Article Text

Abstract
Background: Human immunodeficiency virus/hepatitis C virus (HIV/HCV) coinfection is a common and complex clinical problem in which loss of immunological control of HCV occurs, with increased HCV viral load and more aggressive liver disease. Cellular immune responses, particularly secretion of interferon γ (IFN-γ) appear to be important in the control of HCV, and a detectable HCV specific CD4 response is associated with clearance of the virus. HCV specific CD8+ T cell responses, weak in chronic HCV infection, have been shown to be further impaired in HIV coinfection and this CD8+ T cell deficiency is related to the decline in CD4 T cell count.
Aims: To compare the CD4 T cell response to HCV in HIV/HCV coinfected and HCV monoinfected individuals and to determine the relationship of responses with declining CD4 count.
Patients: The study subjects were a cohort of 68 HCV monoinfected and 67 HCV/HIV coinfected haemophiliac children and adolescents (the Hemophilia Growth and Development Study) who were followed for a seven year period.
Methods: We analysed IFN-γ secreting CD4+ responses to HCV proteins and peptides and HIV p24 antigen using an ELISpot assay.
Results: We found a significant decrease in HCV specific responses among those who were HIV coinfected (10/67 v 36/68; p<0.0001) both in numbers of responders and frequency of specific cells. This did not appear to be closely related to CD4 count.
Conclusions: The reduction in HCV specific CD4 T cells in coinfection provide a cellular mechanism for the loss of control of HCV in coinfected individuals, even in those with relatively preserved CD4+ T cell counts and CD4+ T cell responses to HIV.
- HCV, hepatitis C virus
- HIV, human immunodeficiency virus
- IFN-γ, interferon γ
- EBV, Epstein-Barr virus
- HAART, highly active antiretroviral therapy
- PBMC, peripheral blood mononuclear cell
- hepatitis C virus
- human immunodeficiency virus
- CD4+ T cells
- coinfection
Statistics from Altmetric.com
- HCV, hepatitis C virus
- HIV, human immunodeficiency virus
- IFN-γ, interferon γ
- EBV, Epstein-Barr virus
- HAART, highly active antiretroviral therapy
- PBMC, peripheral blood mononuclear cell
Coinfection with human immunodeficiency virus (HIV) and hepatitis C virus (HCV) is a common problem worldwide, with up to 80% of intravenous drug users and 98% of haemophiliacs infected with both viruses.1 There is increasing evidence that such coinfection can affect the clinical course of either disease2 and, in particular, HCV induced liver disease is a major problem in HIV positive cohorts. These important clinical effects may potentially arise from virological interactions or, very likely, an underlying immunological mechanism.
On their own, the two viruses exhibit differing immunological profiles in chronic persistent infections. HIV induces strong CD8+ T cell responses which are sustained but poor proliferative HIV specific CD4 responses. In contrast, in HCV monoinfection, virus specific CD8 cells are rarely detected in peripheral blood. HCV specific CD4 responses can be readily detected in individuals who spontaneously resolve the infection but are typically weaker in those who develop persistent infection.3 In coinfected individuals, the HCV specific CD8 response, but not that to HIV or Epstein-Barr virus (EBV), has been shown to be strongly dependent on absolute CD4 count,4 indicating a differing requirement for CD4 “help” in maintaining responses to HCV.
The exact role of CD4 responses in control of HCV is unclear but it is thought that IFN-γ secretion is significant. Intrahepatic secretion of IFN-γ by T cells is thought to provide a major antiviral effect. Depletion of CD4+ T cells in animal models leads to failure to control virus, accompanied by the emergence of CD8+ T cell escape mutants.5
The likely explanation for the loss of CD8+ T cell responses in coinfected individuals is through depletion of antigen specific CD4 responses as a result of loss of CD4 cells in HIV infected subjects. In order to investigate whether coinfection with HIV directly influences HCV specific CD4 responses, we have quantitated CD4+ T cell responses to HCV peptides and proteins by ex vivo interferon γ (IFN-γ) ELISpot in a large cohort of coinfected and HCV monoinfected haemophiliac children. Here we report a profound difference in the level of HCV specific CD4+ responsiveness in the two groups.
PATIENTS AND METHODS
Patients
The Hemophilia Growth and Development Study was a multicentre US study that enrolled, from 1989 to 1991, children and adolescents aged 6–19 years old with haemophilia; 126 were HCV monoinfected and 207 were coinfected with HIV-1 and HCV. Details of recruitment and characteristics of this cohort have been reported elsewhere.6 During a seven year follow up period, blood samples were taken every six months and lymphocytes frozen within 24 hours. Two of these frozen samples, with a median time interval of 5.1 years, were selected for study from 103 coinfected and 83 monoinfected individuals and only those samples with high viability and sufficient cell numbers following thawing were selected for assay (mean number of viable cells recovered from HCV monoinfected samples = 2.54×106 (range 0–12.8); coinfected mean = 2×106 (range 0–9.12)). Details of CD4 count, and HIV and HCV viral load were available for each sample and none of the participants had received highly active antiretroviral therapy (HAART) therapy during the trial.
Peptides and recombinant antigens
HCV antigens used in the analysis were 18 HCV core sequence peptides (20mers overlapping by 10) covering amino acids 1–191 which were pooled to give a final concentration of 10 μg/ml of each peptide in the ELISpot assay. Recombinant proteins NS3, NS4, and NS5 (Chiron, Emeryville, California, USA) were also pooled to give a final concentration of 1 μg/ml for each antigen in the assay. Recombinant HIV-p24 (NIBSC, Potters Bar, UK) was used in the assays of coinfected individuals at a concentration of 1 μg/ml.
ELISpot assay for IFN-γ secretion
The ELISpot assay was performed as previously described7 using 200 000 cells per well and plates read with an AID plate reader. Each sample was tested in duplicate against the HCV antigens, and against HIV p24 in the coinfected group, and PHA was used as a positive control. Samples which did not show a PHA response were excluded from the analysis. A response was considered positive if the number of spots was equal to or greater than 2.5 times background (cells alone).
Statistical methods
Statistical analyses were performed by Fisher’s exact or the Mann-Whitney test using Prism 3.0 software.
RESULTS
Subjects were considered to have a positive CD4+ T cell response to HCV if peripheral blood mononuclear cell (PBMC) samples from either time point gave a positive ELISpot response to a HCV antigen. There were not sufficient lymphocytes recovered from the samples after thawing to allow for depletion of the CD8+ population but two previous studies from this group using identical antigens3,8 have shown that ELISpot responses to pooled 20mer core peptides and to both recombinant HCV and recombinant HIV p249 are primarily from the CD4 compartment, although some contribution by CD8 cells cannot be ruled out.
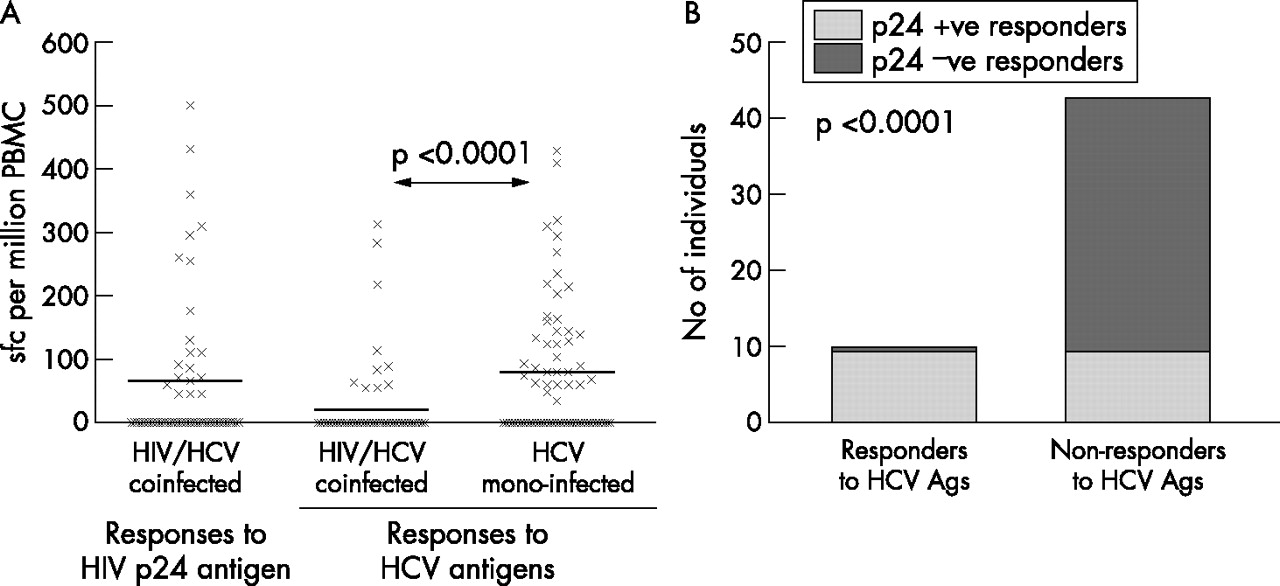
Of the 68 HCV monoinfected individuals tested, 36 (52%) had a CD4 T cell response to HCV while such responses were detected in only 10/67 (15%) coinfected individuals (p<0.0001) (fig 1⇓). Responses were directed at both HCV core and non-structural proteins in both groups but in the monoinfected group there seemed to be a broader response—core peptides were recognised by all 36 of the monoinfected group of responders and 19 of these also recognised NS3-5. In the coinfected group, four recognised core peptides only, five recognised NS3-5 proteins only, and one subject recognised both. Where positive, the frequency of HCV specific cells was similar for either HCV antigen (mean number of spots per million PBMC for core peptides = 152, for NS3-5 = 157 in the monoinfected group, and 104 and 171, respectively, in the coinfected group). Comparison of the peak responses to HCV antigens of the total cohort revealed that the monoinfected group had a significantly higher frequency of IFN-γ producing HCV specific CD4 cells (p<0.0001) (fig 2A⇓).
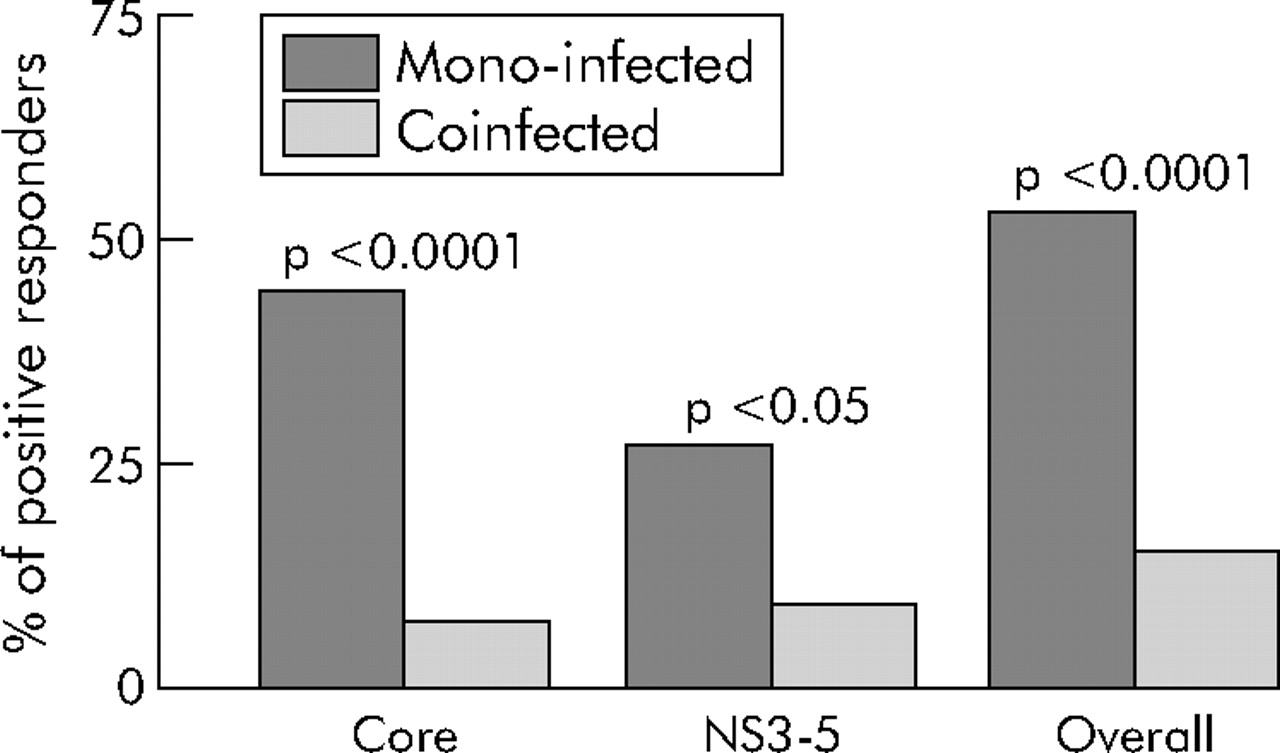
ELISpot responses to hepatitis C virus (HCV) antigens. The percentage of individuals in each group who had a CD4+ T cell interferon γ ELISpot response to HCV antigens is shown. Statistical comparisons were performed using Fisher’s exact test.

{kind=link}
{kind=link}
(A) Frequency of hepatitis C virus (HCV) or human immunodeficiency virus (HIV) p24 specific interferon γ (IFN-γ) producing CD4 cells in each individual. Peak frequency of IFN-γ producing CD4 cells per million peripheral blood mononuclear cells (PBMC) from each individual recognising recombinant HIV p24 or HCV antigens (pooled recombinant NS3-5 antigens or pooled core peptides). Statistical comparison was performed using a Mann-Whitney test. (B) Association between HCV and HIV responses in coinfected individuals. Bars represent the response to HIV p24 antigen in the HIV/HCV coinfected group. HCV antigen responders (n = 10) all responded to p24. Of the 43 HCV antigen non-responders, only 10 responded to HIV p24. Statistical comparison was performed by Fisher’s exact test.
A response to HIV-p24 among coinfected individuals was detected in 20 of 53 tested with this antigen (37%). This is a comparable rate of detection to other studies using cryo-preserved cells.10 The frequency of IFN-γ producing cells, where detected, was a mean of 163 per million PBMC, similar to that for HCV specific cells in positive individuals (fig 2A⇑), and suggests that, overall, failure to detect a HCV response was not the result of a technical deficiency in the assays of the coinfected group. Interestingly, HCV CD4 responses were only detected in those who also had a HIV-p24 response (p<0.0001) (fig 2B⇑), an “all or nothing” phenomena in coinfection that we have recently described for HCV CD8 responses in this cohort.11
As it is often proposed that loss of control of HCV replication in coinfected individuals may be due to loss of specific CD4 cells, we looked to see if there was a relationship between CD4 count or viral load and the HCV specific CD4 response in those that were coinfected. Table 1⇓ shows the mean HCV viral load among those with and without a detectable HCV specific CD4 response in the mono and coinfected groups plus HIV viral load, CD4 counts, and change in CD4 counts over time in the coinfected group. Although HCV viral load was lower among those with HCV responses than among those without, the differences were not significant and hence we could not identify a clear significant relationship between HCV viral load and the presence of a HCV specific CD4 response. There was however a substantial difference in HCV viral load observed between the total mono and coinfected group (p<0.0001) (table 1⇓).
Comparison of clinical details of hepatitis C virus (HCV) CD4 responders
The HCV specific CD4 responses in the coinfected individuals were unexpectedly not influenced by CD4 count as responses could be detected in subjects with a CD4 count of less than 100 (data not shown). HIV viral load, initial CD4 count, and progress of HIV disease, as measured by change in CD4 count, did not seem to influence the presence or absence of a HCV specific CD4 response in the coinfected group.
DISCUSSION
The impact of coinfection on the clinical course of virus infections is significant, particularly in HIV where advances in antiretroviral therapy have succeeded in delaying the onset of AIDS but where the patient can develop severe clinical disease from other persistent viruses, such as HCV, CMV, or EBV. It is clear that HIV has an effect on HCV infection as there is a very evident loss of control of HCV viraemia in coinfected patients.1 The question is whether this is the result of a specific failure of the HCV immune response, a more general immunological effect, or even direct viral interactions. There is much evidence to show that the CD4 response to soluble antigens such as tetanus toxoid in HIV infected individuals declines as the disease progresses12 but the effect on responses to persistent viruses, where the immune system is constantly exposed to antigen, is less clear.
A previous study of the cellular immune response to HIV and HCV in coinfected individuals also demonstrated profound impairment of the HCV response. Lauer and colleagues13 compared groups of coinfected and HIV monoinfected individuals, including those on HAART therapy and a group of HCV monoinfected individuals, looking at the CD4 response to HIV and HCV recombinant proteins by proliferation of fresh PBMC. Responses to HCV proteins were not detected in any of the coinfected individuals, in contrast with the HCV monoinfected group where 47% had detectable responses. The discrepancy between the absence of any detectable HCV responses in the coinfected group in the study by Lauer et al, compared with the 15% of responders detected in our study, is likely to be due to different sensitivities in the techniques used—proliferation is measured after culturing PBMC for six days whereas the ELISpot measures events happening after only 18 hours in culture. The proliferative capacity of the responses we detected was not assessed here, and further studies are required to analyse their full functional status. Detection of responses to HIV-p24 in the coinfected group was similar to those reported here (9/17), was associated with HAART control of HIV plasma viraemia, and was much lower in the HIV monoinfected control group (1/8). This suggests a beneficial effect of HCV infection on the HIV immune response whereas the reverse seems true for the effect of HIV on HCV infection. While HAART treatment has been shown to improve proliferative responses to some recall antigens, it does not appear to increase the HCV response, suggesting again that the HCV immune response is relatively sensitive to the effects of HIV. The study reported here is the first to show the effect of coinfection on the HCV CD4 T cell response without the influence of successful HAART therapy and the first to analyse ex vivo IFN-γ secreting CD4+ T cell responses, thought to be a critical cell type in the control of viral replication.
A study by Kim and colleagues4 of CD8+ T cell responses to HIV and HCV antigens in coinfection found a significant association between diminishing numbers of HCV specific CD8+ T cells and HIV-1 associated CD4+ T cell depletion (p<0.0001) but no association between the frequency of EBV or HIV-1 CD8+ T cells and declining CD4 counts. Taken together, these data suggest that the reduction in HCV specific cells occurs readily during HIV coinfection and has a profound influence on cellular immunity against HCV. However, we have found no clear relationship between declining CD4 count and reduced frequency of HCV specific CD4 responses. Loss of other helper functions rather than IFN-γ secretion may be more critical in explaining the link between CD8 responses and declining CD4 count in the study by Kim and colleagues.4
Studies of coinfection of HIV with other persistent viruses such as CMV provide further evidence of the sensitivity of the HCV CD4+ T cell response to the suppressive effect of HIV. Waldrop and colleagues14 reported that the frequency of IFN-γ producing CMV specific CD4 cells detectable in a group of HIV infected adults was higher than in healthy HIV negative individuals (p<0.03). This was unrelated to the clinical stage of HIV infection and was not observed in CD4 responses to recall responses to mumps or measles antigens where 40% of HIV positive individuals showed a response in comparison with 100% of HIV negative individuals. Strong CD8 and CD4 responses to CMV are sustained over time during HIV infection with a decline in CMV specific CD4+ cells, preceding the onset of CMV associated end organ disease in late acquired immunodeficiency syndrome.14
There have been fewer reports of CD4 responses in coinfection with EBV and HIV. EBV specific CD4 cells are difficult to detect ex vivo during latent EBV infection and in a study of CD4 responses to EBV antigens in a group of healthy EBV seropositive donors and a group infected with HIV, Piriou and colleagues15 were unable to detect differences in the frequency or strength of the response directly ex vivo. After expansion of specific T cells for 6–12 days with EBV peptides, they found comparable numbers of EBV specific CD4 cells in healthy EBV carriers and HAART treated HIV positive individuals while the frequency was lower in untreated HIV positive subjects, although this was only for cells recognising the latent EBNA1 antigen.
The Hemophilia Growth and Development Study cohort studied here represents a unique group of individuals who all acquired HCV and/or HIV infection parenterally and at an early age. None had received HAART. They therefore represent an opportunity to study the natural course of the immune response during coinfection. Evidence from this study and that of Lauer and colleagues13 indicate that the CD4 response to HCV, detectable fairly readily in monoinfection, is severely compromised in coinfection with HIV. The diminished frequency of HCV specific CD4 T cells appears to be specific for this virus as 38% of those coinfected had a detectable HIV specific responses but only 15% a HCV specific response, and none of the coinfected group had a response to HCV only. Although the effect of HIV infection on the CD4 response to recall antigens has been clearly documented, this is perhaps more easily explained by a generalised loss of CD4 cells and failure to replenish specific memory cells if the antigen is not re-encountered. In the case of persistent infections however, even in the face of a low frequency of specific CD4 cells, the constant antigenic stimulation, particularly in the presence of the high viral load present in HCV, might be expected to replenish the CD4 pool as appears to be the case in CMV.16 An alternative likely scenario for HCV is that of antigen induced exhaustion whereby the constant antigenic stimulation leads to loss of specific cells, as is evident in LCMV.17
An alternative explanation for our findings is that the HCV specific CD4 cells are compartmentalised within the liver18 and so would not be detected in PBMC. HCV specific CD4 cells can be grown out of liver biopsies in both HCV monoinfected and HIV/HCV coinfected individuals at equal frequencies but there does seem to be a reduction in their capacity to secrete interleukin 10 in the presence of HIV infection.19 Liver biopsies had not been performed on this cohort as they are haemophiliacs and it seems unlikely that compartmentalisation would be a HIV specific effect, sufficient to explain on its own the profound differences between the mono and coinfected groups.
This study provides further evidence of a pathway whereby impairment of one arm of the immune response to HCV in HCV/HIV infection leads to further loss of control of HCV viraemia. The clinical progress of HCV infection seems to correlate closely with the effect of the virus on specific CD4 cells. Indeed, persistence of CD4+ T cells in the chronic infection, albeit at relatively low levels, may nevertheless play some protective role in the liver during HCV monoinfection given the obvious clinical impact associated with their depletion in coinfection. In HIV/HCV coinfection, where HAART can restore CD4 count and delay progression to AIDS, further studies are required to identify prospectively to what extent restoration of the HCV specific CD4+ T cells can be achieved. Such restoration may be highly relevant to prevention of death from liver disease which is emerging as a significant problem as the HIV epidemic evolves.
Acknowledgments
This work was supported by the James Martin School of the 21st Century, Oxford, the Wellcome Trust, and the National Institutes of Health, National Institute of Child Health and Human Development (HD41224), Bethesda, MD, USA.
REFERENCES
Footnotes
Published online first 16 March 2006
Funding was provided by the EU.
Conflict of interest: None declared.
Linked Articles
- Digest