Article Text

Abstract
Background: “Probiotic” bacteria are effective in treating some inflammatory bowel diseases. However which bacteria confer benefit and mechanisms of action remain poorly defined. Dendritic cells, which are pivotal in early bacterial recognition, tolerance induction, and shaping of T cell responses, may be central in mediating the effects of these bacteria.
Aims: To assess effects of different probiotic bacteria on dendritic cell function.
Methods: Human intestinal lamina propria mononuclear cells, whole blood, or an enriched blood dendritic cell population were cultured with cell wall components of the eight bacterial strains in the probiotic preparation VSL#3 (four lactobacilli, three bifidobacteria, and one streptococcal strains). Dendritic cells were identified and changes in dendritic cell maturation/costimulatory markers and cytokine production in response to probiotic bacteria were analysed by multicolour flow cytometry, in addition to subsequent effects on T cell polarisation.
Results: VSL#3 was a potent inducer of IL-10 by dendritic cells from blood and intestinal tissue, and inhibited generation of Th1 cells. Individual strains within VSL#3 displayed distinct immunomodulatory effects on dendritic cells; the most marked anti-inflammatory effects were produced by bifidobacteria strains which upregulated IL-10 production by dendritic cells, decreased expression of the costimulatory molecule CD80, and decreased interferon-γ production by T cells. VSL#3 diminished proinflammatory effects of LPS by decreasing LPS induced production of IL-12 while maintaining IL-10 production.
Conclusions: Probiotic bacteria differ in their immunomodulatory activity and influence polarisation of immune responses at the earliest stage of antigen presentation by dendritic cells.
- DC, dendritic cell
- ELISA, enzyme linked immunosorbent assay
- IBD, inflammatory bowel diseases
- LPDC, lamina propria DC
- LPS, lipopolysaccharide
- PBMC, peripheral blood mononuclear cell
- PE, phycoerythrin
- TLR, toll-like receptor
- dendritic cell
- mucosal immunology
- probiotic bacteria
- cytokines
Statistics from Altmetric.com
- DC, dendritic cell
- ELISA, enzyme linked immunosorbent assay
- IBD, inflammatory bowel diseases
- LPDC, lamina propria DC
- LPS, lipopolysaccharide
- PBMC, peripheral blood mononuclear cell
- PE, phycoerythrin
- TLR, toll-like receptor
The intestinal immune system forms the largest part of the immune system. It interacts with a complex antigenic load in the form of food antigens, commensal bacteria, and occasional pathogens. Dendritic cells (DC) are pivotal in earliest bacterial recognition and in shaping T cell responses. In the intestine DC have specialised functions most likely as a result of their close proximity with the external environment.1 They are involved in generating regulatory T cells and IgA producing B cells through production of cytokines such as IL-10 and TGFβ.2–4 They contribute to oral tolerance induction5 and determine homing of lymphocytes back into intestinal tissue.6 Intestinal DC interact directly with luminal bacteria by passing their dendrites between epithelial tight junctions into the gut lumen7 and indirectly with bacteria that have gained access via M cells.
Microbial products play a key role in regulating DC responses. DC interact with microbes using pattern recognition receptors, including toll-like receptors (TLR). Using such a system, DC distinguish between and mount different responses to even closely related organisms.8–11 For example, lipopolysaccharide (LPS) from E coli interacts with TLR4 and generates a Th1 response, whereas LPS from Porphyromonas gingivalis interacts with TLR2 and generates a Th2 response.11,12 This ability to respond appropriately to different microbial products is of particular importance in the gastrointestinal tract, where a diverse microbial flora lies close to DC.
Dendritic cells sense antigen in tissues before migrating to draining lymph nodes, where they have the unique ability to activate and influence functional differentiation of naïve T cells. Signals from DC can determine whether tolerance or an active immune response occurs to a particular antigen13,14 and furthermore influence whether a Th1 or Th2 immune response predominates. DC subtype, whether CD11c+ (myeloid) or CD11c- (plasmacytoid/lymphoid), maturation status, and cytokine production contribute to the type of T cell response. For example, DC upregulate the costimulatory molecules, CD80 and CD86, and produce IL-12 which contributes to a Th1 response.15 DC also produce IL-10 and IL-4 which promote the generation of a Th2 response or regulatory T cells.3,16
Some commensal organisms from intestinal flora have health-promoting properties and are used in probiotic preparations. The probiotic mixture, VSL#3 (VSL#3 Pharmaceuticals, Fort Lauderdale, FL, USA), which contains eight different bacterial strains (four lactobacilli, three bifidobacteria, and one Streptococcus thermophilus subsp Salivarius) is beneficial in treating some inflammatory bowel diseases (IBD). Controlled clinical trials have shown that VSL#3 maintains remission of pouchitis17 and prevents onset of pouchitis after pouch formation.18 The non-pathogenic E coli Nissle strain is beneficial in maintenance treatment of ulcerative colitis.19,20
Central to the mechanism of action of probiotics is their modulation of host immune response. An increased understanding of the effects of clinically active probiotic bacteria on the immune system may enable refinement of probiotic treatments for use under defined disease circumstances and enhance our knowledge of immune homeostasis in the specialised environment of the intestine. We hypothesised that probiotic organisms exert modulatory activity on the host immune system via DC. The objective was to investigate effects of combined and individual probiotic strains and LPS on the phenotype and function of both blood and intestinal DC derived from the colonic lamina propria in order to establish which organisms may confer benefits at this early stage in antigen presentation. Cross regulatory effects between microbial products inducing polarised responses were also assessed. Furthermore, the subsequent effect of DC exposed to probiotic bacteria on polarisation of the T cell response was examined.
METHODS
Preparation of bacterial fractions
The bacterial fractions investigated are shown in table 1. Bacterial fractions were prepared as described in Lammers, et al.21 The VSL#3 Lactobacillus and Bifidobacterium strains were grown anaerobically at 37°C in MRS medium (Difco, USA) supplemented with 0.05% L-cysteine. The VSL#3 Streptococcus salivarius subsp thermophilus was grown anaerobically in M17 medium (Difco, USA). Streptococcus faecium was also grown anaerobically in M17 medium. Non-pathogenic Escherichia coli Nissle strain 1917 was grown aerobically in LB (Difco, USA). Mid log cultures were collected by centrifugation and subsequently sonicated (Branson Sonifier W-250, Heinemann, Schwäbisch, Germany) at a power level 5−6 at 30% duty for five minutes. Sonicates were centrifuged and bacterial debris fractions were collected. In assays where all strains were assessed in combination, sonicates of individual strains were pooled.
Bacterial strains
Monoclonal antibodies
The antibodies used were: CD11c-FITC (KB90) (Dako, Cambridgeshire, UK); CD3-PC5 (UCHT-1), CD14-PC5 (MIP9), CD16-PC5 (B73.1), CD19-PC5 (4G7), CD56-PC5 (N901), and CD8-PC5 (B9.11) (Beckman Coulter, High Wycombe, UK); CD34-CyChrome (581), CD86-PE (BU63), CD40-PE (LOB7/6) (Serotec, Oxford, UK); HLA-DR-APC (G46-6), CD80-PE (L307.4), CD83-PE (HB15e), CD3-FITC (UCHT1), CD69-PE (FN50), and CD8-FITC/PE/APC (SK1) (BD Biosciences Pharmingen, Oxford, UK). Isotype matched controls were obtained from the same manufacturers. Intracellular cytokine staining used: IFN-γ-PE (clone D9D10), IFN-γ-FITC (clone B-B1), IL-10-PE (clone JES3-9D7) (Serotec, Oxford, UK), and IL-12(p40)-PE (clone C11.5) and IL-4 PE (3007.11) (BD Biosciences Pharmingen, UK). Neutralising anti-IL-10 antibody (Clone 9D7) was purchased from Endogen (Cheshire, UK).
Blood dendritic cells
Blood was collected from healthy volunteers into lithium heparin and incubated with culture medium alone (RPMI-1640 Dutch Modification, Sigma-Aldrich, UK), varying concentrations of bacterial cell debris fractions (106–109 CFU/ml), LPS (1 μg/ml) or control medium alone for four or 16 hours at 37°C in 5% CO2. An enriched DC population was prepared from peripheral blood mononuclear cells (PBMC). PBMC were prepared by centrifugation of blood over a Ficoll-Paque gradient (Amersham Biosciences, UK) and cultured overnight in complete medium (RPMI-1640 Dutch Modification supplemented with 10% FCS, 2 mM L-glutamine, 100 μg/ml streptomycin, 100 U/ml penicillin) at 37°C in 5% CO2. Non-adherent DC rich low density cells were separated on hypertonic metrizamide (14.5% w/v) by centrifugation and incubated at 37°C in 5% CO2 for four hours with bacterial cell debris fractions, LPS or control medium alone.
Human colonic lamina propria dendritic cells
Human intestinal biopsies were obtained at routine colonoscopy from patients with macroscopically and histologically normal intestine who had been referred with rectal bleeding or altered bowel habit. Informed consent was obtained and the protocol was approved by the local ethics committee. At least 10 mucosal biopsies (approximately 60 mg) were taken per patient.22 Collagenase digestion of tissue was performed using collagenase D (Roche Diagnostics, Mannheim, Germany) in 10 ml RPMI-1640 HEPES (Sigma Aldrich Co Ltd, Irvine, UK), 2% FCS and 2 μM DNase I (Boehringer, Mannheim, Germany). Lamina propria mononuclear cells (2.5×105 cells per well, Falcon 96 well, U bottom) were cultured for four hours at 37°C in 5% CO2 with bacterial cell debris fractions, LPS, or control medium alone.
Cytokine production by DC
Cytokine production by DC subsets was assessed by intracellular staining in whole blood cultures, enriched DC cultures, and intestinal cell cultures in a modification of the method described by North et al.23 Comparison of paired cultures, one incubated with monensin (3 μmol/l) to maintain cytokine within the cells and the other incubated without monensin allowed assessment of cytokine production. In whole blood assays, red cells were lysed with 500 μl Optilyse C (Beckham Coulter, High Wycombe, UK) for 15 minutes at room temperature. Cells were fixed with Leucoperm A (100 μl, Serotec) and permeabilised with Leucoperm B (100 μl, Serotec). The cytokine antibody (5 μl) was added for 30 minutes. Blocking experiments were performed with an unlabelled antibody of an identical clone to the one used for cytokine staining and an irrelevant antibody to assess specificity of binding.
Enzyme linked immunosorbent assay (ELISA)
Concentrations of IL-10 in culture supernatants from enriched DC populations incubated with bacterial fractions were measured by ELISA using a paired antibody kit (Endogen, UK). IL-10 coating antibody was used at 2 μg/ml and biotinylated antihuman IL-10 detecting antibody at 0.75 μg/ml. Absorbance values were read at 450 nm on an ELISA plate reader. The assay’s detection limit was 36 pg IL-10/ml.
Differentiation of T cells after exposure to bacterial products
Two methods were used to assess effects of exposure to bacterial products on the subsequent T cell response. Naïve CD4+ T cells were prepared from PBMC by negative selection to a purity >87% using a StemSep kit (StemCell Technologies Inc, Vancouver, Canada). DC rich low density cells (8×103 cells per well) exposed to control medium alone, VSL#3 (108 CFU/ml) or LPS (1 μg/ml) were co-cultured with 4×105 per well allogeneic naïve CD4+ T cells for six days. At this time point, cultures were supplemented with 30 U/ml recombinant IL-2. After a further six days, cells were cultured for four hours in complete medium with phorbol-myrsitate-acetate (PMA-16.2 nmol/l), ionomycin (2 μmol/l), and monensin (3 μmol/l) or monensin alone. Cytokine production was assessed by intracellular staining.23
To assess whether prior exposure to bacterial products influences IFN-γ production by polyclonally activated T cells, blood was incubated with varying concentrations of bacterial fractions in culture medium for 14 hours at 37°C in 5% CO2. To assess the role of IL-10, a neutralising anti-IL-10 antibody or an isotype matched control antibody was added to some cultures (20 μg/ml). Subsequently, cultures were activated with phorbol-myrsitate-acetate (PMA) and ionomycin, in addition to monensin, or treated with monensin alone for a further four hours at 37°C in 5% CO2. Cytokine production was assessed by intracellular staining.
Data and statistical analysis
Data were acquired uncompensated using a FACSCalibur (Becton Dickinson) flow cytometer. Objective compensation was applied using the Compensation Toolbox within the WinList Version 5.0 flow cytometry software (Verity, Topsham, ME USA). The proportion of cytokine positive cells was determined by subtraction of histograms using the super-enhanced Dmax (SED) normalised subtraction facility in WinList. Staining of cells cultured in the absence of monensin was subtracted from staining of cells cultured in the presence of monensin, giving a measure of ongoing cytokine production.24 This technique allows positive cells to be resolved in situations where distribution histograms overlap. The use of the same antibody to label cells from both monensin treated and untreated cultures gives this technique a high degree of sensitivity for detecting small changes in antibody binding. Specificity of antibody labelling was confirmed in competition experiments with unlabelled antibodies. Absolute cell counts were obtained by reference to counts of Flow-Count fluorospheres acquired simultaneously. Two tailed t tests were used to compare proportions and absolute numbers of cells. Data were paired where appropriate. Values of p<0.05 were regarded as significant.
RESULTS
Identification of dendritic cells
Blood DC were identified as HLA-DR+ lineage negative (CD3−, CD14−, CD16−, CD19−, CD34−, CD56−) cells. Within this gate, CD11c+ (myeloid) and CD11c− (plasmacytoid) DC populations were present. Cytokine production by CD11c+ and CD11c− DC populations was assessed by intracellular staining. Cytokine detection was competitively inhibited by unlabelled antibody of the same clone as that used for cytokine detection, but not by unlabelled antibody of irrelevant specificity (fig 1).

Cytokine production by blood dendritic cells. Dendritic cells (DC) were identified by multicolour flow cytometry as an HLA−DR+ lineage negative (CD3−, CD14−, CD16−, CD19−, CD34−, and CD56−) population. Within this DC gate, CD11c+ and CD11c− DC subsets were identified. The cytokine production by these DC subsets was assessed by intracellular cytokine staining and super-enhanced Dmax normalised subtraction. One parameter histograms for the intracellular staining of IL-10 with and without added monensin are shown. In this example, cultures were stimulated with VSL#3 probiotic bacteria. Intracellular cytokine production was determined by subtracting labelling of the sample without monensin from labelling of the sample with monensin using WinList software. In the histograms with subtraction percentages, the outline of the histogram indicates staining of monensin treated cells and the shaded area represents the proportion of cytokine positive cells after the subtraction. Cytokine detection was competitively inhibited by unlabelled antibody of the same clone as that used for cytokine detection, but not by unlabelled antibody of irrelevant specificity.
VSL#3 upregulates IL-10 and downregulates IL-12 production by blood and colonic lamina propria dendritic cells
The probiotic combination VSL#3 at a dose equivalent to 108 CFU/ml upregulated IL-10 production and decreased IL-12 production by CD11c+ and CD11c− DC (fig 2). This increase in IL-10 production in the presence of VSL#3 was dose dependent over the range 106–109 CFU/ml with maximal IL-10 detected at 108 CFU/ml (data not shown). In contrast, there was a trend towards enhanced IL-12 production by DC in the presence of LPS. Co-culture of VSL#3 at 108 CFU/ml with LPS at 1 μg/ml significantly diminished the proinflammatory effects of LPS by decreasing IL-12 while maintaining IL-10 production (see fig 3 for representative experiment). In analysis of paired pooled data, addition of VSL#3 to LPS stimulated cultures significantly reduced the proportion of CD11c+ DC producing IL-12 (42.5 (SEM 9.85) % for LPS alone v 0% for LPS plus VSL#3; p<0.05, n = 3). IL-10 production induced by VSL#3 was not significantly altered by addition of LPS (57 (SEM 3.4) % IL-10+ CD11c+ DC for VSL#3 alone v 51.3 (SEM 12.9) % for VSL#3 plus LPS; n = 3).

Enhanced IL-10 production and decreased IL-12 production by blood DC in the presence of probiotic bacteria (VSL#3). Whole blood was incubated with control medium alone, VSL#3 probiotic bacteria (at a dose equivalent to 108 CFU/ml) or LPS (1 μg/ml) for four hours and (A) IL-10 production and (B) IL-12 production was assessed by intracellular cytokine staining. The proportion of cytokine staining CD11c+ and CD11c− DC is shown (mean (SEM), n = 9−13).

Co-culture of VSL#3 with LPS decreased the proinflammatory effects of LPS. In whole blood, VSL#3 at 108 CFU/ml and LPS at 1 μg/ml were co-cultured and cytokine production assessed by intracellular staining. The proportion of CD11c+ DC producing IL-10 and IL-12 in these co-cultures is shown. The data shown are representative of three independent experiments.
The enhanced IL-10 production by DC in whole blood with VSL#3 was confirmed by incubating an enriched DC population with VSL#3 and assessing IL-10 production by both intracellular cytokine staining (fig 4A) and ELISA (fig 4B).

Enhanced IL-10 production by an enriched DC population in the presence of probiotic bacteria (VSL#3). (A) An enriched DC population (metrizamide separated) was cultured with control medium alone, VSL#3 probiotic bacteria (at a dose equivalent to 108 CFU/ml) or LPS (1 μg/ml) for four hours and the IL-10 production assessed by intracellular cytokine staining. (B) The concentration of IL-10 in the supernatants of enriched DC cultured with control medium, VSL#3 (at a dose equivalent to 108 CFU/ml) or LPS (1 μg/ml) was measured by ELISA at four hours and 16 hours. The results are representative of three independent experiments.
To determine whether upregulation of IL-10 production by VSL#3 also occurred with intestinal DC, we examined cytokine production by human colonic LPDC. Intestinal LPDC were identified from lamina propria mononuclear cells as a CD11c+ HLA−DR+ lineage-negative (CD3−, CD14−, CD16−, CD19−, CD34−, CD56−) population (fig 5). This population had the phenotype of immature DC, consistent with our previous findings22 A proportion of unstimulated LPDC produced IL-10 (18.6 (SEM 6.5) %, n = 11) and this IL-10 production was upregulated following incubation with VSL#3 (44.1 (SEM 7.4) %, n = 9). A small proportion of unstimulated LPDC produced IL-12 (8.6 (SEM 3.8) %, n = 6) and this was not upregulated by VSL#3. In contrast, LPS enhanced the proportion of DC producing IL-12.

Cytokine production by lamina propria CD11c+ DC extracted from mucosal tissue. DC were identified by multicolour flow cytometry as HLA−DR+ lineage-negative (CD3−, CD14−, CD16−, CD19−, CD34−, and CD56−). Within this gate, CD11c+ DC were identified. The effect of control medium alone, VSL#3 (at a dose equivalent to 108 CFU/ml) or LPS (1 μg/ml) on IL-10 and IL-12 production by intestinal derived CD11c+ DC is shown (mean (SEM), n = 9−11).
Individual bacterial strains differentially modulate IL-10 and IL-12 production and expression of costimulatory/maturation markers by dendritic cells
Different bacterial strains evoked different cytokine responses when cultured with whole blood (fig 6). All bifidobacteria significantly upregulated IL-10 production by both CD11c+ and CD11c− DC and this increase in IL-10 was dose dependent over the range of concentrations 106–108 CFU/ml (data not shown). In contrast, lactobacilli decreased or had no significant effect on production of IL-10 by both CD11c+ and CD11c− DC. Streptococcal strains had no significant effect on IL-10-producing DC.

Effect of individual bacterial strains on IL-12 and IL-10 production by CD11c+ and CD11c− DC. When addition of bacteria to the culture increased cytokine production compared with that seen in the absence of any added stimulus for a given individual, the proportion of cytokine producing cells was taken as a net positive value. If the bacteria decreased cytokine production compared with that seen in the absence of bacteria, the proportion and absolute number of cells was taken as a net negative value. The data indicate the mean (SEM) for three to six independent experiments. (A) The effect of the different probiotic strains at 108 CFU/ml on IL-10 production by CD11c+ and CD11c− DC. (B) The effect of the different probiotic strains at 108 CFU/ml on IL-12 production by CD11c+ and CD11c− DC. * Significant changes in cytokine production in the presence of the bacterial stimuli.
None of the bacteria tested induced IL-12 production by CD11c+ or CD11c− DC. Some components of VSL#3, illustrated in figure 6, significantly inhibited IL-12 production.
VSL#3 strains differentially modulated DC expression of maturation and costimulatory molecules (table 2). The proportion of DC expressing CD80 was significantly less in cultures stimulated with the bifidobacteria strains than in unstimulated cultures: B infantis and B breve affected both CD11c+ and CD11c− DC; only CD11c− DC were significantly affected by B longum. In unstimulated cultures the majority of DC expressed CD40. None of the bacterial preparations significantly altered the proportion of CD40+ DC although there was a trend towards a reduction in cultures stimulated with bifidobacteria. Consistent with an effect of bifidobacteria on CD40 expression, analysis of the level of CD40 expressed demonstrated a significantly lower level of CD40 on CD11c+ DC in the presence of B infantis and B breve (data not shown). The proportion of DC expressing CD83 was also high in unstimulated cultures and there was a tendency for all bacterial strains to increase this proportion further, with L casei and L plantarum significantly increasing the level. CD86 was not significantly altered by any of the bacterial strains.
Effect of individual bacterial strains on costimulatory and maturation markers
VSL#3 treatment of dendritic cells inhibits generation of Th1 cells
To determine whether the ability of VSL#3 or its components to increase IL-10 production by DC influenced the subsequent T cell response, two different assays were performed. Purified naïve CD4+ T cells were activated in the presence of enriched allogeneic DC treated with VSL#3, LPS, or medium alone and the cytokine profile of the differentiated T cells was examined. Enriched DC were potent stimulators of allogeneic naïve CD4+ T cells irrespective of pretreament with bacterial products (data not shown). However, significantly fewer T cells committed to IFN-γ production were generated in cultures activated with VSL#3-treated DC (21.8 (SEM 5.0) %, n = 4) than in cultures stimulated with LPS treated DC (42.0 (SEM 4.1) %, n = 4) or unstimulated DC (36.3 (SEM 5.0) %, n = 4) (fig 7). The proportion of differentiated T cells making IL-10 or IL-4 did not differ significantly between the cultures.
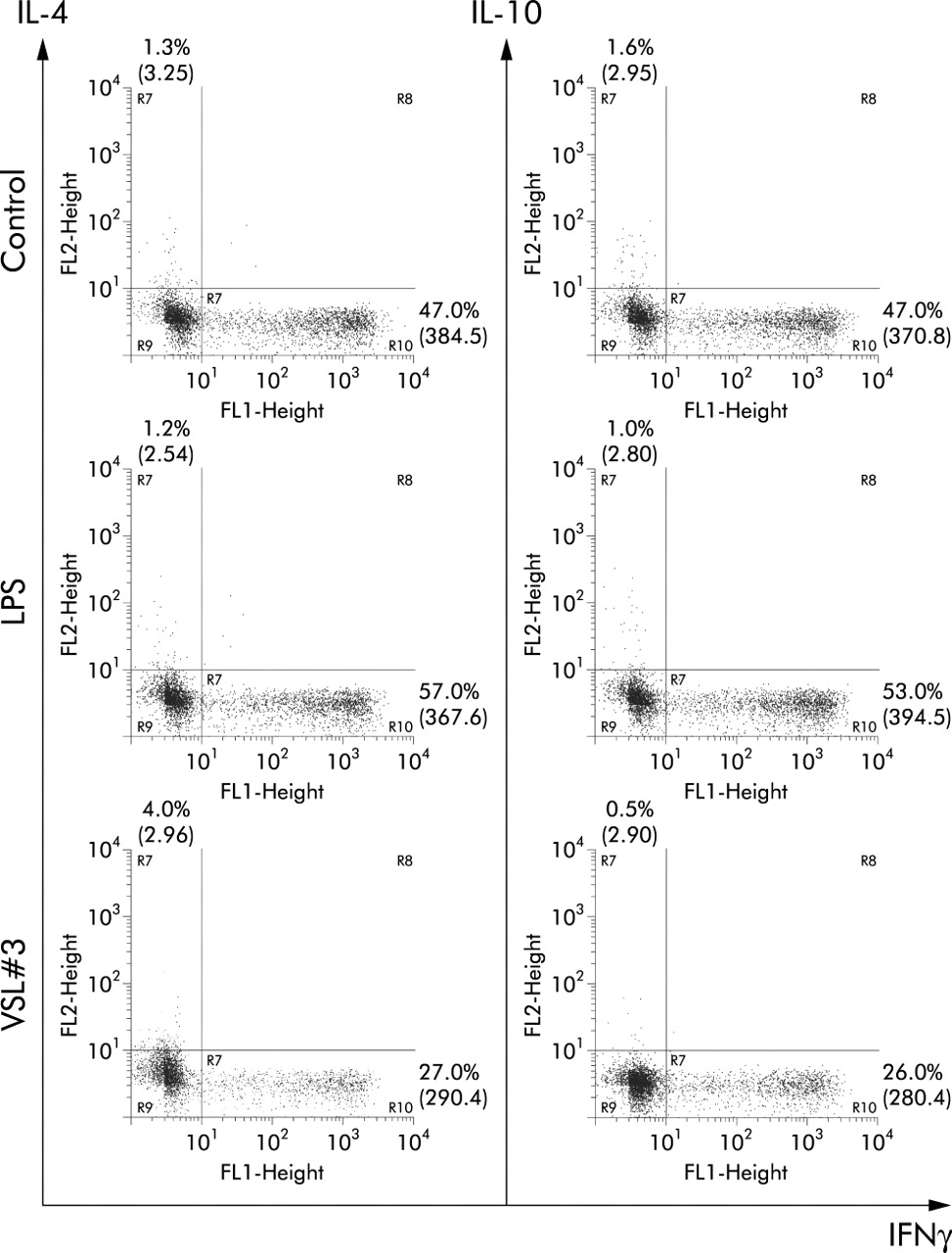
Probiotic bacteria (VSL#3) inhibit the generation of IFNγ-producing T cells. Naïve T cells and enriched DC treated with control medium, VSL#3 (at a dose equivalent to 108 CFU/ml) or LPS (1 μg/ml) were co-cultured. Cells were restimulated with PMA and ionomycin and cytokine production was assessed by intracellular cytokine staining. Staining for IFN-γ, IL-4, and IL-10 is shown. The proportion of cells and the geometric mean of fluorescence intensity are represented. Results are representative of four independent experiments.
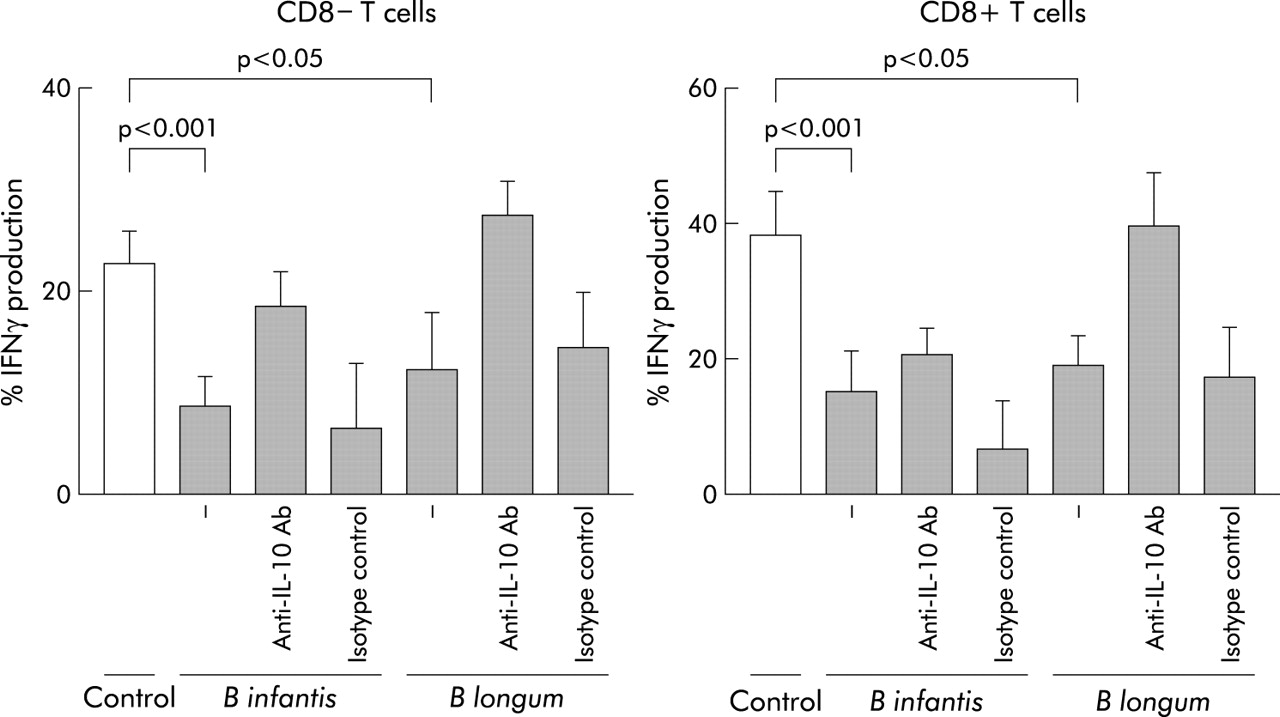
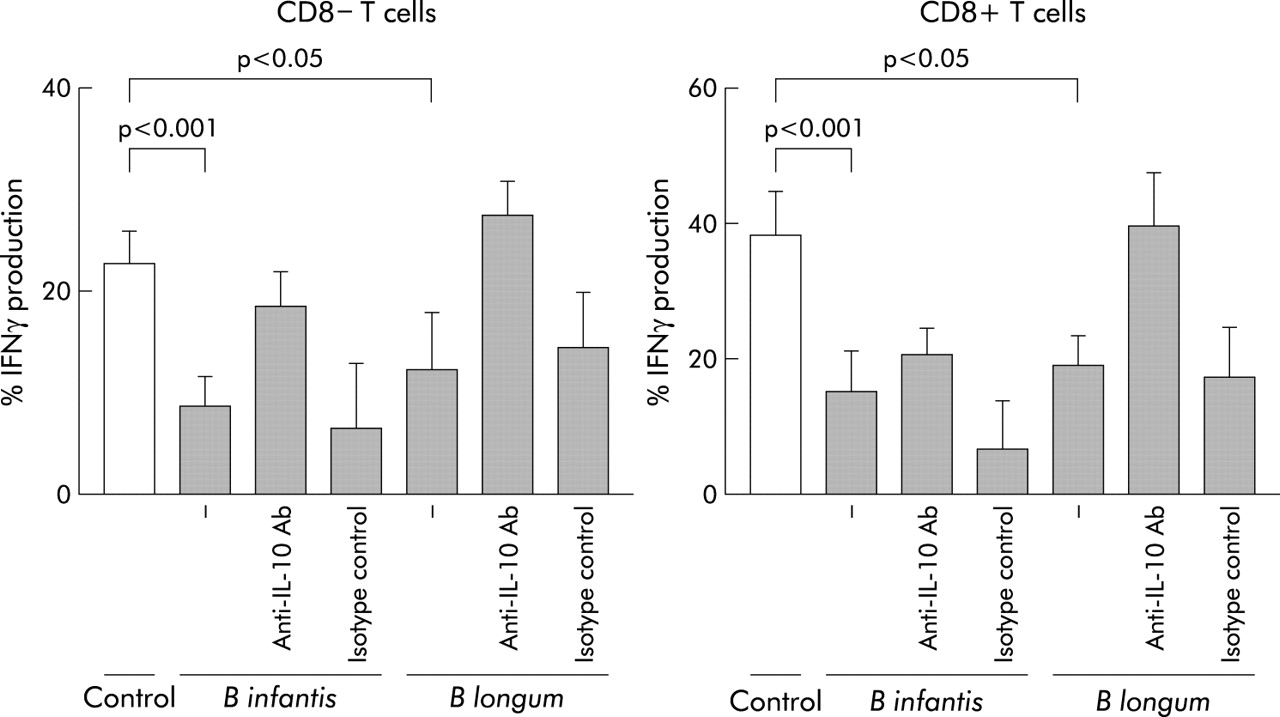
In addition, single probiotic strains were incubated with whole blood and the effect on IFN-γ production by CD8+ and CD8− T cells was assessed after polyclonal stimulation with PMA and ionomycin. Prior exposure to B longum and B infantis, but not the other bacteria tested, significantly reduced the proportion and absolute numbers of CD8− T cells and CD8+ T cells producing IFN-γ in a dose dependent manner (fig 8). Compared with medium control, the proportion of CD8− T cells producing IFN-γ was reduced by 46% with B longum (n = 9 independent experiments) and 62% with B infantis (n = 9 independent experiments); the proportion of CD8+ T cells producing IFN-γ was reduced by 50% (B longum) and 61% (B infantis). The effects of bacterial debris fractions on IFN-γ production were selective for cytokine production, rather than a reflection of reduced T cell activation or a toxic effect of the bacterial preparation, as over 97% were activated (CD69+) by PMA and ionomycin irrespective of prior exposure to bacteria (data not shown). To assess whether the decrease in IFN-γ by T cells was dependent on IL-10, a neutralising anti-IL-10 blocking antibody was added to some cultures. For both CD8+ and CD8− T cells, the inhibition of IFN-γ production by B longum was reversed significantly by a neutralising anti-IL-10 antibody, but not an isotype matched control. Anti-IL-10 reversed the inhibition of IFN-γ production by B infantis for CD8− T cells only. (fig 8).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Individual probiotic strains decrease IFN-γ production by polyclonally stimulated T cells. Individual probiotic strains at 108 CFU/ml were incubated with whole blood. Subsequently the cultures were stimulated with PMA and ionomycin and the effect of preculturing with the probiotic strain on cytokine production was observed. In some cultures an anti-IL-10 neutralising antibody or its isotype control were added and the effect on IFN-γ assessed. Results indicate the mean (SEM) of at least nine experiments.
DISCUSSION
The clinically active probiotic combination VSL#3 was a potent inducer of IL-10 by intestinal and blood DC and inhibited generation of proinflammatory Th1 cells. Individual strains within VSL#3 displayed distinct and diverse immunomodulatory effects. The most marked anti-inflammatory effect was shown by bifidobacterial species which upregulated IL-10 production by DC and decreased expression of the costimulatory molecules CD80 and CD40. These effects of probiotic bacteria on DC may underlie their anti-inflammatory activity.
The individual bacteria in VSL#3 displayed different effects on cytokine production by DC, arguing against a common bacterial component such as peptidoglycan mediating the effects. Differences between strains have been reported previously; B bifidum, B breve, and B infantis stimulated more IL-10 and less IL-12 and TNFα from a murine macrophage like cell line than B adolescentis.25 Moreover, there was no simple correlation between reported DC expression of TLR and responses to VSL#3. Thus responses of CD11c+ and CD11c− DC subsets to VSL#3 were broadly similar although these two populations express a different array of the known TLRs.26 The divergent patterns of responses to VSL#3 and expression of TLRs suggests that other receptors may be involved in DC recognition of VSL#3 or that recognition of VSL#3 components by TLRs on non-DC may contribute.
Intestinal DC are functionally different from those at other sites.3,6,27 In vitro, VSL#3 increased the proportion not only of blood DC making IL-10 but also of colonic lamina propria DC making IL-10, suggesting that one mechanism of action of VSL#3 in vivo is to trigger DC production of anti-inflammatory IL-10 in intestinal mucosa. Patients with pouchitis taking VSL#3 have increased mucosal levels of IL-10 and decreased levels of IFN-γ28 and in murine colitis, proinflammatory TNF-α and IFN-γ are decreased following treatment with VSL#3.29 Enhanced production of IL-10 by DC may limit mucosal inflammation either by direct anti-inflammatory effects or by enhancing the generation or activity of regulatory T cells (Tr1). In experimental colitis, local administration of IL-10 in bacterial30 or viral31 vectors ameliorates inflammation. Adoptive transfer of regulatory T cells cures or prevents intestinal inflammation in murine colitis by IL-10 dependant mechanisms.32–34 In another mucosal environment in the lung, high concentrations of IL-10 produced by pulmonary DC are associated with the generation of regulatory T cells.2
Bacteria may differentially alter antigen presentation and the level of DC activation. Bifidobacteria generally downregulated the proportion of DC expressing CD80 but not the proportion of those expressing CD86. This differential effect between members of the B7 family of costimulatory molecules may be related to the difference in proportions of DC expressing CD80 and CD86 in unstimulated cultures. Confirming other work,35 we found more DC expressing CD86 than CD80 in the absence of stimulation. Thus, CD80 expression may be more readily modulated through environmental signals. Both CD80 and CD86 bind to CD28 and CD152 (CTLA4) on T cells. It remains controversial whether they have distinct functions,36 but relative amounts of CD80 and CD86 on antigen presenting cells may influence the type of T cell response generated.
Effects of bifidobacteria were not confined to CD80 and CD86. B breve and B infantis reduced the level of CD40 expression on DC. Signalling through CD40 increases IL-12 production by DC and enhances their survival.37,38 Increased levels of expression of CD40 on DC39 and its ligand, CD154, on T cells40 in IBD tissue suggests that this interaction is important in mucosal inflammation. VSL#3 treatment of IBD may mediate downregulation of CD40 expression, breaking the inflammatory cycle of mutual T cell-DC stimulation.
In contrast to bifidobacteria, lactobacilli appeared to generate “semi-mature” DC,41 a phenotype characterised by increased costimulatory marker expression but low production of proinflammatory cytokines. Such “semi-mature” DC may contribute to the production of regulatory T cells and subsequent tolerance in vivo.41
VSL#3 inhibited T cell production of IFN-γ in two different systems. This reduction in proinflammatory cytokine production was not accompanied by an obvious increase in either IL-4-producing or IL-10-producing T cells arguing against a simple model in which VSL#3 induces a switch from Th1 to Th2 production. On the basis of the current data, its is not possible to determine whether the effect of VSL#3 treated DC acts at the level of T cell instruction or whether it operates by selection and expansion of different T cell precursors. DC stimulated comparable proliferative responses irrespective of bacterial treatment, suggesting that a major effect on lymphocyte viability is unlikely. IL-10 may contribute to the effect on T cells because neutralising IL-10 partially reversed VSL#3 mediated downregulation of IFNγ.
In conclusion, VSL#3, which is clinically beneficial in the treatment of intestinal inflammation, changes the cytokines in DC from an inflammatory towards a regulatory profile, which may be instrumental in the immunomodulatory activity of probiotic treatments. An increased understanding of the effects of probiotic organisms on the immune system may enable refinement of probiotic treatments and enhance our knowledge of immune homeostasis in the specialised environment of the intestine.
Acknowledgments
This work was supported by grants from the Wellcome Trust and Medical Research Council (UK).