The Placental Microbiome Varies in Association with Low Birth Weight in Full-Term Neonates


Abstract
:1. Introduction
2. Experimental Section
2.1. Ethics Statement
2.2. Participants
2.3. Placental Samples
2.4. Anthropometric Measurement
2.5. Microbial Diversity Analysis
2.5.1. DNA Extraction and Polymerase Chain Reaction (PCR) Amplification
2.5.2. IlluminaMiSeq Sequencing and Bioinformatic Analysis
2.6. Statistical Analysis
3. Results
3.1. Clinical Characteristics and Anthropometric Measurements
3.2. Characteristics of Sequencing Results

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | N (Male/Female) | Maternal Age (Years) | Maternal BMI before Pregnancy | Gestational Age (Weeks) | Birth Weight (g) | Body Length (cm) | Ponderal Index (kg/m3) | Head Circumference (cm) | Placenta Weight (g) |
|---|---|---|---|---|---|---|---|---|---|
| NBW | 12 (6/6) | 31 ± 3 | 21.3 ± 2.2 | 39.1 ± 0.8 | 3452 ± 96 | 50.8 ± 1.1 | 26.2 ± 1.4 | 34.6 ± 0.8 | 701.2 ± 106.7 |
| LBW | 12 (6/6) | 31 ± 4 | 21.1 ± 2.9 | 38.9 ± 0.9 | 2623 ± 207 | 49.2 ± 4.6 | 23.2 ± 5.2 | 31.9 ± 1.6 | 492.8 ± 38.5 |
| p value | - | 0.76 | 0.82 | 0.58 | <0.01 ** | 0.34 | 0.02 * | 0.013 * | <0.01 ** |
| NBW | LBW | p Value | |
|---|---|---|---|
| Sequences | 17,600 ± 3693 | 16,150 ± 3768 | 0.49 |
| OTUs | 49 ± 4.18 | 48.44 ± 7.89 | 0.026 * |
| Chao | 57 ± 7.19 | 54.44 ± 13.33 | 0.044 * |
| Shannon | 0.96 ± 0.12 | 1.06 ± 0.36 | 0.083 |
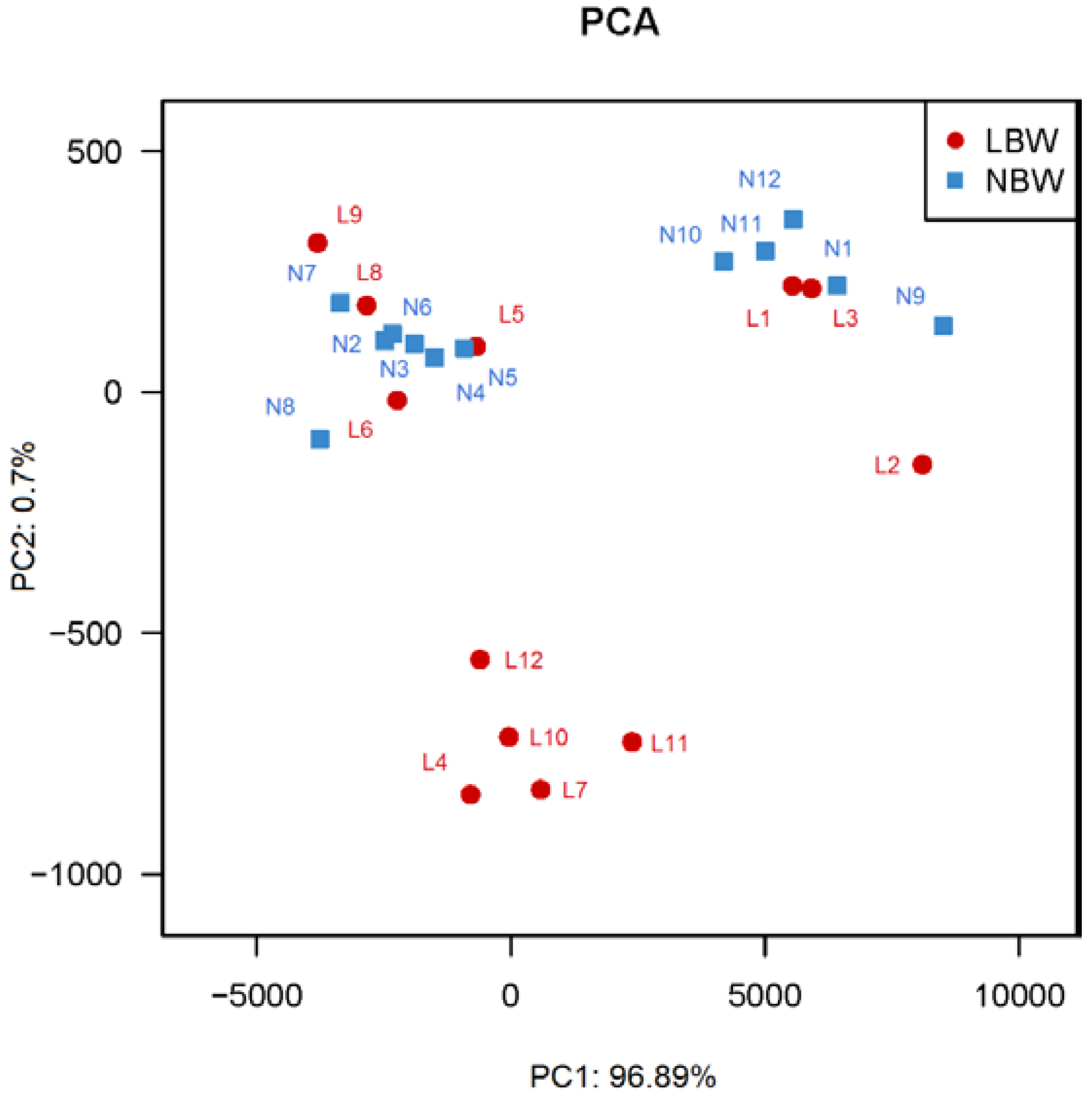
3.3. Principal Coordinates Analysis (PCA) between LBW and NBW Groups

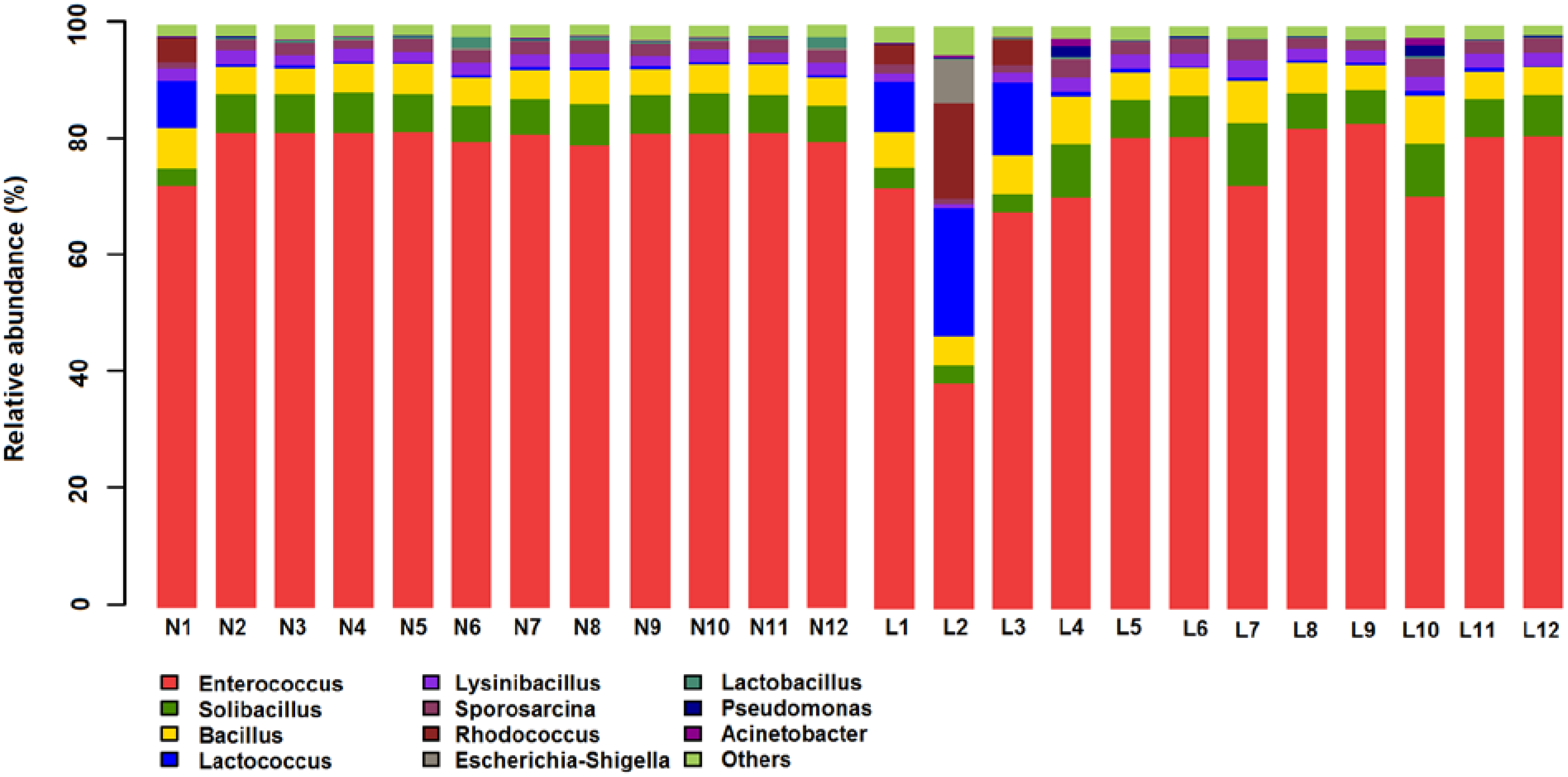
3.4. Microbial Structures of the Placenta in LBW and NBW Groups



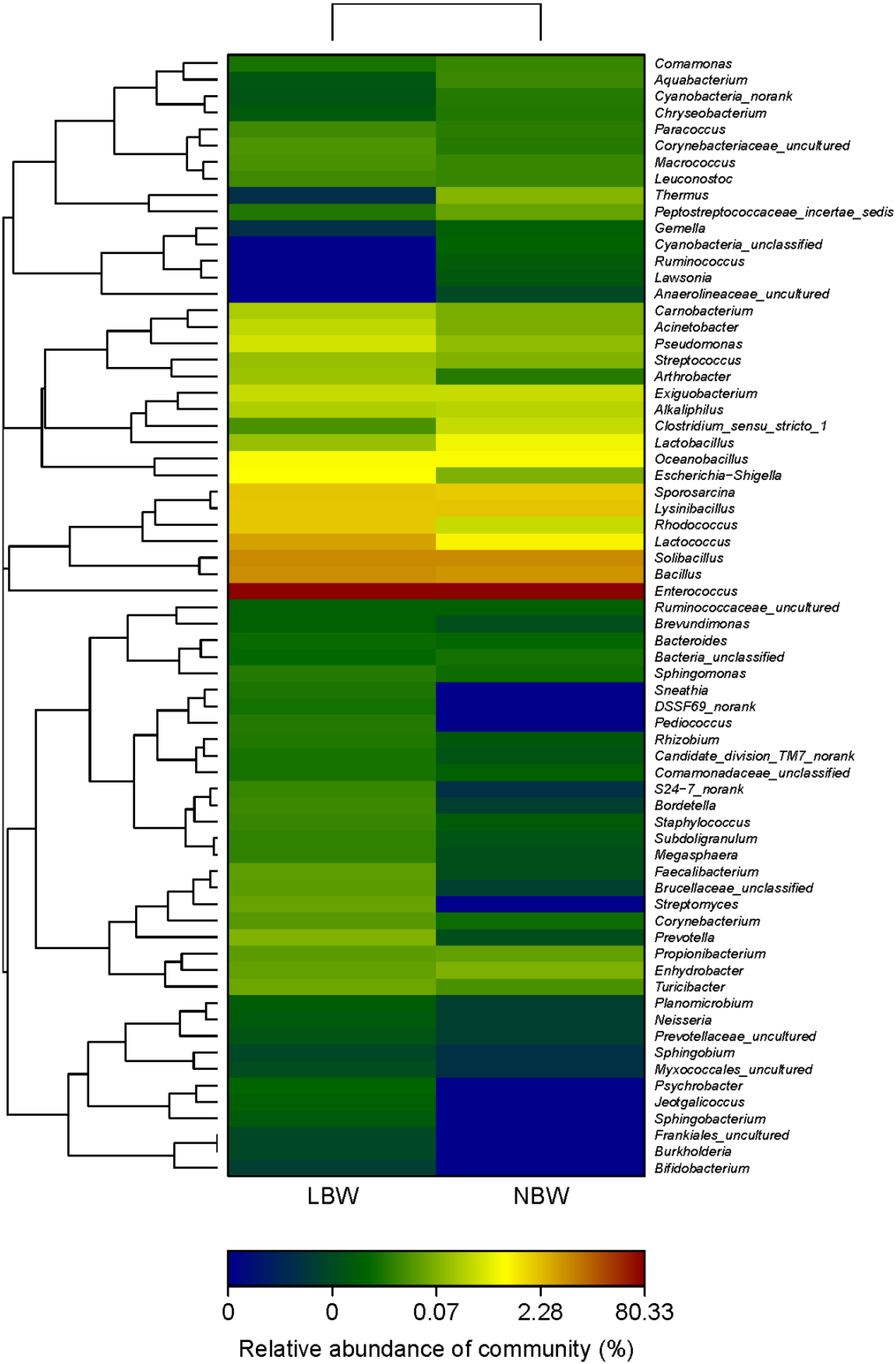
3.5. Phylotypes Significantly Different between LBW and NBW Groups
| Taxonomic Rank | NBW (%) | LBW (%) | * p Value | |
|---|---|---|---|---|
| phylum | Fusobacteria | 0.000 | 0.011 | 0.001 |
| phylum | Cyanobacteria | 0.020 | 0.003 | 0.008 |
| genus | Cyanobacteria_unclassified | 0.007 | 0.000 | 0.001 |
| genus | DSSF69_norank | 0.000 | 0.013 | 0.001 |
| genus | Jeotgalicoccus | 0.000 | 0.005 | 0.001 |
| genus | Pediococcus | 0.000 | 0.017 | 0.001 |
| genus | Sneathia | 0.000 | 0.011 | 0.001 |
| genus | Clostridium_sensu_stricto_1 | 0.253 | 0.026 | 0.002 |
| genus | Lactobacillus | 0.531 | 0.115 | 0.005 |
| genus | Ruminococcus | 0.004 | 0.000 | 0.007 |
| genus | Megasphaera | 0.003 | 0.016 | 0.008 |
| genus | Faecalibacterium | 0.002 | 0.038 | 0.014 |
| genus | Lawsonia | 0.004 | 0.000 | 0.014 |
| genus | Sphingobacterium | 0.000 | 0.007 | 0.016 |
| genus | Cyanobacteria_norank | 0.014 | 0.003 | 0.047 |
3.6. Variation in the Placental Microbiome Associated with Birth Weight

4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hocher, B.; Slowinski, T.; Bauer, C.; Halle, H. The advanced fetal programming hypothesis. Nephrol. Dial. Transpl. 2001, 16, 1298–1299. [Google Scholar] [CrossRef]
- Hales, C.N.; Barker, D.J.P.; Clark, P.M.S.; Cox, L.J.; Fall, C.; Osmond, C.; Winter, P.D. Fetal and Infant Growth and Impaired Glucose-Tolerance at Age 64. Br. Med. J. 1991, 303, 1019–1022. [Google Scholar] [CrossRef] [PubMed]
- Poulsen, P.; Vaag, A. The intrauterine environment as reflected by birth size and twin and zygosity status influences insulin action and intracellular glucose metabolism in an age- or time-dependent manner. Diabetes 2006, 55, 1819–1825. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Zhang, Z.X.; Li, W.H.; Feng, K.; Sun, Q.; Cohen, H.J.; Xu, T.; Wang, H.; Liu, A.M.; Gong, X.M.; et al. Low birth weight is associated with components of the metabolic syndrome. Metab. Clin. Exp. 2010, 59, 1282–1286. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Zhang, Z.X.; Cohen, H.J.; Wang, H.; Li, W.; Wang, T.; Xu, T.; Liu, A.; Gai, M.Y.; Ying, S.; et al. Evidence of a relationship between infant birth weight and later diabetes and impaired glucose regulation in a Chinese population. Diabetes Care 2008, 31, 483–487. [Google Scholar] [CrossRef] [PubMed]
- Nelson, K.E.; Weinstock, G.M.; Highlander, S.K.; Worley, K.C.; Creasy, H.H.; Wortman, J.R.; Rusch, D.B.; Mitreva, M.; Sodergren, E.; Chinwalla, A.T.; et al. A catalog of reference genomes from the human microbiome. Science 2010, 328, 994–999. [Google Scholar] [PubMed]
- Consortium, H.M.P. Structure, function and diversity of the healthy human microbiome. Nature 2012, 486, 207–214. [Google Scholar] [Green Version]
- Qin, J.; Li, Y.; Cai, Z.; Li, S.; Zhu, J.; Zhang, F.; Liang, S.; Zhang, W.; Guan, Y.; Shen, D.; et al. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature 2012, 490, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ottman, N.; Smidt, H.; de Vos, W.M.; Belzer, C. The function of our microbiota: Who is out there and what do they do? Front. Cell. Infect. Microbiol. 2012, 2, 104. [Google Scholar] [CrossRef] [PubMed]
- Watson, E.D.; Cross, J.C. Development of structures and transport functions in the mouse placenta. Physiology (Bethesda, Md.) 2005, 20, 180–193. [Google Scholar] [CrossRef] [PubMed]
- Wassenaar, T.M.; Panigrahi, P. Is a foetus developing in a sterile environment? Lett. Appl. Microbiol. 2014, 59, 572–579. [Google Scholar] [CrossRef] [PubMed]
- Aagaard, K.; Ma, J.; Antony, K.M.; Ganu, R.; Petrosino, J.; Versalovic, J. The placenta harbors a unique microbiome. Sci. Transl. Med. 2014. [Google Scholar] [CrossRef] [PubMed]
- Doyle, R.M.; Alber, D.G.; Jones, H.E.; Harris, K.; Fitzgerald, F.; Peebles, D.; Klein, N. Term and preterm labour are associated with distinct microbial community structures in placental membranes which are independent of mode of delivery. Placenta 2014, 35, 1099–1101. [Google Scholar] [CrossRef] [PubMed]
- Antony, K.M.; Ma, J.; Mitchell, K.B.; Racusin, D.A.; Versalovic, J.; Aagaard, K. The preterm placental microbiome varies in association with excess maternal gestational weight gain. Am. J. Obst. Gynecol. 2015, 212, 653.e651–653.e616. [Google Scholar] [CrossRef] [PubMed]
- Mercuro, G.; Bassareo, P.P.; Flore, G.; Fanos, V.; Dentamaro, I.; Scicchitano, P.; Laforgia, N.; Ciccone, M.M. Prematurity and low weight at birth as new conditions predisposing to an increased cardiovascular risk. Eur. J. Prev. Cardiol. 2013, 20, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Xiao, X.; Zhang, Q.; Mao, L.; Li, M.; Yu, M.; Xu, J.; Wang, Y. Correlation of high-molecular-weight adiponectin and leptin concentrations with anthropometric parameters and insulin sensitivity in newborns. Int. J. Endocrinol. 2014, 2014, 435376. [Google Scholar] [CrossRef] [PubMed]
- UPARSE. Available online: http://drive5.com/uparse/ (accessed on 15 June 2015).
- RDP. Available online: http://rdp.cme.msu.edu/ (accessed on 15 June 2015).
- Amato, K.R.; Yeoman, C.J.; Kent, A.; Righini, N.; Carbonero, F.; Estrada, A.; Gaskins, H.R.; Stumpf, R.M.; Yildirim, S.; Torralba, M.; et al. Habitat degradation impacts black howler monkey (Alouatta pigra) gastrointestinal microbiomes. ISME J. 2013, 7, 1344–1353. [Google Scholar] [CrossRef] [PubMed]
- White, J.R.; Nagarajan, N.; Pop, M. Statistical methods for detecting differentially abundant features in clinical metagenomic samples. PLoS Comput. Biol. 2009, 5, e1000352. [Google Scholar] [CrossRef] [PubMed]
- Gangarapu, V.; Yildiz, K.; Ince, A.T.; Baysal, B. Role of gut microbiota: Obesity and NAFLD. Turkish J. Gastroenterol. 2014, 25, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishna, B.S. Role of the gut microbiota in human nutrition and metabolism. J. Gastroenterol. Hepatol. 2013, 28 (Suppl. 4), 9–17. [Google Scholar] [CrossRef] [PubMed]
- Le Chatelier, E.; Nielsen, T.; Qin, J.; Prifti, E.; Hildebrand, F.; Falony, G.; Almeida, M.; Arumugam, M.; Batto, J.M.; Kennedy, S.; et al. Richness of human gut microbiome correlates with metabolic markers. Nature 2013, 500, 541–546. [Google Scholar] [CrossRef] [PubMed]
- Ejtahed, H.S.; Mohtadi-Nia, J.; Homayouni-Rad, A.; Niafar, M.; Asghari-Jafarabadi, M.; Mofid, V. Probiotic yogurt improves antioxidant status in type 2 diabetic patients. Nutrition 2012, 28, 539–543. [Google Scholar] [CrossRef] [PubMed]
- Naito, E.; Yoshida, Y.; Makino, K.; Kounoshi, Y.; Kunihiro, S.; Takahashi, R.; Matsuzaki, T.; Miyazaki, K.; Ishikawa, F. Beneficial effect of oral administration of Lactobacillus casei strain Shirota on insulin resistance in diet-induced obesity mice. J. Appl. Microbiol. 2011, 110, 650–657. [Google Scholar] [CrossRef] [PubMed]
- Satokari, R.; Gronroos, T.; Laitinen, K.; Salminen, S.; Isolauri, E. Bifidobacterium and Lactobacillus DNA in the human placenta. Lett. Appl. Microbiol. 2009, 48, 8–12. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, J.; Xiao, X.; Zhang, Q.; Mao, L.; Yu, M.; Xu, J. The Placental Microbiome Varies in Association with Low Birth Weight in Full-Term Neonates. Nutrients 2015, 7, 6924-6937. https://doi.org/10.3390/nu7085315
Zheng J, Xiao X, Zhang Q, Mao L, Yu M, Xu J. The Placental Microbiome Varies in Association with Low Birth Weight in Full-Term Neonates. Nutrients. 2015; 7(8):6924-6937. https://doi.org/10.3390/nu7085315
Chicago/Turabian StyleZheng, Jia, Xinhua Xiao, Qian Zhang, Lili Mao, Miao Yu, and Jianping Xu. 2015. "The Placental Microbiome Varies in Association with Low Birth Weight in Full-Term Neonates" Nutrients 7, no. 8: 6924-6937. https://doi.org/10.3390/nu7085315