDR=0.003, coef=0.218), that were consistent in the meta-analysis. By contrast, we saw a negative association of Fcal with the cluster comprised of fish and nuts (FDR=0.038, coef=−0.102) and of CgA with a cluster of breads and legumes (FDR=0.005, coef=−2.484) supporting the pro-inflammatory and anti-inflammatory role of the microbial features that were associated with these foods (online supplemental tables 6).
Discussion
In this study we have shown how habitual dietary choices can impact the human gut ecosystem and its inflammatory potential by studying the relations between unsupervised dietary patterns, intestinal inflammatory markers and gut microbial composition and function across four cohorts. We identified significant associations that replicate across patients with Crohn’s disease, ulcerative colitis, irritable bowel syndrome and the general population, implying a potential for microbiome-targeted dietary strategies to alleviate and prevent intestinal inflammation.
We showed that dietary patterns comprising legumes, breads, fish and nuts are associated with a lower abundance of clusters of opportunistic bacteria, pathways for the synthesis of endotoxins and inflammatory markers in stool. Higher proportions of these bacteria and pathways have been implicated in IBD and colorectal cancer combined with inflammation through their metabolites such as LPS.22 32 Conversely, we observed higher abundances of commensals such as Roseburia, Faecalibacterium and Eubacterium spp with the consumption of nuts, oily fish, fruits, vegetables, cereals and red wine across all cohorts. These bacteria are known for their anti-inflammatory effects in the intestine through fermentation of fibre to SCFAs.13 A dietary pattern that is traditionally high in these foods is the Mediterranean diet which has been linked to a lower IBD-risk in retrospective studies.33
Accumulating literature demonstrates an anti-inflammatory role of polyphenol-rich foods such as coffee, tea, red wine and fruit. We observed a higher Oscillibacter abundance and a lower abundance of pro-inflammatory pathways with coffee consumption. Increases in Oscillibacter have been shown on administration of tea-phenols or berry-phenols at the expense of potentially pathogenic species in mice.34 Moreover, we saw positive associations of red wine intake with several acetate and butyrate producers. Red wine polyphenols have been shown to increase Faecalibacterium prausnitzii and Roseburia hominis while reducing E. coli and Enterobacter cloacae abundance, C-reactive protein (CRP) and cholesterol levels in healthy and obese individuals.35 36 In contrast, total alcohol intake and spirits were associated with pro-inflammatory pathways in our study. Alcohol-induced reduction of Bifidobacteria and higher endotoxin production has been suggested to increase intestinal inflammation in patients with GI cancers and liver disease.37 Together, these findings support the earlier finding that moderate red wine intake is linked to higher microbial diversity, a parameter of gut health,26 38 while also showing that alcohol is a limiting factor, especially in the context of intestinal inflammation. Red wine polyphenol extracts may have a role to potentiate SCFA-producers and to promote beneficial actions of probiotics through a host-microbe mutualism.35 36
We found a consistent association of plant protein intake with several fermentation pathways and the synthesis of anti-inflammatory nutrients and L-ornithine amino acid. Concordantly, a recent study in vegetarians showed an enrichment of pathways related to carbohydrate, amino acid, cofactor and vitamin metabolism.39 Animal models have demonstrated that nutrients produced by microbial metabolism of plant polysaccharides downregulate the expression of pro-inflammatory cytokines,40 suggesting an anti-inflammatory potential of plant-based diets through gut microbial metabolism.
We consistently observed inverse taxonomical associations of animal and plant foods across all cohorts. While animal protein intake was associated with lower Bifidobacterium abundance, the opposite direction was found for plant protein. A lower Bifidobacterium abundance has been observed in omnivores compared with vegans.15 30 Here, we replicate this link also in patients with IBD and IBS, in whom Bifidobacteria are generally depleted. An intervention with glycated pea protein in mice has been shown to increase Bifidobacterium and Lactobacillus abundance at the expense of Bacteroides fragilis and Clostridium perfringens,41 suggesting a specific role of plant protein besides other plant-derived nutrients to modulate the gut microbiome.
Animal protein dominated diets also tend to include higher amounts of saturated fats, which are impactful on the microbiome themselves.5 42 We here observed a positive association of the total fat intake and meat consumption with species that are dominant in the upper GI tract and oral cavity, while the opposite direction was found for plant-derived foods. Higher colonisation of these bacteria in the intestine has been reported in IBD, liver cirrhosis, colon cancer32 43 as well as several IMIDs such as arthritis and multiple sclerosis2 and has been linked to high-fat diets.30 Microbial carbohydrate fermentation normally creates a mildly acidic environment that inhibits overgrowth of these bacteria. A switch from a normal fat/carbohydrate ratio to a high-fat diet can impact the gut microbial composition and colonic pH. While there are many disease-related factors that influence the intestinal pH, our findings tentatively suggest that a high-fat omnivore diet affects the intestinal pH, further favouring colonisation of these bacteria in the intestine, as opposed to plant-dominated diets.
By contrast, fish showed consistent positive associations with Roseburia hominis and Faecalibacterium prausnitzii in our study. Fish is high in omega-3 polyunsaturated fatty acids (n-3 PUFA). Administration of n-3 PUFA in animal models has induced a decrease in pathobionts and pro-inflammatory metabolites and increased anti-inflammatory symbionts.5 Conversely, high-fat diets rich in n-6 PUFAs have depleted SCFA-producers and increased CRP levels in humans.42 These findings imply a role for optimised n-6/n-3 fatty acid ratios for gut microbiome targeted diets.
Finally, we have shown positive associations of fast food, processed meat, soft drinks and sugar with Fcal and the abundance of Clostridium bolteae, Ruminococcus obeum, Ruminococcus gnavus and Blautia hydrogenotrophica, Firmicutes that increase energy harvesting from the diet and are implicated in obesity and IMIDs.2 44 Functional studies consistently demonstrated an impact of food processing on the gut microbiome, leading to gut permeability and intestinal inflammation through an increase in mucolytic bacteria like Ruminococcus gnavus, Akkermansia muciniphila and Proteobacteria, production of endotoxins and induction of TH17 cells.5–11 Especially in the combination with a low fibre intake, these bacteria turn to the mucus layer, leading to an erosion of the gut barrier.5 A high consumption of sugar and soft drinks combined with a low vegetable intake has already been linked to IBD.7 We observed higher Fcal levels with the consumption of a high-sugar and high-fat diet, while the opposite was seen for plant-based foods. While this observation may not have a clinical benefit yet in the setting of IBS or HC with pre-clinical levels of Fcal, it implies a role for dietary strategies already at the public health level. Our findings suggest the gut microbiome as a link between diet and disease risk.
Our study has several limitations related to its cross-sectional nature and the complex interplay between diet and the gut microbiome. First, the cross-sectional nature of this study cannot identify causality in the observed associations. Second, while the use of whole shot-gun metagenomic sequencing allowed us to explore predicted metabolic profiles, further studies using faecal metabolomics and in vitro studies will be needed to confirm an increase or decrease in certain microbial functions and metabolites. Third, the time that is needed to elicit a lasting response of gut microbiota to dietary changes has not been well defined. Studies suggest that long-term habitual diet has a larger impact on a ‘core’ gut microbiome composition and function17–19 while short-term interventions have temporary effects.15 16 45 Longitudinal studies using high-resolution multi-omics data and dietary interventions with long-term follow-up will help us determine the time-dynamics of the gut microbiome in future, considering day-to-day variations and intestinal transit.29
Conclusions
Despite these limitations, we were able to derive dietary patterns that consistently correlate with groups of bacteria and functions known to infer mucosal protection and anti-inflammatory effects. We believe that the diet-microbiota associations that we described in this manuscript are robust: the results are consistent in the different cohorts and also remained significant after adjusting for additional cohort-specific factors such as medication usage. The findings suggest shared responses of the gut microbiota to the diet across patients with CD, UC, IBS and the general population that may be relevant to other disease contexts in which inflammation, gut microbial changes and nutrition are a common thread.3 46 A decrease in the here identified bacteria and their anti-inflammatory functions has already been identified in numerous inflammatory diseases, including cancer, atherosclerosis, obesity, non-alcoholic steatohepatitis, liver cirrhosis and IBD.3 4 22 32 Long-term diets enriched in legumes, vegetables, fruits and nuts; a higher intake of plant over animal foods with a preference for low-fat fermented dairy and fish; while avoiding strong alcoholic drinks, processed high-fat meat and soft drinks, have a potential to prevent intestinal inflammatory processes via the gut microbiome (table 2). Poor adherence to these principles has already been linked to an increased risk of IBD.33 47 48 We provide support for the idea that the diet can be a significant complementary therapeutic strategy through the modulation of the gut microbiome.3 49 For example, pre-clinical evidence shows that SCFA-producers such as Bifidobacterium species aid in invigorating a tumour specific T-cell response, raising the efficacy of cancer immunotherapy.50 It can be speculated that consumption of plant-based diets increases the abundance of these gut microbiota, further augmenting treatment responses.
Overview of diet-gut microbiome associations consistent across cohorts in this study and their pro-inflammatory or anti-inflammatory role
Data availability statement
All relevant data supporting the key findings of this study are available within the article and the supplementary files. Raw metagenomic sequencing reads and extended phenotypic data are available from the European Genome-phenome Archive data repository: 1000 IBD cohort [EGAD00001004194] and LifeLines Deep cohort [EGAD00001001991]. Codes used for generating the microbial profiles are publicly available at:[https://github.com/WeersmaLabIBD/Microbiome/blob/master/Protocol_metagenomic_pipeline.md]. All statistical analysis scripts are written in R and can be found here: https://github.com/WeersmaLabIBD/Microbiome/blob/master/Diet_Microbiome.md.
Ethics statements
Ethics approval
The study was approved by the Institutional Review Board (IRB) of the University Medical Center Groningen (1000IBD 2008.338; LifeLinesDEEP M12.113965).
Acknowledgments
The authors thank all the participants of the IBD and LifeLines-DEEP cohort for their contribution of samples and questionnaires. We thank BH Jansen for logistical and laboratory support as well as the IBD and research nurses at the UMCG for patient inclusion and collection of samples.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Footnotes
LAB and AVV are joint first authors.
Contributors RKW designed the study. LAB and AVV performed analyses. LAB wrote the manuscript. LAB, AVV, FI and RKW gathered during the study. RG and AK provided statistical advice. AVV, FI, VC, VP, CW, AK, MJECK, JF, GD, AZ and RKW were involved in data collection and preparation and critically reviewed the manuscript.
Funding LAB and RKW are supported by a research grant from the Seerave Foundation. RG and RKW are supported by the collaborative TIMID project (LSHM18057-SGF) financed by the PPP allowance made available by Top Sector Life Sciences & Health to Samenwerkende Gezondheidsfondsen (SGF) to stimulate public–private partnerships and co-financing by health foundations that are part of the SGF. JF, AZ and AK are supported by the Gravitation grant ExposomeNL from the from the Dutch Organization for Scientific Research (Nederlandse Organisatie voor Wetenschappelijk Onderzoek, NWO) (024.004.017). AZ is further supported by a European Research Council (ERC) Starting Grant (715772) and a NWO-VIDI grant (016.178.056). JF is supported by the NWO Gravitation Netherlands Organ-on-Chip Initiative (024.003.001) and the ERC Consolidator grant (101001678). JF and AZ are further supported by a grant from the Dutch Heart Foundation (CardioVasculair Onderzoek Nederland, CVON) (2018-27). CW is supported by the NWO Gravitation grant (024.003.001), a Spinoza award (NWO SPI 92-266) and a grant from the Netherlands’ Top Institute Food and Nutrition (GH001).
Competing interests RKW acted as consultant for Takeda and received unrestricted research grants from Takeda and Johnson and Johnson pharmaceuticals and speaker fees from AbbVie, MSD, Olympus and AstraZeneca. FI received a speaker fee from AbbVie. GD reports speakers’ fees from Janssen Pharmaceuticals, Takeda and Pfizer. MC received invited speaking fees from Takeda. No disclosures: All other authors have nothing to disclose.
Patient and public involvement Patients and/or the public were not involved in the design, or conduct, or reporting, or dissemination plans of this research.
Provenance and peer review Not commissioned; externally peer reviewed.