


Abstract
Methyl-n-pentylnitrosamine (MPN) is carcinogenic for the rat esophagus. To determine organ specificity for MPN activation by human tissues, microsomes isolated from human organs (snap-frozen <6 h after death or removed surgically) were incubated with [pentyl-3H]MPN, and [3H]pentaldehyde formation was measured by high-pressure liquid chromatography of its 2,4-dinitrophenylhydrazone using radioflow assay. With 100 μM MPN, mean depentylation rates were 6.6 (liver), 2.9 to 3.8 (kidney, stomach, small intestine, and colon), and 0.4 to 1.6 (esophagus, lung, and skin) pmol of pentaldehyde/mg of protein/min. Of 14 human esophagi, four showed relatively high depentylation rates of 3.3 to 4.1 pmol/mg/min. Apparent Km was 80 to 160 μM (Vmax, 3–15 pmol/mg/min) for three esophagi, 90 to 130 (2 livers), and 1330 (1 kidney) μM. Rat tissues showed mean depentylation rates for 100 μM MPN of 24.9 (liver), 14.5 (esophagus), 7.0 (lung), and 0.0 to 2.7 (5 other tissues) pmol/mg/min. MPN depentylation by rat cytochrome P450 2A3 showed an apparentKm of 8 μM (Vmax, 70 pmol/nmol of P450/min) and was competitively inhibited by the CYP2A inhibitor coumarin (apparentKi, 4 μM). Coumarin (0.4 mM) inhibited microsomal depentylation of 100 μM MPN by 37 to 62% for human esophagus, liver, kidney, and colon and for rat esophagus but not for rat liver and lung. MPN depentylation by rat esophageal microsomes increased up to 90% on adding P450 reductase. The results indicate organ-specific MPN metabolism by rat but not human esophagus. Nevertheless, the relatively high activity of four human esophagi might indicate increased susceptibility of some individuals to carcinogenesis by unsymmetrical dialkylnitrosamines.
There are especially high rates of squamous cell carcinoma of the esophagus in South Africa, Iran, and China, and the incidence of esophageal adenocarcinomas is increasing rapidly in Western countries (Mirvish, 1995). Nitrosamines are probably initiating agents for esophageal cancer, especially squamous cell carcinoma, for several reasons (Magee, 1989; Mirvish, 1995). Esophageal cancer is induced in rats by nitrosamines, such as methyl-n-pentylnitrosamine (MPN2) (Fig. 1), methylbenzylnitrosamine (MBZN), N-nitrosopiperidine, and the tobacco-specific compound N′-nitrosonornicotine (NNN), but not by dimethylnitrosamine (DMN) (Preussmann and Stewart, 1984). NNN and related nitrosamines that occur in tobacco smoke are the probable initiators of esophageal cancer in smokers (Hecht and Hoffmann, 1989;Magee, 1989). Asymmetric dialkylnitrosamines similar to MPN that are derived from moldy corn could contribute to the etiology of esophageal cancer in China and South Africa (Mirvish, 1995). The corn moldFusarium moniliforme was reported to convertiso-pentylamine toN-methyl-N-iso-pentylamine, which is readily nitrosated to yield iso-MPN; however, we failed to confirm this result (Mirvish, 1995). Esophageal exposure to nitrosamines could occur by direct absorption while foods are being swallowed (Haorah et al., 1999). Subjects from Chinese counties with high incidences of esophageal cancer showed elevated urinary excretions of N-nitrosoproline, an indicator of in vivo nitrosation (Wu et al., 1993). A case-control study in Linxian found significant associations of the c1/c1 genotype of the cytochrome P450 isozyme CYP2E1 with esophageal hyperplasia and cancer (Tan et al., 2000). Possibly, the c1/c1 phenotype of CYP2E1 metabolizes nitrosamines less efficiently in the liver than does the c2/c2 phenotype, hence allowing more nitrosamine to reach the esophagus.

Metabolism of MPN to yield formaldehyde and a pentylating agent (route a), PENT and a methylating agent (route b), and stable hydroxy-MPNs (route c).
Nitrosamines are activated by hydroxylation on the carbon atom(s) adjacent to the nitrosamine group. In the rat esophagus, this reactions is catalyzed by one or more unidentified cytochrome P450s (Murphy and Spina, 1994; Chen et al., 1999a; Gopalakrishnan et al., 1999). MPN metabolism by depentylation to give pentaldehyde (PENT) and an agent that methylates DNA bases (route b of Fig. 1) is a more likely activation mechanism than route a of Fig. 1, in which MPN is demethylated to give formaldehyde and a pentylating agent, because 7- and O6-methylguanine, but not 7- andO6-pentylguanine, were detected in esophageal DNA of rats treated with MPN (Huang et al., 1993). Rat liver microsomes produced formaldehyde, PENT, 4-hydroxy-MPN, and nitrite from 6 mM MPN (Ji et al., 1989). Human esophageal and liver microsomes metabolized 6 mM MPN to yield formaldehyde and PENT (Huang et al., 1992). When [3H]PENT production from [pentyl-3H]MPN was assayed; theKm values for MPN depentylation by rat esophageal and liver microsomes, rat CYP2E1, human CYP2E1, and human CYP2A6 [a liver, nasal, and lung P450 (Su et al., 2000)] were 64, 610, 210, 115, and 17 μM, respectively (Chen et al., 1999a). MPN metabolism by rat and human esophageal microsomes was strongly inhibited by CO, a P450 inhibitor (Huang et al., 1992; Chen et al., 1999a).
In the present study, we performed further experiments on depentylation by human and rat microsomes of (in standard runs) 100 μM3H-labeled MPN of high specific activity (Chen et al., 1999a). Even if human esophagus actively depentylated MPN, this would indicate that nitrosamines might preferentially induce human esophageal cancer only if this activity was high relative to that for other organs. Therefore, we compared the depentylating activity of human esophageal microsomes with that of microsomes prepared from other human organs. For similar reasons, we also determined the MPN depentylation activity of microsomes from six rat organs, in addition to the previously studied (Chen et al., 1999a) esophagus and liver.
The unusually low Km of 17 μM for MPN depentylation by CYP2A6 (Chen et al., 1999a) suggested that a CYP2A enzyme was responsible for MPN metabolism by rat esophagus. CYP2A3, a rat ortholog of human CYP2A6, occurs mainly in nasal mucosa and, to minor extents, in the lung and esophagus (Su et al., 1996;Gopalakrishnan et al., 1999) and catalyzed MBZN activation with an apparent Km of only 0.6 μM (Von Weymarn et al., 1999). Hence, CYP2A3 might also activate MPN in the rat esophagus and nasal cavity where MPN induces tumors (Bulay and Mirvish, 1979). Accordingly, we also investigated MPN depentylation by CYP2A3 (Liu et al., 1996). Finally, we examined the effect of adding P450 reductase on MPN metabolism by rat esophageal microsomes.
Materials and Methods
Synthesis of [3H]MPN.
1-Bromo-2-pentene or 5-bromo-1-pentene was reacted with methylamine to give methyl-2-pentenylamine or methyl-4-pentenylamine, respectively (Chen et al., 1999a). Hydrogenation with tritium at Moravek Biochemicals (Brea, CA) yielded [2,3-pentyl-3H]- or [4,5-pentyl-3H]methylpentylamine. As required, about 100 mCi of the amine was nitrosated to give [2,3-pentyl-3H]MPN or [4,5-pentyl-3H]MPN (Chen et al., 1999a). The [3H]MPN was purified by thin layer chromatography on alumina developed with hexane-ether-acetic acid (40:60:5) and, on the day of the experiment, by reaction with semicarbazide to remove PENT, followed by thin layer chromatography on alumina developed with hexane-ether (4:6) (Chen et al., 1999a). For final MPN concentrations of <50 μM, the contribution of [3H]MPN to the total MPN was significant and was determined by gas chromatography-thermal energy analysis of the [3H]MPN. Specific activity of the [3H]MPN was typically 160 μCi/μmol.
Tissues Studied.
See Acknowledgments for the suppliers of the human tissue specimens. Each specimen weighed 5 to 50 g. All tissues appeared normal and did not include obvious tumors. The position of esophageal specimens within the esophagus was not reported. Most tissues were obtained from autopsies, all of which were performed ≤6 h after death. At autopsy, the tissues were snap-frozen in liquid N2 and shipped in dry ice. Other tissues were obtained during operations and handled similarly. Tables1 and 2 summarize the pathology reports. Rat esophagi of freshly killed adult male Sprague-Dawley rats were obtained from Harlan Bioproducts for Science (Indianapolis, IN), who stripped away the outer connective tissue, flash-froze the tissues in liquid N2, and mailed them in dry ice (Chen et al., 1999a). Other rat tissues were obtained from male Sprague-Dawley rats 6 to 8 weeks old (Sasco, Omaha, NE) that were euthanized with CO2. Tissues were stored at −70°C, most for <6 months, before use.
Depentylation of 100 μM MPN by microsomes from human tissues
Kinetic rate constants for MPN depentylation by human and rat microsomes
Isolation of Microsomes.
From human tissues
Each microsome sample was obtained from a different donor. All operations were performed in a cold room at 5°C. The stored tissues were frozen in liquid N2 without prior thawing, wrapped in cheese cloth, and pulverized with a hammer. Portions (5–10 g) of softer tissues (liver, kidney, and lung) were dispersed with a Tissumizer (Tekmar-Dohrmann, Mason, OH) in 0.1 M potassium phosphate buffer (pH 7.4, 3 ml/g of tissue) containing 0.1 mM dithiothreitol and 0.1 mM phenylmethylsulfonyl fluoride (buffer A) and further homogenized with a motor-driven Potter-Elvejhem tissue grinder (Labglass, Vineland, NJ) fitted with a Teflon piston (Chen et al., 1999a). For harder tissues (esophagus, stomach, intestines, and skin), 5 to 10 g pieces of frozen tissue were ground to a fine powder with a mortar and pestle precooled with liquid N2. The powder from human esophagus was homogenized in a chilled 15-ml Tenbroeck Pyrex-glass tissue homogenizer (Scientific Products, Edison, NJ) in 3 ml/g of tissue of 0.1 M sodium pyrophosphate buffer (pH 7.4) containing 1 mM EDTA, 1 mM dithiothreitol, and 5 mM phenylmethylsulfonyl fluoride (buffer B) (Murphy and Spina, 1994; Chen et al., 1999a). The homogenizing medium for all other human tissues was buffer A (3 ml/g of tissue). Microsome fractions were isolated by differential centrifugation (Huang et al., 1992), resuspended in buffer A or B containing 20% glycerol, and analyzed for protein by the Lowry method (Lowry et al. 1951). The suspensions were stored at −70°C, most for <3 months, before use. The activity of several samples that were re-examined was mostly unaffected after storage for 2 years.
From rat tissues.
Microsomes from 16 to 18 rat esophagi at a time were prepared as before (Murphy and Spina, 1994; Chen et al., 1999a). In brief, esophagi frozen in liquid N2 were crushed in a Bessman tissue pulverizer (Fisher Scientific, Springfield, NJ) and gently homogenized in buffer B (1 ml/esophagus) using a Tenbroeck homogenizer. Microsomes from other rat tissues were prepared as described for the corresponding human tissues.
Metabolic Experiments.
These followed methods used before (Huang et al., 1992; Chen et al., 1999a). Incubations were performed in 5-ml stoppered disposable polystyrene round-bottom tubes with 12 to 16 tubes per experiment. In standard runs, we mixed 100 mM potassium phosphate buffer (pH 8) with 10 mM MgCl2 and 5 mM semicarbazide HCl (the pH was then 7.4). We then added 10 to 30 μl of a microsomal suspension containing 100 to 200 μg of protein, MPN (20 to 2000 μM, 2–10 μCi/tube) and, finally, an NADPH-generating system (2 mM NADP, 10 mM glucose 6-phosphate, and 2 units of glucose-6-phosphate dehydrogenase; final concentrations are indicated in all cases). The total volume was 0.5 ml, and the final pH was 7.4. The mixtures were incubated for 20 min at 37°C with shaking twice per second [reaction rates are constant over this time (Chen et al., 1999a)]. Appropriate control tubes were included. Duplicate tubes were used for each condition. Reactions were stopped by adding Ba(OH)2 and ZnSO4, and PENT 2,4-dinitrophenylhydrazone was prepared and separated by high-pressure liquid chromatography on an analytical C18 column developed with a gradient rising from 50 to 70% ethanol in water (Chen et al., 1999a). The PENT hydrazone showed a retention time of 21 min (Fig.2).
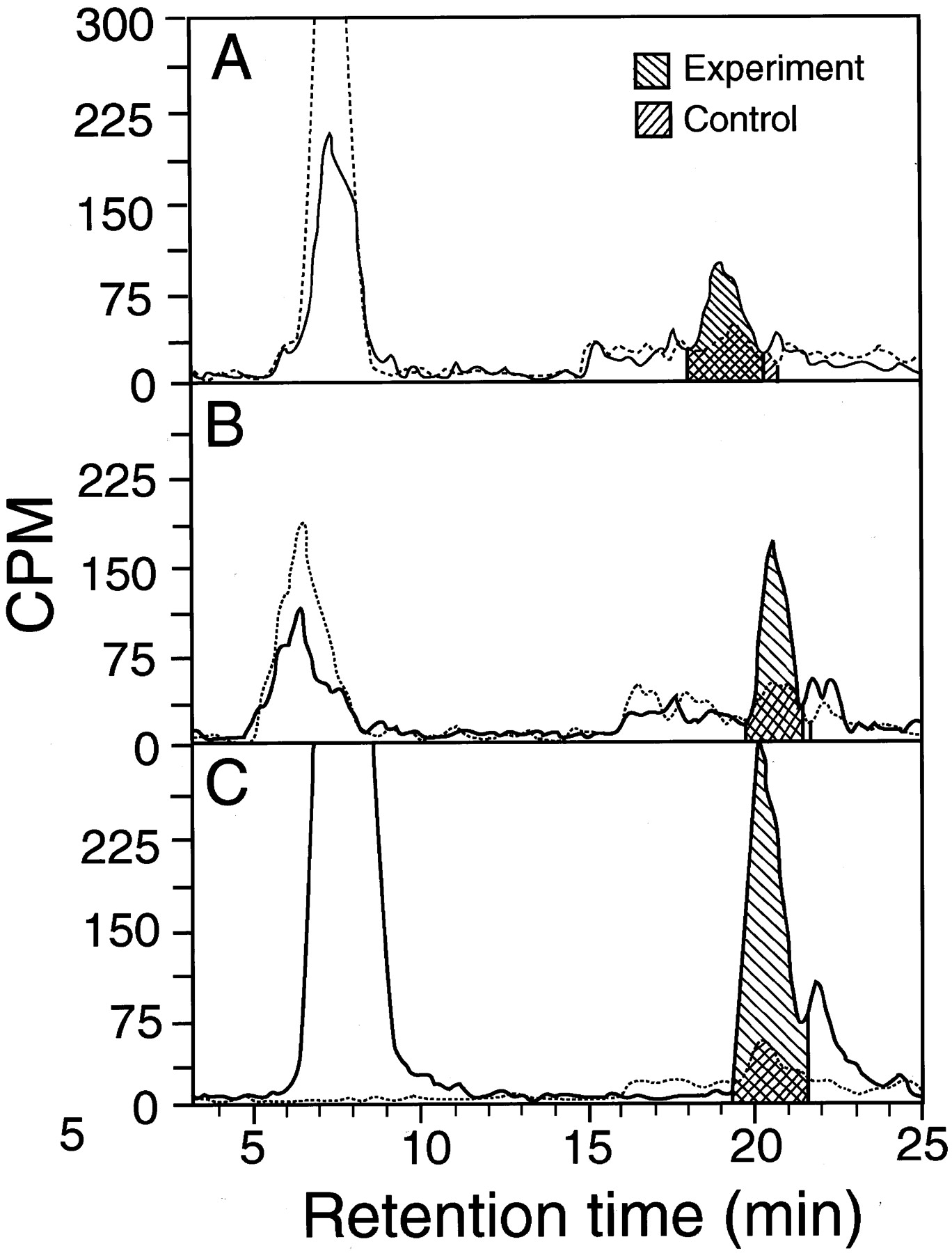
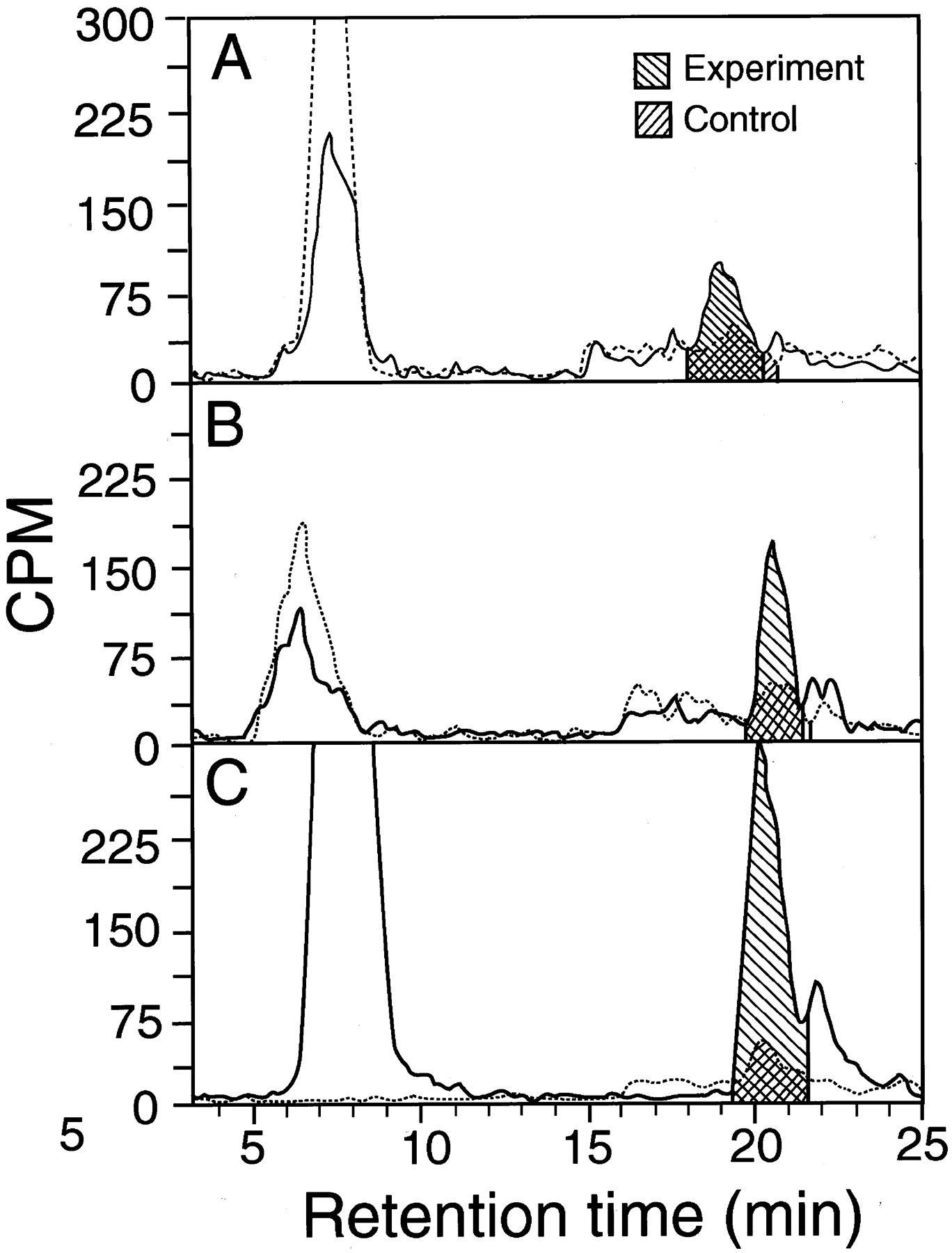
Tracings of high-pressure liquid chromatography runs in which [3H]PENT yield was determined after 100 μM [3H]MPN was incubated with and without microsomes.
The peak at 7 to 8 min is unchanged [3H]MPN, and that at 19 to 21 min is the 2,4-dinitrophenylhydrazone of [3H]PENT. The recovery of [3H]MPN was variable, presumably due to its volatility. A, metabolism by human esophageal microsomes (sample 2 of Table 2; 100 μg of protein/tube). B, metabolism by rat esophageal microsomes (50 μg of protein/tube). C, metabolism by rat liver microsomes (50 μg of protein/tube). ——, results in the presence of microsomes. · · ·, results in the absence of microsomes. Shadings show areas used to calculate [3H]PENT yields.
We collected 1-ml fractions and counted them in a Beckman scintillation counter (Beckman Coulter, Inc., Fullerton, CA) (Chen et al., 1999a) or counted the eluate directly with a flow scintillation analyzer (500 TR Series, Packard, Meriden, CT) with integration of peak areas by the Flo-one program with “force horizontal” baseline (Fig.2). Peak width of the PENT hydrazone was about 3 min. Counts per minute for blanks without microsomes were subtracted from experimental values. The results, expressed as counts per minute, were used to calculate PENT yield from MPN. The yield at 20 min was expressed as rate per minute, assuming that the rate did not change for 20 min, as found in a test with phenobarbital-induced rat liver microsomes (Chen et al., 1999a). Yields were graphed with a Prism program (GraphPad Software, San Diego, CA) for determining nonlinear regression curves.Km and Vmaxwere obtained from the best-fitting straight lines for Lineweaver-Burke plots using Excel 4.0 (Microsoft, Redmond, WA).
Results
Comments on Methods.
Most of these studies were carried out using [2,3-pentyl-3H]MPN, but some of the earlier tests used [4,5-pentyl-3H]MPN. As explained previously (Chen et al., 1999a), the yield of [3H]PENT should not depend on which isomer is employed because MPN activation does not involve compounds with labile hydrogen at C-2 to C-5 and because the product PENT was not subjected to alkaline conditions and hence should not have enolized. The two isomers seemed to give similar results, but the3H 2,3-pentyl isomer of MPN seemed to be more readily synthesized and more stable than the 3H 4,5-pentyl isomer. The efficiency of the conversion of PENT to its hydrazone is 23% (Chen et al., 1999a). As before (Chen et al., 1999a), the results were not corrected for this loss.
MPN Metabolism by Human Microsomes.
Table 1 shows the depentylation rates when human microsomes were incubated with 100 μM MPN. This MPN concentration was used in standard runs even though it is lower than some of theKm values for human esophagus and liver (Table 2) and hence did not give accurate comparisons of the maximum rates. A standard MPN concentration of 100 μM was used for three reasons. 1) The use of low MPN levels brought the MPN level closer to those of nitrosamines that people are exposed to. 2) In studies on microsomes, if we had raised the standard MPN level to 300 μM, for example, we would probably have recruited P450 isozymes that metabolize MPN with relatively high Km values. In fact, this probably occurred in our previous study with 6 mM unlabeled MPN (Huang et al., 1992). 3) If we had raised the standard MPN concentration, we would have obtained a reduced percent yield of PENT because absolute PENT yield would not have been much higher than that obtained with 100 μM. However, we would have had to use [3H]MPN of a higher specific activity to obtain reasonable counts for the PENT hydrazone peak, and this would have raised the background counts. In fact, this was somewhat of a problem when high-MPN concentrations were used in the measurements ofKm.
Mean depentylation activity for human tissues was highest for the liver, moderate for kidney, small intestine, stomach, and colon, low for esophagus (24% of that for the liver), and especially low for lung and skin. Esophageal microsomes from 14 subjects were tested (Table3). The results for different samples of the same batch of microsomes showed reasonable agreement with most standard errors <30% of the mean values. In contrast, the mean activity for esophageal microsomes from different subjects varied from 0 to 4.1 pmol of PENT/mg of protein/min. Of the 14 esophagi examined, six were apparently normal tissue from individuals with cancer. The only noted relationship to the cause of death was that none of the cancer cases showed esophageal activity ≥1.8 pmol of PENT/mg/min.
Depentylation of 100 μM MPN by 14 samples of human esophageal microsomes
Table 2 shows the kinetic rate constants (apparentKm and Vmax) and metabolic rates for the depentylation of [3H]MPN by human microsomes from three esophagi, two livers, and one kidney. These results were obtained using 20 to 2000 μM MPN and are compared in Table 2 with our published values (Chen et al., 1999a) for rat esophagus and liver. Lineweaver-Burk plots were constructed (e.g., see Fig.3). The Kmvalues for microsomes from human esophagus and liver were 80 to 160 μM and were 1.25 to 2.5 times the Km for rat esophageal microsomes. The Vmax for human esophageal microsomes was 17 to 75% of that for rat esophageal microsomes. Coumarin (0.4 mM) inhibited the depentylation of 100 μM MPN by 37 to 62% in tests on microsomes from human esophagus, liver, kidney, and colon (Table 4). As before (Chen et al., 1999a), coumarin was incubated with the microsomes for 15 min before the addition of MPN. The effect of coumarin on human lung microsomes could not be examined because of their low activity (Table1).

Kinetics of [3H]MPN depentylation by microsomes (100 μg of protein/tube) from human esophagus no. 1 (Table 3).
This figure shows a Lineweaver-Burk plot of 1/S versus 1/V, where S = substrate concentration and V = rate of reaction. Each point gives the results for an individual tube. This figure indicates aKm of 80 μM and aVmax of 15 pmol/mg of protein/min.
Effect of 0.4 mM coumarin on the depentylation of 100 μM MPN by human and rat microsomes
MPN Metabolism by Rat Microsomes and CYP2A3.
Table 5 shows depentylation rates when 100 μM MPN was incubated with microsomes from eight rat tissues. The average depentylation rate of 14.5 pmol/mg of protein/min for esophagus was exceeded only by that for liver, the order of activity being liver > esophagus > lung > colon > forestomach > glandular stomach > kidney > small intestine. Coumarin inhibited the depentylation of 100 μM MPN by 55% when rat esophageal microsomes were tested but had no effect with rat liver and lung microsomes (Table 4).
Rate of depentylation of 100 μM MPN by microsomes from rat tissues
We examined MPN depentylation by rat CYP2A3 expressed in Sf-9 insect cells using a baculovirus (Liu et al., 1996). The CYP2A3 was incubated with a 4-fold molar excess of rabbit P450 reductase because this ratio produced maximum testosterone metabolism by this P450 (Liu et al., 1996). Under these conditions, the Km was 8 μM, and Vmax was 70 pmol/nmol of P450/min (Fig. 4). Coumarin inhibition of MPN depentylation by rat CYP2A3 was then investigated. The results were expressed as Dixon and Cornish-Bowden plots, i.e., plots of 1/V, and S/V, respectively, against inhibitor (coumarin) concentration, where S = substrate concentration and V = rate of reaction (Fig.5). The MPN concentration where the two lines met in the Dixon plot indicated an apparentKi of 4 μM, and the nearly parallel lines in the Cornish-Bowden plot indicated that the inhibition was competitive (Cornish-Bowden, 1974).

Kinetics of MPN depentylation by rat CYP2A3 (60 pmol/tube) with added P450 reductase.
See the legend to Fig. 3 for explanation of this figure. Microsomes from Sf-9 cells containing CYP2A3 (Liu et al., 1996) were used. P450 reductase was purified from liver microsomes of phenobarbital-treated rabbits (French and Coon, 1979) and had a specific activity of 45 to 60 μmol of cytochrome c that is reduced/mg of protein/min. CYP2A3 (60 pmol in 29 μl of 50 mM Tris-acetate buffer, pH 7.4, containing 1 mM EDTA and 20% glycerol) and P450 reductase (230 pmol in 29 μl of 10 mM phosphate buffer, pH 7.4, containing 0.1 mM EDTA and 10% glycerol) were gently mixed at intervals for 30 min at 0°C. The standard incubation components were then added, and depentylation was assayed as described under Materials and Methods. CYP2A3 or P450 reductase were omitted in control tubes. This figure indicates values of 8 μM forKm and 70 pmol/nmol of P450/min forVmax.
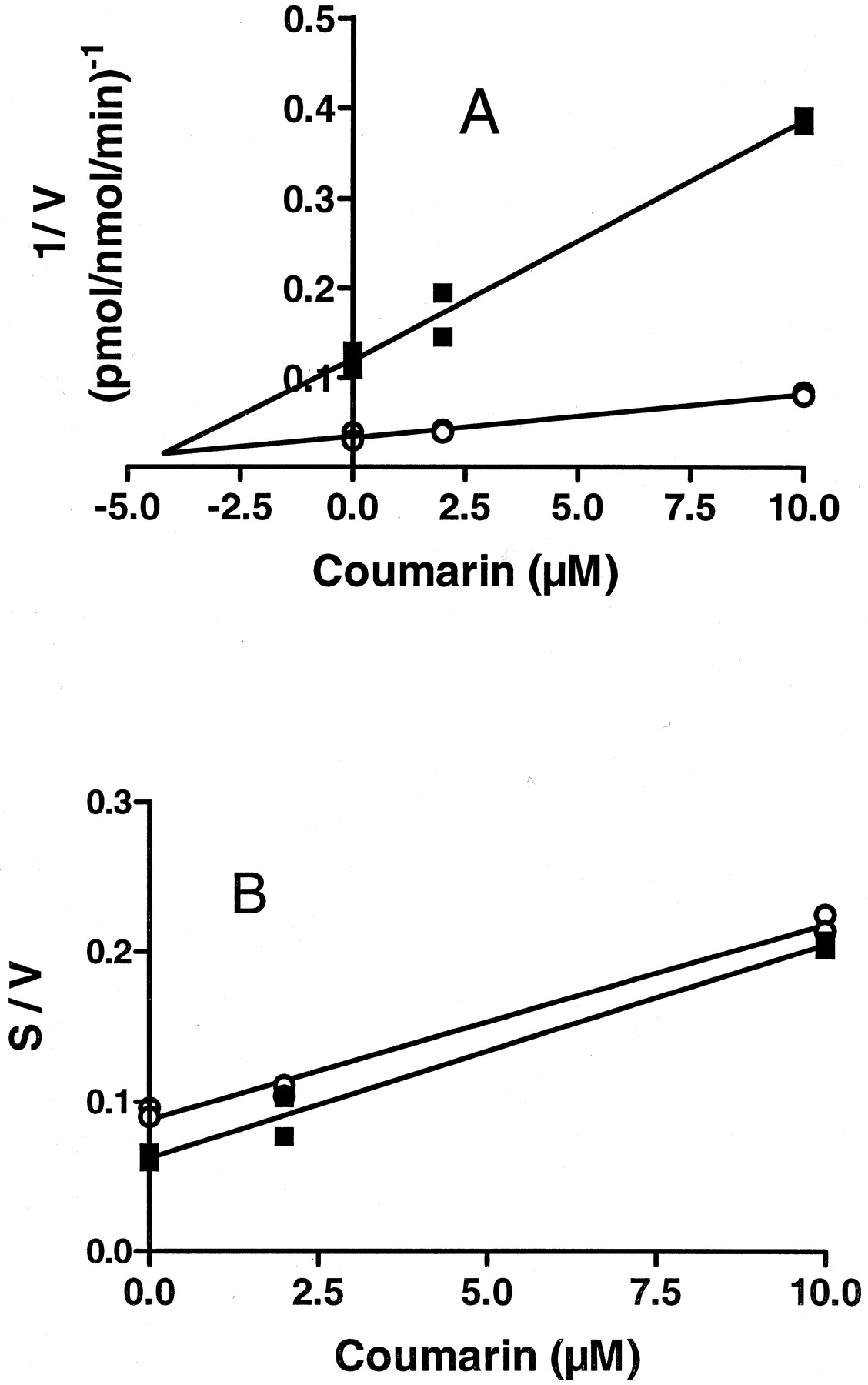
Coumarin inhibition of MPN metabolism by rat CYP2A3.
A, Dixon plot (1/V versus inhibitor concentration). B, Cornish-Bowden plot (S/V versus inhibitor concentration; S = substrate concentration,V = rate of reaction). Coumarin (2 and 10 μM final concentrations) was preincubated with CYP2A3 in phosphate buffer containing MgCl2 and semicarbazide for 15 min at room temperature. Control tubes were preincubated similarly but without coumarin. [3H]MPN and the NADPH-generating system were then added, and depentylation was assayed as described underMaterials and Methods and in footnote aof Table 4. This figure indicates a Ki of 4 μM. ▪, tests with 0.53 μM MPN; ○, tests with 2.65 μM MPN.
Finally, we determined the effect of adding varied amounts of P450 reductase on the depentylation of 100 μM MPN by rat esophageal and liver microsomes. Addition of the reductase increased PENT yield from 100 μM MPN by up to 90% (esophagus) and 54% (liver) (Fig.6). In the case of esophagus, 80 pmol of reductase increased depentylation by only 15%, whereas 160 pmol of reductase increased the yield by 74%. Incubation of MPN with 300 pmol of reductase in the absence of P450 did not produce a significant yield of PENT.
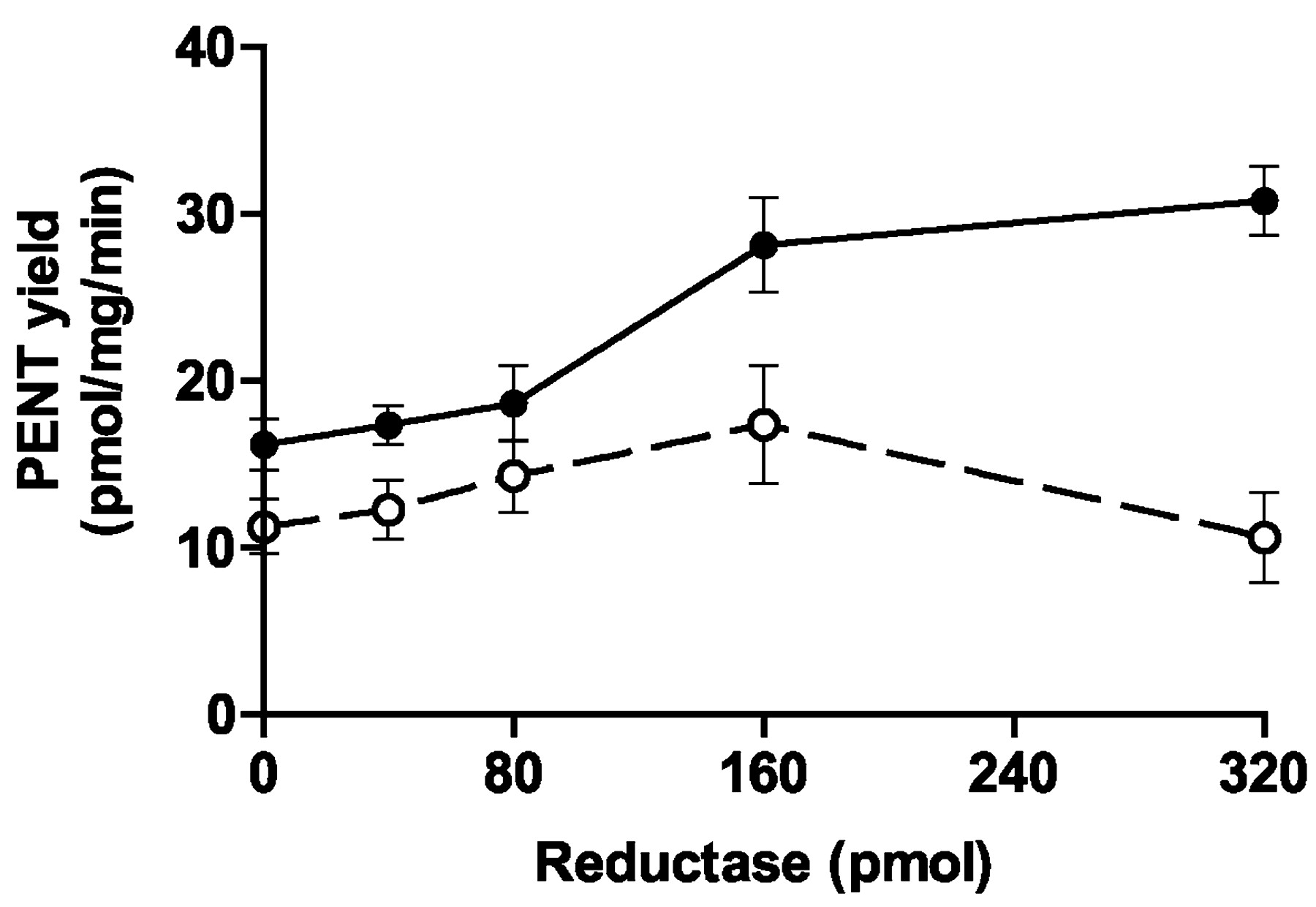
Effect of P450 reductase on MPN depentylation by rat esophageal and liver microsomes.
The microsomes (50 μg of protein/12–14 μl of suspension) were preincubated for 30 min on ice with 0 to 320 pmol of P450 reductase (see Fig. 4) in 0 to 40 μl of suspension, phosphate buffer (270–300 μl) containing MgCl2 and semicarbazide was added, the mixture was incubated with [3H]MPN (final concentration, 100 μM) and NADPH-generating system (total volume, 500 μl), and depentylation was determined (see Materials and Methods). Means and (as vertical bars) standard deviations of the results for four tubes per condition are shown. ●, esophageal microsomes; ○, liver microsomes.
Discussion
MPN Metabolism in Human Tissues.
Although human esophageal microsomes generally exhibited low activity for MPN dealkylation, microsomes from 4 of the 14 esophagi showed relatively high activities of 3.3 to 4.1 pmol of PENT/mg of protein/min for the metabolism of 100 μM MPN (Table 3), about half the mean value of 6.6 pmol/mg/min for human liver (Table 1). If this high activity occurred only in mucosal microsomes [likely because nitrosamines are chiefly activated in rat esophagus by basal mucosal cells (Koenigsmann et al., 1988)], the activity of the mucosal microsomes may have been much higher than the values reported here because the mucosa is only 15 to 20% of the total human esophagus, all of which was used to prepare the microsomes. The Km andVmax values for human esophageal microsomes were similar to those for human liver microsomes and had somewhat higher Km values for human than for rat esophageal microsomes (Table 2). The relative activity of two enzyme preparations at low-substrate concentrations is indicated by the relativeVmax/Km ratios, which should equal the relative activities at a single, low-substrate concentration. In agreement with this concept,Vmax/Km for esophagus no. 1 divided by the same ratio for esophagus no. 10 was 6.8 (Table 2), and the depentylation rate for 100 μM MPN in esophagus no. 1 divided by that for esophagus no. 10 (Table 3) showed a fairly similar value of 9.1. Other comparisons ofVmax/Km for the different microsome samples also showed reasonable agreements with comparisons of the rates for 100 μM MPN. The unusually highVmax of 74 pmol/mg/min for human kidney (Km, 1330 μM) might have been due to the high-MPN level used with this tissue and, hence, the recruitment of additional P450s not involved at low-MPN concentrations.
With regard to the P450s known to occur in human esophagus, evidence was found for CYP1A1/1A2 in all tested human esophagi and for CYP2B6 in 12 of 25 esophagi (Murray et al., 1994; Nimura et al., 1997). In human esophagus, Lechevrel et al. (1999) detected the mRNAs of P450s 1A1, 1A2, 2A, 2E1, 3A5, 4A1, and 4B1 and expression of P450s 1A1/1A2, 2A6, 2E1, 3A4/5, and 4A. Barrett's esophagus showed expression of P450s 1A2, 2E1, 2C9/10, and 3A4 (Hughes et al., 1999). All of 22 samples of human esophageal microsomes metabolized NNN and DMN (Smith et al., 1998). NNN activation was reduced 20 to 30% by a CYP3A inhibitor. Coumarin 7-hydroxylase activity (characteristic of some CYP2A enzymes) was not detected. It was concluded that the esophageal metabolism of NNN and DMN was catalyzed by CYP3A4 and CYP2E1, respectively (Smith et al., 1998). MBZN is metabolized in human liver chiefly by CYP2A6 and CYP2E1 (Morse et al., 1999). DMN, diethylnitrosamine, and methylphenylnitrosamine were dealkylated by CYP2A13, a human lung enzyme (Su et al., 2000). Our finding that 0.4 mM coumarin inhibited MPN metabolism by 37 to 62% in the human esophagus, liver, kidney, and colon (Tables 3 and 4) indicates that these tissues may contain CYP2A enzymes.
Our results with human tissues, which were frozen <6 h after death, could have been affected by postmortem changes (e.g., freezing human liver samples, thawing them, and then keeping them at room temperature for 5 h led to 90% losses of the P450s) (Yamazaki et al., 1997). Here, frozen tissue samples were thawed at 6°C and immediately homogenized and centrifuged to obtain the microsomes. Therefore, most of the P450 activity in the frozen human tissues probably persisted in the microsomes. Also, the microsomes were stored in 20% (2.2 M) glycerol to increase their stability. Glycerol inhibits DMN metabolism by rat CYP2E1 with a Ki of 53 mM (Yoo et al., 1987) and, hence, might have inhibited MPN metabolism. However, this effect is known to occur only with CYP2E1, which probably is not involved in MPN metabolism by rat esophagus. No differences were found when MPN depentylation by rat esophageal microsomes and phenobarbital-induced liver microsomes, suspended in 20% glycerol (concentration in incubation medium, 40–130 mM), was compared with MPN metabolism in glycerol-free medium (Chen et al., 1999a).
Comparison with MPN Metabolism in Rat Tissues and by the Rat P450, CYP2A3.
Rat esophageal microsomes showed apparentKm values of 64, 3.8 to 5.1, and 49 μM for the activation of MPN, MBZN, and NNN, respectively (Murphy et al., 1990; Murphy and Spina, 1994; Chen et al., 1999a; Von Weymarn et al., 1999). The activity of these microsomes for MPN metabolism was higher than that of microsomes from six other rat organs and was exceeded only by those from the liver (Table 5). This supports the view that these nitrosamines induce tumors of the rat esophagus because they are preferentially activated there. The main reason why these nitrosamines do not induce liver cancer despite extensive metabolism in that organ may be the slow rate of cell turnover in the liver [basal cells of the mouse esophageal mucosa have half-lives of 4.5 days (LeBlond et al., 1964)]. The 10-fold higher Km for MPN compared with that for MBZN is consistent with the approximately 10 times lower carcinogenicity of MPN for the rat esophagus (Haorah et al., 1999). Nevertheless, MPN is nearly as specific an esophageal carcinogen as MBZN (Preussmann and Stewart, 1984).
MPN metabolism by rat esophageal microsomes was inhibited up to 65% by coumarin (Chen et al., 1999a). Most of this inhibition occurred only when coumarin was preincubated with the microsomes for 15 to 30 min before the incubation with MPN, which was also found for MBZN metabolism by these microsomes (Chen et al., 1999a; Von Weymarn et al., 1999). These results support the view (Von Weymarn et al., 1999) that an active metabolite of coumarin causes the inhibition. Coumarin inhibited the metabolism of other nitrosamines by rat esophageal and nasal microsomes (the latter are rich in CYP2A3) (Bereziat et al., 1995; Patten et al., 1998; Von Weymarn et al., 1999), the mouse liver enzyme, CYP2A5 (Negishi et al., 1989), and the human enzyme CYP2A6, where coumarin showed a Ki of 7.5 μM for the inhibition of MPN depentylation (Chen et al., 1999a). The hypothesis (Gopalakrishnan et al., 1999) that the rat esophageal P450 that activates esophagus-specific nitrosamines is a CYP2A isozyme is supported by these observations and by findings that CYP2A3 metabolized MPN and MBZN with Km values of only 8 and 0.63 μM, respectively (Fig. 4; Von Weymarn et al., 1999). However, the P450 in rat esophageal microsomes is not CYP2A3 because these microsomes contain only low levels of this P450 (Gopalakrishnan et al., 1999) and, unlike CYP2A3, are poor catalysts of coumarin 7-hydroxylation (Chen et al., 1999a; Gopalakrishnan et al., 1999). Our findings that CYP2A3 metabolism of MPN showed aKm of 8 μM (Fig. 4) and was competitively inhibited by coumarin with a Ki of 4 μM (Fig. 5) extends the list of nitrosamines metabolized by CYP2A3 and the list of CYP2A enzymes (which now includes P450s 2A3, 2A5 and 2A6) that are inhibited by low levels of coumarin.
Our observation that added P450 reductase increased the rate of MPN metabolism by up to 90% in the rat esophagus and 54% in the rat liver (Fig. 6) suggests that the level of this enzyme limits nitrosamine metabolism in these tissues. Similar findings were reported for warfarin metabolism by rat liver microsomes (Kaminsky and Guengerich, 1985). The activities of P450s in microsomes are far lower than those of the same levels of purified P450s incubated with excess reductase, probably because different P450s in microsomes compete for a limited amount of reductase (Kaminsky and Guengerich, 1985).
General Comments.
The highly sensitive assay of [3H]MPN depentylation used here and previously (Chen et al., 1999a) offers a useful method for studying the metabolism of certain CYP2A isozymes and of CYP2E1 in small samples of microsomes or of the individual P450s. Our results using 100 μM MPN resemble our preliminary results for four human esophagi using 6 mM MPN (Huang et al., 1992) in that the mean rate of MPN metabolism for human esophagus in both studies was 9 to 11% of that for rat esophagus and was <25% of that for human liver. Human esophagus did not generally show a high level of MPN metabolism compared with other human organs. The relatively high MPN metabolism in 4 of 14 human esophagi could be due to differences in tissue handling or to intrinsically high-P450 activity, which could increase the susceptibility to esophageal carcinogenesis by unsymmetrical dialkylnitrosamines.
In human esophagus, CYP2E1 may be involved in the metabolism of nitrosamines, especially DMN, because this P450 exhibited polymorphism associated with esophageal cancer (Tan et al., 2000), and rat and human CYP2E1 showed apparent Km values of 20 to 22 μM (Yoo et al., 1990; Patten et al., 1992) for DMN demethylation, and 210 and 115 μM, respectively, for MPN depentylation (Patten et al., 1992; Chen et al., 1999a). Human esophageal microsomes activated DMN (Smith et al., 1998), Therefore, if nitrosamines cause human esophageal cancer in high-incidence areas of China and South Africa where cigarette smoking is probably not a factor (Mirvish, 1995), then symmetrical dialkylnitrosamines might be at least as important in the etiology as unsymmetrical dialkylnitrosamines, such as MPN and MBZN. In contrast, unsymmetrical dialkylnitrosamines are the most potent carcinogens in the rat esophagus (Preussmann and Stewart, 1984). However, the tobacco-specific cyclic nitrosamine NNN is metabolized by both human and rat esophageal microsomes (Murphy and Spina, 1994; Smith et al., 1998), is carcinogenic for the rat esophagus (Preussmann and Stewart, 1984), and is probably involved in the etiology of esophageal cancer in smokers (Hecht and Hoffmann, 1989). The finding that rat esophagus showed high activity for MPN metabolism relative to that for most other rat organs seems to explain why MPN and, perhaps, certain other nitrosamines induce esophageal cancer in rats and is attributed to an unidentified esophagus-specific P450. This might be a homolog of CYP2A13, a human lung P450 that demethylated methylphenylnitrosamine, a rat esophageal carcinogen (Su et al., 2000), or might be CYP2B21, a novel P450 expressed in rat esophagus (Brookman-Anissah et al., 2001).
Acknowledgments
We thank Lawrence Schopfer (Eppley Institute) for advice about enzyme kinetics, Peter Guengerich (Vanderbilt University, Nashville, TN) for advice about the effects of P450 reductase, and the National Disease Research Interchange (Philadelphia, PA) and the Western Division of the Cooperative Human Tissue Network (Department of Pathology, Case Western Reserve University, Cleveland, OH) for supplying the human tissue samples.
Footnotes
-
↵1 Parts of this study were presented at two meetings (Chen and Mirvish, 1996; Chen et al., 1999b).
-
This research was supported by Grant RO1-CA-35628 and core Grant P30-CA-36727 from the National Cancer Institute, Grant RO1-ES-07462 from the National Institute for Environmental Health Sciences, and Grant 97B-125 from the American Institute of Cancer Research.
- Abbreviations used are::
- MPN
- methyl-n-pentylnitrosamine
- MBZN
- methylbenzylnitrosamine
- NNN
- N′-nitrosonornicotine
- DMN
- dimethylnitrosamine
- PENT
- pentaldehyde
- P450
- cytochrome P450
- Received March 15, 2001.
- Accepted June 6, 2001.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}