Article Text

Abstract
Background: We previously showed that neonatal maternal separation (MS) of rat pups causes immediate and long-term changes in intestinal physiology.
Aim: To examine if administration of probiotics affects MS-induced gut dysfunction.
Methods: MS pups were separated from the dam for 3 h/day from days 4 to 19; non-separated (NS) pups served as controls. Twice per day during the separation period, 108 probiotic organisms (two strains of Lactobacillus species) were administered to MS and NS pups; vehicle-treated pups received saline. Studies were conducted on day 20, when blood was collected for corticosterone measurement as an indication of hypothalamus–pituitary–adrenal (HPA) axis activity, and colonic function was studied in tissues mounted in Ussing chambers. Ion transport was indicated by baseline and stimulated short-circuit current (Isc); macromolecular permeability was measured by flux of horseradish peroxidase (HRP) across colonic tissues; and bacterial adherence/penetration into the mucosa was quantified by culturing tissues in selective media. Colonic function and host defence were also evaluated at day 60.
Results: Isc and HRP flux were significantly higher in the colon of MS versus NS pups. There was increased adhesion/penetration of total bacteria in MS pups, but a significant reduction in Lactobacillus species. Probiotic administration ameliorated the MS-induced gut functional abnormalities and bacterial adhesion/penetration at both day 20 and 60, and reduced the elevated corticosterone levels at day 20.
Conclusions: The results indicate that altered enteric flora are responsible for colonic pathophysiology. Probiotics improve gut dysfunction induced by MS, at least in part by normalisation of HPA axis activity.
Statistics from Altmetric.com
Intestinal epithelial cells are continuously exposed to noxious bacteria, antigens and toxins in the gut lumen, as well as to beneficial commensal organisms. The balance between potentially pathogenic and non-pathogenic flora is critical for homeostasis and preventing intestinal disease. For example, in patients with inflammatory bowel disease (IBD)1 and irritable bowel syndrome (IBS),2 there are changes in spatial organisation and composition of gut organisms compared with control individuals, with reduced levels of Lactobacillus and Bifidobacterium species. Animal models indicate a role for bacteria in the development of intestinal inflammation;3 4 disease improvement occurs following manipulation of flora with antibiotics or probiotics.5 6 Models of spontaneous intestinal disease, including genetic deficiencies in interleukin 10 (IL10),7 IL28 and T cell receptors,9 are spared from developing inflammation when the mice are housed under germ-free conditions.
The colonisation process begins at parturition, is crucial for long-term maintenance of the microbiota, and results in the development of gut defences such as secretory immunoglobulin A (sIgA) production (which does not develop in germ-free animals).10 Age-dependent changes in microbiota exist, with altered levels of Enterobacteriaceae, Clostridium and Bacteroides from the suckling period through weaning in rats.11 Colonisation with certain strains of bacteria may be important in preventing the development of intestinal diseases including colitis. Studies have reported that colonising IL10 gene-deficient mice, which spontaneously develop colitis as adults, with Lactobacillus species as neonates can prevent development of disease.12 Exposure of animals to stress during establishment of the intestinal microflora may result in altered colonisation, thereby predisposing them to the development of diseases in later life.
Neonatal maternal separation (MS) is a well-established model of early life trauma.13 14 Studies have shown that early psychological trauma in neonates results in long-lasting alterations in colonic physiology including increased secretory state,15 enhanced macromolecular permeability,15 increased motility16 and visceral hyperalgesia.16 17 For these reasons, MS has been suggested to be a model of IBS,16 a condition where child abuse has often been documented.18 MS also adversely affects gut physiology at the time of separation, as reported in rat pups where the mucosal barrier was found to be defective.19 Alterations in mucosal barrier function in neonates may result in increased antigen uptake, sensitisation and susceptibility to stress-induced dysfunction in later life. Postnatal microbial colonisation in mice has been shown to play an important role in the development of the hypothalamus–pituitary–adrenal (HPA) axis,20 which is essential for the normal stress response, and has been shown to be altered in animals exposed to MS.13 21
This study was designed to determine if changes in colonic flora are present in rat pups during MS and if re-establishing normal flora can prevent colonic barrier dysfunction. We found that probiotic administration to neonatal rats during separation restored colonic flora and repaired the gut barrier, effects which persisted until adulthood. Furthermore, probiotics reduced the elevated corticosterone in MS pups, suggesting that their effects on the gut were mediated at least in part via normalisation of HPA axis activity.
METHODS
Animals
Primiparous timed-pregnant Sprague–Dawley female rats were obtained from Charles River Laboratories (St. Constant, Quebec) on gestational days 13–14. Dams were individually housed in cages lined with chip bedding on a 12:12 h light–dark cycle (lights on at 8:00 h) with free access to food and water. Day of birth was considered to be day 1. All procedures were approved by the Animal Care Committee at McMaster University.
Study design
Neonatal rat pups were exposed to MS from day 4 until day 19, and sacrificed the following day. Rats were killed by decapitation, trunk blood was collected, and colonic tissue was excised and placed in Kreb’s buffer. Segments were mounted in Ussing chambers for functional studies, with additional segments processed for bacterial culture studies. In order to study the long-term effects of probiotics, a separate group of animals was subjected to neonatal MS while treated with probiotics or vehicle, weaned on day 23, and on days 60–70 exposed to a mild stressor (30 min of water avoidance stress (WAS)). Functional studies were then performed.
Maternal separation
Dams and their litters (culled to 12 pups) were randomly assigned to the MS protocol or to the control non-separation (NS) protocol. MS pups were individually separated from the dam for 3 h per day from days 4 to 19 of life, whereas control pups (NS) remained in their home cage with the dam and were left undisturbed. MS pups were removed from the home cage, taken into an adjacent room and placed into individual compartments within plastic cages lined with chip bedding. Cage temperature was maintained at 37±0.5°C by heating pads. Separation was performed between 9:00 and 12:00 h, minimising the effects of circadian rhythm.
Corticosterone
HPA axis activity was assessed by measuring corticosterone. Serum was isolated from trunk blood via centrifugation and frozen at −20°C until analysis. A radioimmunoassay kit (ICN, Costa Mesa, CA, USA) was used to measure the serum corticosterone concentration.
Ussing chamber studies
Segments of distal colon were cut along the mesenteric border and mounted in Ussing chambers (WP instruments, Narco Scientific, Mississauga, ON, Canada), exposing 0.6 cm2 of tissue area to 8 ml of circulating oxygenated Kreb’s buffer maintained at 37°C. The buffer consisted (in mM) of: 115 NaCl, 1.25 CaCl2, 1.2 MgCl2, 2.0 KH2PO4 and 25 NaHCO3 at pH 7.35±0.02. Additionally, glucose (10 mM) was added to the serosal buffer as a source of energy, which was balanced by mannitol (10 mM) in the mucosal buffer. Agar–salt bridges were used to monitor potential differences across the tissue and to inject the required short-circuit current (Isc) to maintain the potential difference at zero as registered by an automated voltage clamp. A computer connected to the voltage clamp system recorded Isc continuously. Baseline Isc values were obtained at equilibrium, ∼15 min after the tissues were mounted. Isc, an indicator of active ion transport, was expressed in μA/cm2.
Electrical field stimulation
The function of enteric nerves was examined by electrical field stimulation (EFS). EFS of enteric nerves was performed in Ussing chambers by passing current pulses from one side of the tissue to the other via stimulating electrodes and a Grass stimulator (Grass Medical Instruments, Quincy, MA, USA).22 The stimulus consisted of 10 mA, 10 Hz and 0.5 mS pulse width delivered in trains for a total of 5 s. EFS resulted in a transient increase in Isc which was measured as the difference between the basal and maximal Isc. The cholinergic component of the EFS was obtained by subtracting the response in the presence of cholinergic antagonists (the muscarinic antagonist atropine (10−5 M) and/or the nicotinic antagonist hexamethonium (10−5 M)) from the overall response. To determine whether differences in responses in tissues from MS versus NS pups were due to abnormalities in the epithelium (vs changes in enteric nerves), the ΔIsc to the cholinergic muscarinic agonist, bethanechol (10−4 M), in the presence of the neural blocker, tetrodotoxin (10−6 M), was determined. Concentrations selected were those which were maximally effective based on preliminary experiments or previously reported findings.23
Macromolecular permeability
Horseradish peroxidase (HRP) (44 kDa) was used as a probe to examine the effects of MS on macromolecular permeability. Type VI HRP (Sigma Chemical Co, St Louis, MO, USA) was added (10−5 M) to the luminal buffer 15 min following mounting. Serosal samples (500 μl) were taken at 30 min intervals for 2 h and replaced with fresh buffer to maintain constant volume. Enzymatic activity of HRP was measured using a modified Worthington method24 employing a kinetic assay, as previously described.19 The flux of HRP from the mucosa to the serosa was calculated as the average value of two consecutive stable flux periods (60–90 and 90–120 min). HRP flux was expressed as pmol/cm2/h.
Bacterial adherence and penetration
Distal colonic tissue samples were collected for the measurement of adherent and penetrating bacteria. Segments of 1 cm were excised, weighed and washed thoroughly in sterile saline. Pieces were then homogenised in sterile Triton X-100 (0.1%) and diluted serially. Dilutions were then spread on blood agar plates (PML Microbiologicals, Mississauga, ON, Canada) and incubated at 37°C for 24 h. To study Lactobacillus species specifically, samples were plated on selective deMan Rogosa Sharpe (MRS; EMC Chemicals, Gibbstown, NJ, USA) agar and incubated anaerobically at 37°C for 48 h. Colonies were then counted and expressed as colony-forming units per gram of tissue (CFU/g).25
Effect of probiotics
A commercially available probiotic preparation containing live Lactobacillus rhamnosus strain R0011 (95%) and L helveticus strain R0052 (5%) (Lacidofil®) was provided by Institut Rosell-Lallemand Inc (Montreal, PQ, Canada). This particular combination of Lactobacillus species prevented the altered host–microbial response associated with chronic WAS in adult rats,26 Citrobacter rodentium-induced colitis in mice27 and enterohaemorrhagic Escherichia coli-induced rearrangement of cytoskeletal proteins in human intestinal cells.28 The freeze-dried mixture was re-hydrated in sterile water and 10 μl of probiotic mixture (108 organisms) was administered (orally and rectally) twice a day between days 4 and 20 to MS and NS pups. (This dose was based on a previous publication.12) Litters were randomly selected for probiotic or vehicle administration.
Probiotics have been shown to colonise the gut transiently.29 PCR was used to confirm this transient colonisation of specific Lactobacillus species in probiotic-treated animals. Faecal samples from MS and NS colons were collected at sacrifice and stored at −20°C until further use. DNA was extracted using a faecal extraction kit (Qiagen, Mississauga, ON, Canada) and stored at −20°C. PCR was performed using specific primers for L rhamnosus strain R0011 (Institut Rosell-Lallemand) since this strain is the predominant one (95%) in the probiotic mixture. Two different reactions were performed to identify this strain accurately, as indicated by the manufacturer. The PCR mixture contained 0.2 mM each dNTP, 1 pmol/μl of each primer (table 1), 5 μl of template DNA, 0.05 U/μl of Taq polymerase and 1× PCR buffer. The amplification for PCR 1 consisted of 30 s denaturation at 94°C, 30 s annealing at 53°C and a 1 min extension at 72°C for 30 cycles. Conditions for PCR 2 included a 30 s denaturation at 94°C, 30 s annealing at 61°C and a 1 min extension at 72°C for 30 cycles. A positive reaction using both PCR conditions indicated the presence of L rhamnosus strain R0011.
Mild acute stress
On postnatal days 60–70, MS rats previously treated with vehicle or probiotics as neonates were subjected to WAS for 30 min. (We had previously shown that adult rats exposed to MS as neonates had exaggerated gut responses to WAS.15) Rats were placed on a platform in a container filled with warm water (25°C) to 2 cm below the platform. Following the stress period, rats were immediately killed by decapitation. Colonic segments were removed and placed in oxygenated Krebs for Ussing chamber studies; additional samples were obtained for bacterial cultures.
Statistics
Results are expressed as the mean (SE). Groups were compared using analysis of variance (ANOVA) with Newman–Keuls for post hoc comparisons. Differences of p<0.05 were considered to be significant.
RESULTS
MS causes colonic dysfunction and impairs host resistance
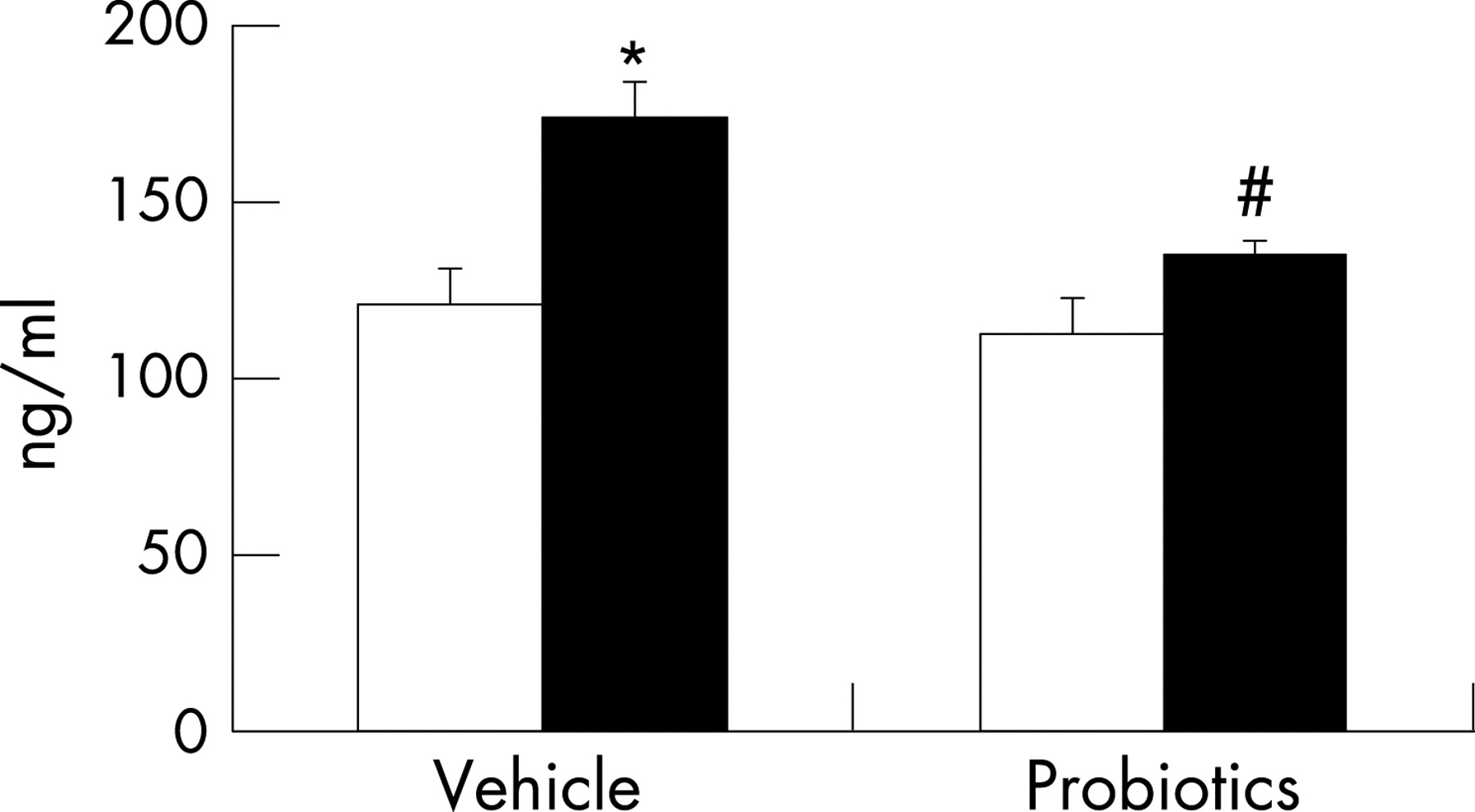
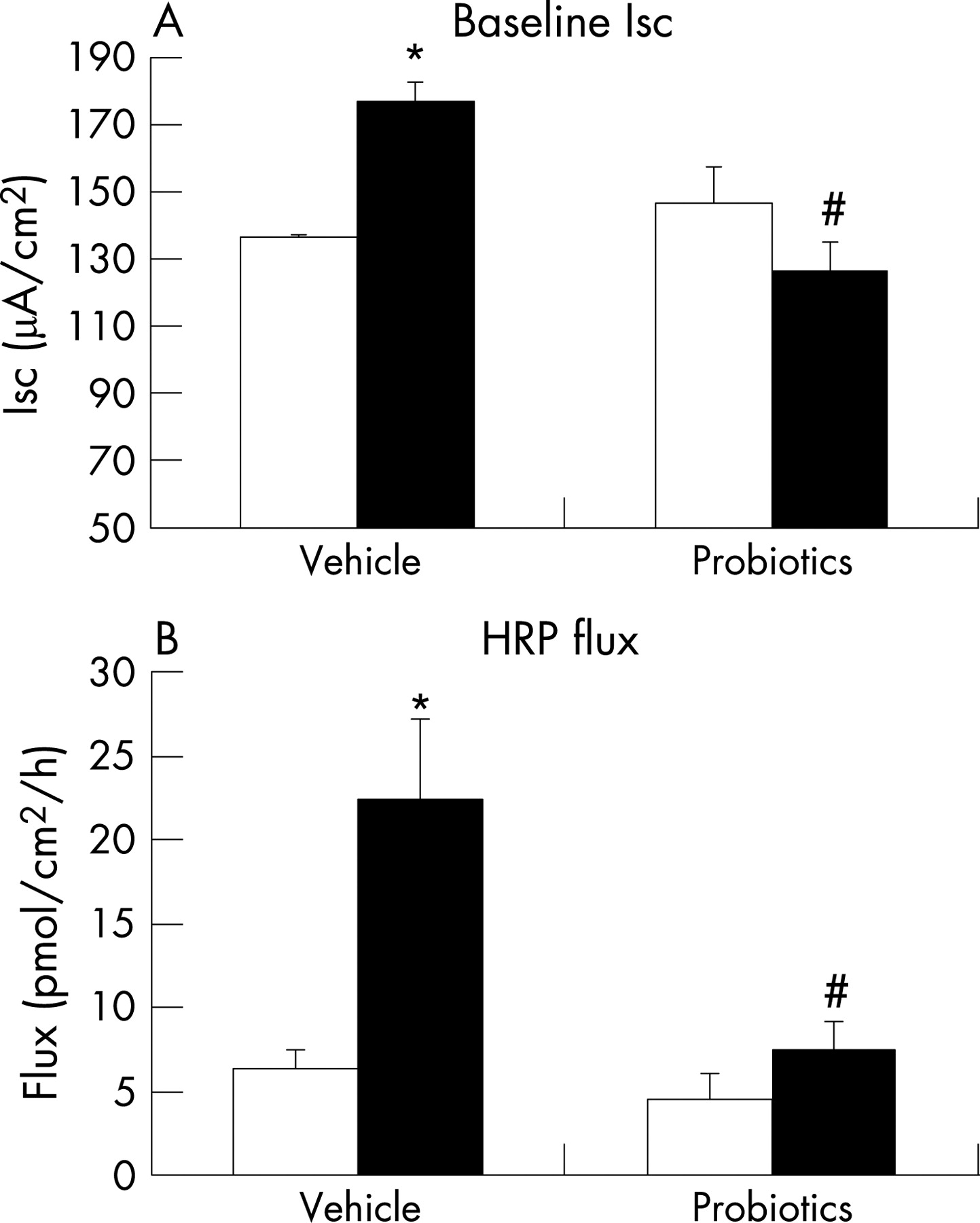
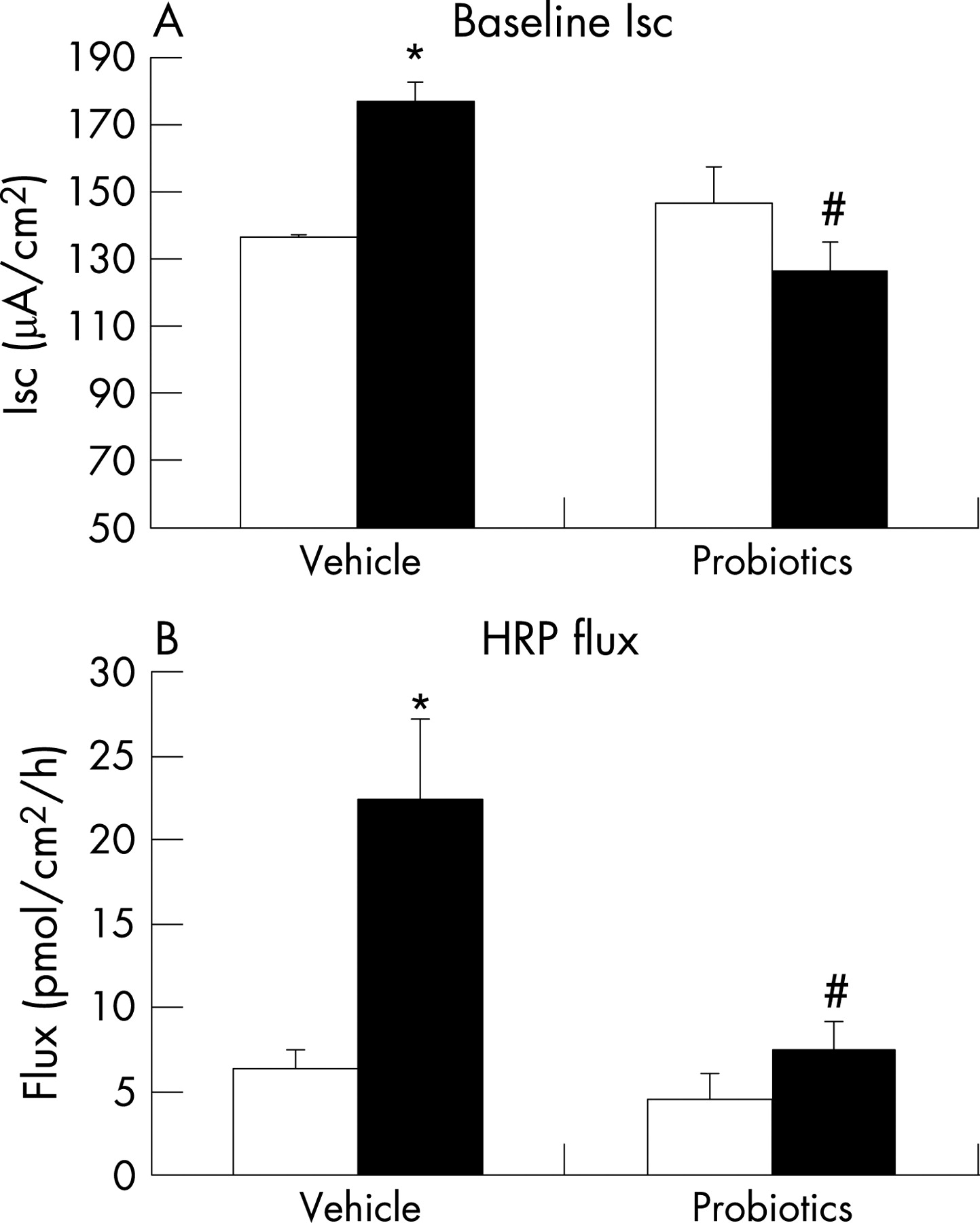
Similar to findings in our previous study,19 compared with NS rat pups, MS pups had elevated levels of circulating corticosterone (fig 1), suggesting enhanced activity of the HPA axis. With respect to colonic function, we confirmed earlier findings of increased baseline Isc and HRP flux (fig 2A and B), indicating altered mucosal barrier function. Adherent bacteria were dramatically increased in the colon of MS rats; in contrast, the number of Lactobacillus organisms was significantly reduced (table 2), implying altered colonisation resulting from early life stress.
MS results in increased cholinergic activity
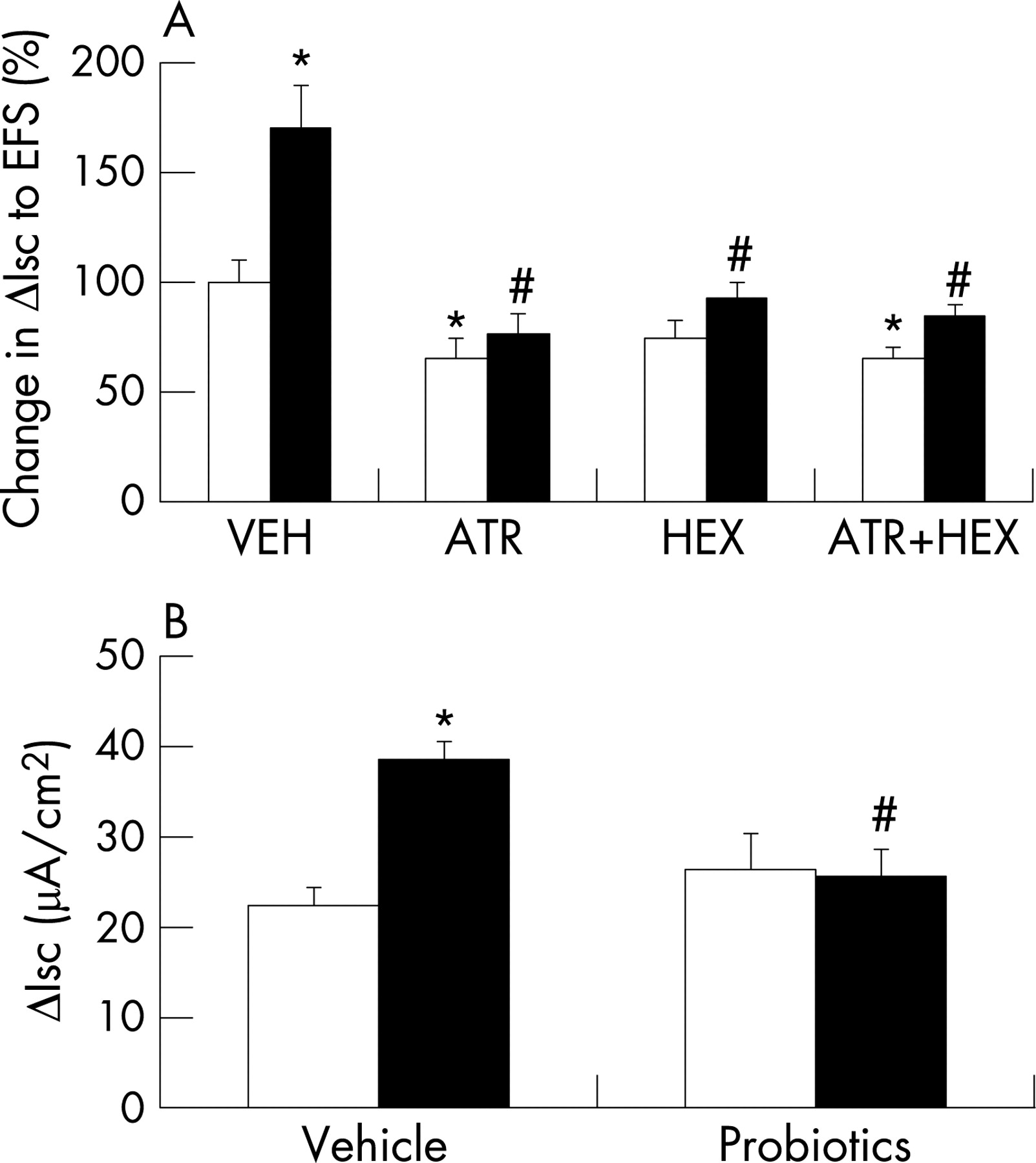
In addition to increased baseline Isc, we also documented a significantly enhanced ΔIsc to EFS in the colon of MS versus NS rat pups (fig 3A). However, the non-cholinergic components of these responses (after addition of atropine or hexamethonium) were not different in tissues from MS versus NS pups. Subtraction indicated that the cholinergic components were significantly greater in MS tissues (muscarinic: NS, 8.3 (2.6) μA/cm2, MS, 21.7 (6.5) μA/cm2, p<0.05, n = 7–8; nicotinic: NS, 2.2 (1.5) μA/cm2, MS, 8.7 (1.7) μA/cm2, p<0.05, n = 9–11; mean (SE)). The Isc response to bethanechol (in the presence of tetrodotoxin to eliminate neural involvement) was similar in tissues from both groups (NS, 57.2 (14.0) μA/cm2, MS, 49.5 (8.2) μA/cm2, n = 6–10). Taken together, these findings suggest that the abnormal response to EFS was not due to a change in the epithelium (receptors or secretory machinery), but rather to increased cholinergic activity in the colon of MS pups.
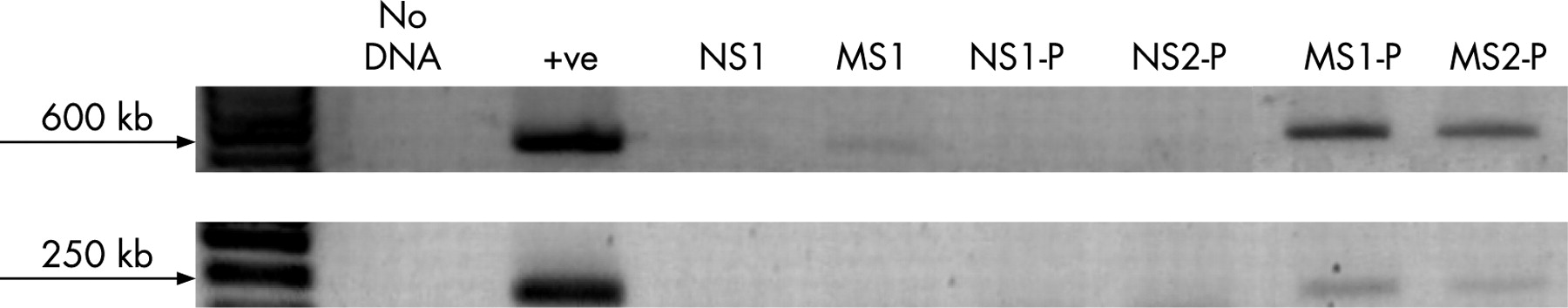
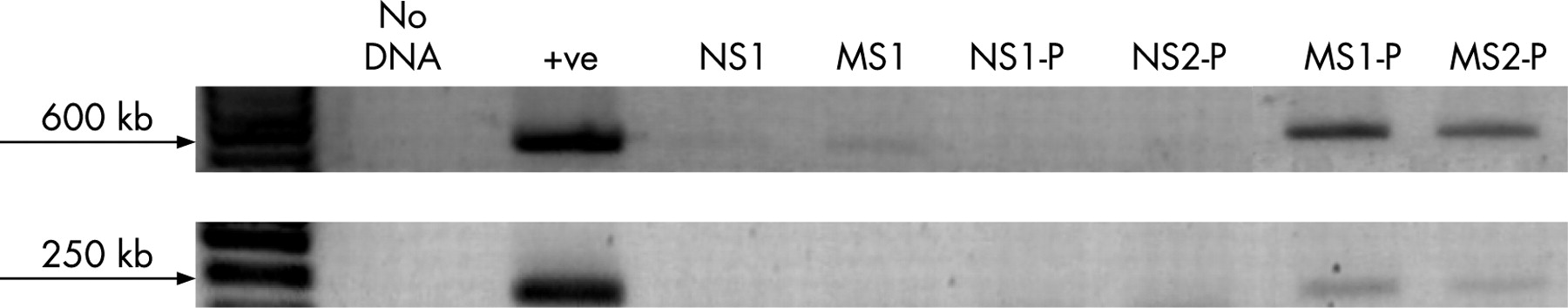
Administration of probiotic organisms induces their transient colonisation in MS pups
Since MS pups appeared to be deficient in Lactobacillus species in their gut flora, we administered probiotics in an attempt to restore beneficial bacteria to the microbiota. To confirm the transient colonisation of animals with the administered probiotic organisms, strain-specific PCRs were performed using faecal samples collected from MS and NS pups. MS and NS animals not administered probiotics served as negative controls, while DNA extracted from purified R0011 colonies was used as the positive control. Lactobacillus rhamnosus strain R0011 was present in faeces of MS pups, with minimal evidence of this organism in NS pups (fig 4). The absence of detection of R0011 in the faecal samples of NS pups suggests that the presence of normal numbers of Lactobacillus organisms in their gut flora prevented transient colonisation by the exogenously administered probiotic organisms.
Probiotics restore normal ion transport and macromolecular permeability in colon of MS pups
Daily administration of probiotics resulted in significant reduction in the elevated baseline Isc in the colon of MS animals, returning values to those found in NS control pups (fig 2A). Similarly, probiotics prevented the MS-induced increase in HRP flux (fig 2B). Finally, probiotics normalised the enhanced ΔIsc to EFS in MS pups (fig 3B). Probiotics had no effect on any physiological parameter in NS pups.
Probiotics prevent abnormal adherence/penetration of commensal organisms in the colon of MS pups
Administration of probiotics inhibited the MS-induced increase in total bacteria adhering to the colonic mucosa (NS, 78.3 (29.6) CFU(103)/g tissue; MS, 88.1 (23.0) CFU(103)/g tissue, n = 7–9) and restored numbers of Lactobacillus organisms (NS, 64.9 (36.4) CFU(103)/g tissue; MS, 60.1 (26.2) CFU(103)/g tissue, n = 7–8) to those observed in NS controls.
Probiotics ameliorate MS-induced enhanced activity of the HPA axis
We measured corticosterone as an indicator of HPA axis activation.30 We have previously shown that neonatal MS causes an increase in basal serum corticosterone levels, which is maximal at day 20 and persists until day 30 (10 days following completion of the MS protocol).19 Unexpectedly, treatment with probiotics significantly reduced the enhanced level of circulating corticosterone in MS pups to the control value (fig 1), suggesting normalised activity of the HPA axis. There was no effect of probiotics on the corticosterone value in NS pups.
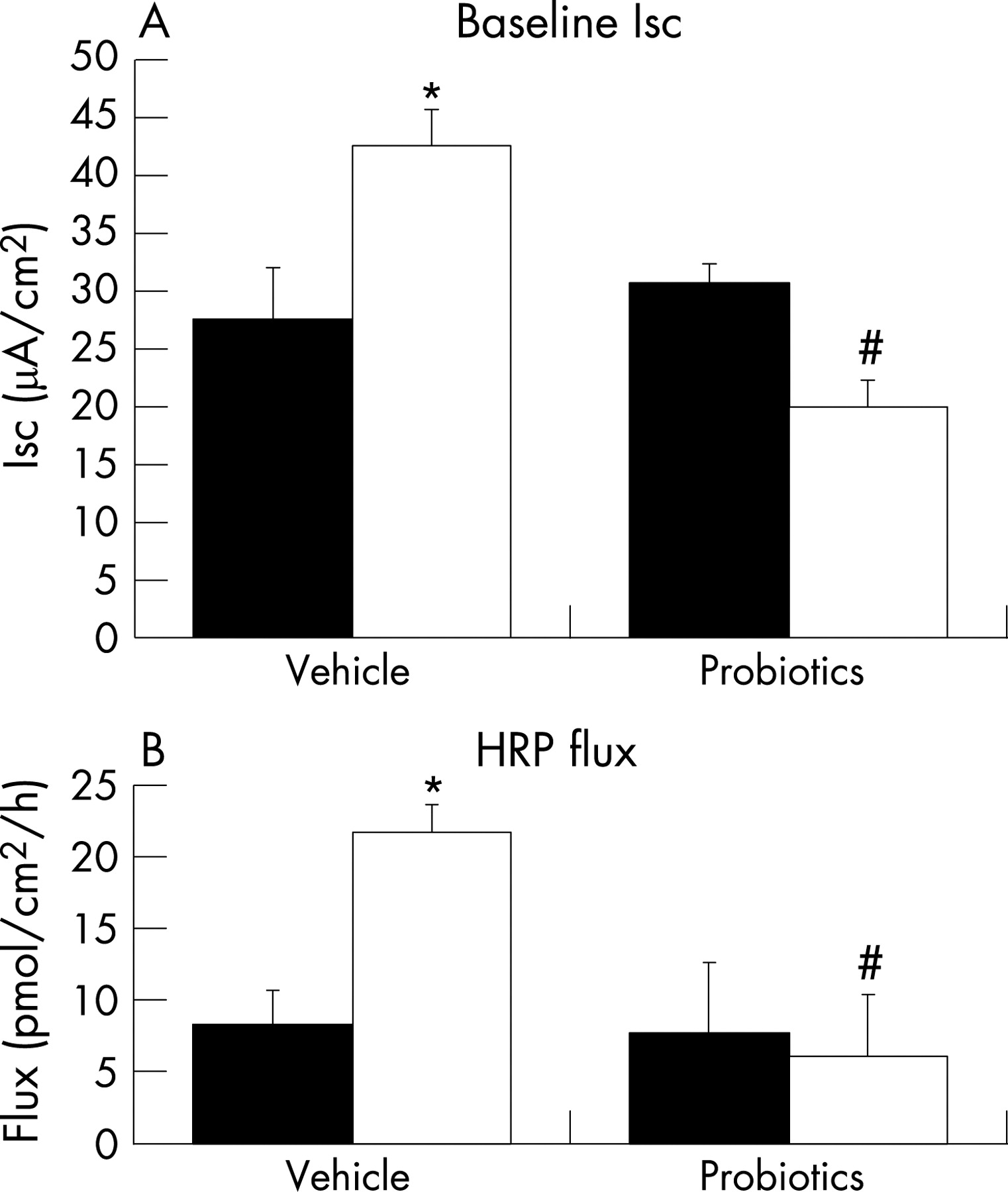
The beneficial effects of probiotics in the colon persist in older rats
We previously showed that neonatal rats exposed to MS develop an adult phenotype which is sensitive to psychological stress; although gut function is largely normal under baseline conditions, such rats display a stress-induced ion secretory state and a leaky barrier.15 Here, we found that administration of probiotics during early life significantly reduced the elevated Isc and HRP flux in 60- to 70-day-old MS rats exposed to WAS, but had no effect in similarly treated adult NS rats (fig 5). Furthermore, the stress-induced elevated bacterial adherence/penetration in the colon was reduced in MS rats treated as neonates with probiotics compared with vehicle controls (830 (267) CFU/g tissue vs 172 (105) CFU/g tissue; p<0.05, n = 7–8).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
DISCUSSION
Our study demonstrated that MS of neonatal rats disrupted normal bacterial colonisation of the gut, impaired colonic physiology and reduced host defence. Administration of probiotics normalised gut physiology and prevented bacterial attachment/penetration into the mucosa. Furthermore, probiotics ameliorated the enhanced HPA axis activity induced by MS. Our findings implicate inadequate colonisation with Lactobacillus species in both the colonic pathophysiology and the central nervous system changes resulting from exposure to early life stress.
Colonisation of the intestine with specific organisms is essential for the development of mucosal immunity, including the gut-associated lymphoid tissue, sIgA and T helper cell responses.10 Colonisation occurs in multiple steps, beginning at birth and continuing through weaning. Several factors, including infant feeding, environmental influences and genetics, are important regulators of colonisation. For example, exposure to prenatal acoustic stress altered bacterial colonisation of the gut in neonatal monkeys.31 In our rat model, we documented a fivefold decrease in CFU/g tissue of Lactobacillus species in the colon of MS animals compared with NS controls. Abnormal colonisation in MS pups was associated with barrier dysfunction, including decreased ion secretion resulting in a reduced driving force for water secretion to flush away noxious material from the mucosal surface, and increased permeability for macromolecules. In addition, the number of total bacteria adherent to the mucosa was significantly enhanced in MS pups compared with controls.
Epithelial ion transport and macromolecular permeability are tightly regulated by neurotransmitters originating from enteric nerves.32 33 Since acetylcholine is the dominant secretomotor neurotransmitter of the enteric nervous system which can directly stimulate epithelial cells to secrete chloride34 and increase transcellular permeability,32 we examined if MS affected cholinergic regulation. We found that the Isc response to EFS, an indication of the secretory response to enteric nerve stimulation, was increased in MS pups. The cholinergic component of the response was significantly greater in rat pups exposed to MS compared with NS controls, while the non-cholinergic component was similar. However, the non-neuronal (ie, in the presence of tetrodotoxin) Isc response to exogenous cholinergic agonist addition was unaffected by MS, suggesting that the abnormal EFS response in MS pups was related to changes in cholinergic nerves rather than epithelial cells. Our previous study19 implicated corticotrophin-releasing hormone (CRH) and activation of peripheral CRH receptors in MS-induced colonic barrier dysfunction. Other studies have provided evidence that stress-induced CRH can be released in the colon from extrinsic nerves35 36 or intrinsic nerves of the enteric nervous system,37 and CRH receptors have been demonstrated on enteric cholinergic neurons.38 Taken together, our studies suggest that MS stimulated CRH release which increased cholinergic activation of colonocytes, resulting in ion secretion and enhanced permeability. It is more difficult to explain the mechanism by which MS resulted in increased adherence of microbes to the mucosa. However, goblet cells also respond to acetylcholine with mucin secretion,24 and ongoing stimulation may deplete these cells of molecules which prevent close contact of microbes to the host mucosa. Therefore, it is possible that stress-induced release of CRH indirectly causes ion secretion, increased permeability and mucosal–bacterial interactions at least in part via enhanced cholinergic drive. It is likely that the reduced Lactobacillus component of the flora also played an important role in allowing other microbes to access the epithelial surface.
Since Lactobacillus species are important for normal colonic development,10 the supplementation of probiotic organisms was used to examine the potential benefit of such bacteria on barrier dysfunction. It has previously been shown that changing the composition of colonic bacterial flora modulates barrier function.39 Chronic stress in adult rats induced barrier dysfunction, including enhanced bacterial adhesion and penetration,40 which was reversed by pretreatment and co-treatment with probiotics.26 Garcia-Rodenas et al showed that MS-induced altered gut function was improved by feeding animals, after completion of the separation protocol, a diet containing probiotics.41 Taken together, these investigations suggest that specific commensal microflora are important in maintaining normal colonic physiology during times of stress. Therefore, we studied the effects of probiotics on the colonic pathophysiology in neonatal MS animals. Treatment with Lactobacillus organisms not only restored normal gut function and host defence in MS rat pups, but the beneficial effects persisted into adulthood. A previous study reported that similar probiotics administered to adult rats during exposure to chronic psychological stress did not completely restore normal barrier function, although they did inhibit enhanced translocation of commensal bacteria.26 In this study, administration of probiotics was performed in neonatal life. Colonisation during this time is key to maintaining appropriate barrier function, and early life stress interferes with this process. Therefore, it is likely that probiotic treatment during exposure of pups to MS is more effective in maintaining barrier function in neonates, and the beneficial effects persist until adulthood.
More remarkably, probiotics also prevented the development of MS-enhanced HPA axis activity indicated by elevated serum corticosterone levels. The interpretation of our results is somewhat limited by the assessment of corticosterone at one time point, although we have previously shown that this elevation resulting from MS is maximal at postnatal day 20 and persists until day 30, suggesting that the increase we observed is reliable. Stress triggers the release of CRH in the hypothalamus, causing adrenocorticotrophic hormone (ACTH) secretion from the pituitary gland and subsequently glucocorticoids from the adrenal glands. Disruptive events in early life, including MS42 43 or endotoxin exposure,44 can affect the HPA axis, since the neonatal period is critical in its development. IBS patients with a history of childhood abuse18 have enhanced HPA axis activity, indicated by elevated basal cortisol, as well as increased CRH-induced ACTH and cortisol responses compared with controls.45 Normal bacterial colonisation also affects the development of the HPA axis, shown by an exaggerated response to restraint stress in germ-free mice.20 In that study, the heightened ACTH and corticosterone responses were reversed by administration of a commensal organism, Bifidobacterium infantis, with the beneficial effects thought to be mediated by a neural mechanism. Taken together, these findings suggest that probiotic organisms can positively influence the dysregulated HPA axis resulting from early life stress. Further studies are needed to provide a more complete understanding of how probiotics affect the HPA axis.
What is the signalling that informs the brain regarding flora in the gut lumen? Our study did not address this, but we can speculate. Several studies indicate that enterochromaffin cells (which may also contain CRH46) are affected by enteric flora.47 48 Furthermore, neuroendocrine mediators from enterochromaffin cells can activate afferent nerves that project to central structures with connections to the HPA axis.49 Other studies support this hypothesis, including findings that signals can travel through afferent nerves in the vagus/pelvic nerve.35 36 This pathway may participate, at least in part, in mediating the HPA axis dysregulation in response to early life stress. Further studies in progress are addressing this issue.
In conclusion, our study suggests that MS-induced colonic pathophysiology was mediated, at least in part, by impaired colonisation with Lactobacillus bacteria, since normal function was restored by probiotic supplementation with these organisms. Furthermore, probiotic treatment significantly reduced the enhanced HPA axis activity due to MS, effects probably mediated in part by enteric nerves. This study reveals the benefit of probiotic supplementation during periods of neonatal stress as a means of preventing stress-induced effects on colonic physiology in neonates and adult rats. This study also highlights the importance of the neonatal period in determining the responsiveness to stress later in life.
Acknowledgments
The authors would like to thank Dr Thomas Tompkins of Institut Rosell-Lallemand Inc. for providing the probiotic organisms, and Dr Karen Hagen from Institut Rosell-Lallemand Inc. for providing the primers for the PCRs.
REFERENCES
Footnotes
Funding: This research was supported by grants from the Canadian Institutes for Health Research and the Crohn’s and Colitis Foundation of Canada.
Competing interests: None.
- Abbreviations:
- ACTH
- adrenocorticotrophic hormone
- CFU
- colony-forming units
- CRH
- corticotrophin-releasing hormone
- EFS
- electrical field stimulation
- HPA axis
- hypothalamus–pituitary–adrenal axis
- HRP
- horseradish peroxidase
- IBD
- inflammatory bowel disease
- IBS
- irritable bowel syndrome
- IL
- interleukin
- Isc
- short-circuit current
- MS
- maternal separation
- NS
- non-separation
- sIgA
- secretory immunoglobulin A
- WAS
- water avoidance stress
Linked Articles
- Corrections