Article Text

Abstract
Objective: Intestinal infection evokes hypersensitivity in a subgroup of patients with irritable bowel syndrome (IBS) long after healing of the initial injury. Trinitrobenzene sulfonic acid (TNBS)-induced colitis in rodents likewise results in delayed maintained hypersensitivity, regarded as a model of some aspects of IBS. The colon and rectum have a complex sensory innervation, comprising five classes of mechanosensitive afferents in the splanchnic and pelvic nerves. Their plasticity may hold the key to underlying mechanisms in IBS. Our aim was therefore to determine the contribution of each afferent class in each pathway towards post-inflammatory visceral hypersensitivity.
Design: TNBS was administered rectally and mice were studied after 7 (acute) or 28 (recovery) days. In vitro preparations of mouse colorectum with attached pelvic or splanchnic nerves were used to examine the mechanosensitivity of individual colonic afferents.
Results: Mild inflammation of the colon was evident acutely which was absent at the recovery stage. TNBS treatment did not alter proportions of the five afferent classes between treatment groups. In pelvic afferents little or no difference in response to mechanical stimuli was apparent in any class between control and acute mice. However, major increases in mechanosensitivity were recorded from serosal afferents in mice after recovery, while responses from other subtypes were unchanged. Both serosal and mesenteric splanchnic afferents were hypersensitive at both acute and recovery stages.
Conclusions: Colonic afferents with high mechanosensory thresholds contribute to inflammatory hypersensitivity, but not those with low thresholds. Pelvic afferents become involved mainly following recovery from inflammation, whereas splanchnic afferents are implicated during both inflammation and recovery.
Statistics from Altmetric.com
A subset of patients develop irritable bowel syndrome (IBS) symptoms following acute gastrointestinal infection.1 2 3 These patients, termed post-infectious IBS (PI-IBS), continue to suffer pain and discomfort despite recovery of overt intestinal damage. In all patients with IBS, hypersensitivity to colorectal mechanical stimuli – either smooth muscle contraction, luminal contact, or gut distension – is a characteristic feature.4 5
Experimentally induced colonic damage might provide a model for PI-IBS whereby acute damage induces hypersensitivity to colorectal distension which is maintained following recovery from the initial insult. The acute stage of trinitrobenzene sulfonic acid (TNBS) colitis is well characterised, with a predominantly Th1 immune driven initial insult resulting in structural damage that is maximal 3 days post-treatment, still evident 1 week later, and healed 2 weeks post-treatment.6 7 8 9 This is associated with visceral hypersensitivity that is initially present at least 3 days post-treatment and maintained 1 week later but is resolved 2 weeks post-TNBS.6 8 At later stages, however, the hypersensitivity returns, and can be greater than that in the acute stage.10
The mechanism of visceral hypersensitivity in IBS, or in the TNBS model, is not known. Mechanosensory function of primary afferent endings in the colon is a crucial step in the transmission of painful stimuli to the central nervous system. How the sensory innervation of the colon is altered in the acute and chronic stages after an insult is therefore important to determine, in order to understand which types of sensory fibres along which pathways are targets for treatments to reduce symptoms of PI-IBS.
The colon transmits a complex array of sensory information to the central nervous system via two separate nerve supplies: the splanchnic and pelvic nerves, whose sensory function is particularly well characterised in the mouse compared to other species.11 Briefly, the splanchnic pathway consists predominantly of afferent endings responding to distortion of serosal or mesenteric blood vessels that do not respond either to mucosal contact or low intensity smooth muscle stretch, but may respond to high intensities of stretch.11 These have been characterised anatomically as varicose branching axons in close proximity to blood vessels.12 In contrast, four classes of pelvic afferents exist, three of which respond to fine mucosal stroking and/or low intensity stretch, in addition to serosal afferents like those in the splanchnic pathway.11 Splanchnic afferents respond much more frequently to nociceptive and inflammatory mediators.13 14 It follows, therefore, that the splanchnic pathway is tuned more toward detection of noxious events when compared to the pelvic pathway.
An important unanswered question is which aspects of the rich diversity of mechanosensory information from the colon are altered by inflammation, and therefore which populations of colonic sensory fibres are involved in post-inflammatory hypersensitivity. These may constitute a specific target for alleviation of symptoms in post-infectious IBS. Therefore, in this study we aimed to characterise the alterations in colonic afferent mechanosensitivity in both acute and post-inflammatory states following TNBS colitis and determine specific subpopulations of afferents involved.
Methods
Induction of colitis
Male C57 BL/6 mice were used. TNBS-treated mice were 12 weeks old at time of treatment. Control mice were 13 or 16 weeks old at the time of experimentation. Two separate experimental groups were established, one for histological scoring of extent of inflammation, the other for macroscopic scoring and electrophysiological experiments. Mice were fasted overnight but with access to nutrition from 5% glucose solution. Colitis was induced by singular intrarectal administration of 0.1 ml trinitrobenzene sulfonic acid (TNBS) (130 μg/ml in 30% EtOH) via a polyethylene catheter inserted 3 cm from the anus of isoflurane-anaesthetised mice.
Histological assessment of colitis
Mice were humanely killed (CO2 inhalation and cervical dislocation) at day 0 (control), 3 (d3), 7 (acute) and 28 (recovery) days post-treatment. The distal 3 cm of colon was removed following trans-cardial perfusion with 4% paraformaldyhyde in 0.1 mol/l phosphate-buffered saline (PBS) and post-fixed overnight at 4°C. Paraffin sections (6 μm ) were cut and stained with haematoxylin and eosin. Microscopic assessment was then blindly scored according to the following:15 mucosal architecture (0–3, normal to extensive damage), cellular infiltrate (0–3, no infiltrate to transmural infiltration), muscle thickening (0–3, normal to extensive thickening), crypt abcesses (0 absent, 1 present), goblet cell depletion (0 absent, 1 present). A maximum possible total is 11.15 A blinded scorer assessed 10 sections per mouse, and the means of the measurements were calculated for each mouse and each treatment.
Macroscopic assessment of colitis
Immediately prior to colonic electrophysiology experiments, in colons at days 0, 7 and 28 the extent of inflammation was blindly scored macroscopically by two scorers according to the following:15 0, no damage; 1, hyperaemia without ulcers; 2, hyperaemia and thickening of the bowel wall without ulcers; 3, one site of ulceration without bowel wall thickening; 4, ⩾2 sites of ulceration; 5, 0.5 cm of inflammation and major damage; 6–10, 1 cm of major damage, with score increased by 1 for every 0.5 cm of damage observed to a maximum of 10. Added to this score were: 0 or 1 for the absence or presence of diarrhoea; 0 or 1 for the absence or presence of stricture; 0, 1 or 2 for the extent of adhesions. The total maximum score is 13.
Colonic electrophysiological recordings
Nerve dissection
Mice were humanely killed at days 0 (control), 7 (acute) and 28 (recovery) post-TNBS. The colon and rectum (5–6 cm) and attached mesentery were removed and afferent recordings made from pelvic and splanchnic nerves in specially constructed organ baths, as described in detail previously11 13 14 16
Characterisation of colonic afferent properties
Receptive fields were identified by systematically stroking the mucosal surface and mesenteric attachment with a stiff brush. Once identified, receptive fields were assessed with three distinct stimuli to enable classification: focal compression of the receptive field with calibrated von Frey hairs (70, 160, 400, 600, 1000, 1400 and 2000 mg; each force applied five times for a period of 3 s with a 10 s interval), mucosal stroking with calibrated von Frey hairs (10, 200, 500 and 1000 mg force; each force applied 10 times) and circular stretch (normally 1–5 g in 1 g increments; each weight applied for a period of 1 min with a 1 min interval between each application). In some experiments 1–11 g was used to determine the responsiveness of afferents to higher intensity stretch. Stretch was applied as described previously.11 13 14 16 Categorisation of afferent properties was accorded based on previously published classification11 16 and consisted of mesenteric, serosal (responding to focal compression but not low intensity stroking or low intensity circular stretch), mucosal (responding to fine mucosal stroking; 10 mg von Frey hair), muscular (responding to low intensity circular stretch <5 g, but not fine mucosal stroking) or muscular/mucosal (responding to both low intensity stretch and fine mucosal stoking).
Data recording and analysis
Electrical signals generated by nerve fibres were amplified, filtered, digitised and stored as described previously.11 13 14 16 Action potentials were discriminated as single units on the basis of waveform, amplitude and duration. Data are expressed as mean with the SEM, and n is the number of afferents. Data were analysed using either one or two way analysis of variance (ANOVA). Differences were considered significant at p<0.05. Stimulus–response functions were constructed by assessing the mean number of action potentials per second (spikes/second) generated in response to mechanical stimuli.
Results
Macroscopic and histological examination of colon
Significant damage was present in acute inflammatory mice when compared to control mice (p<0.05, fig 1A,B), indicating mild inflammation. Macroscopic scoring was not significantly different between control and recovery mice (p>0.05) indicating healing from the inflammatory event. Histological examination of TNBS treated colons demonstrated significant damage was induced by day 3 of treatment that had largely recovered by day 7 (acute) and was negligible by day 28 (recovery) (fig 1A,C).
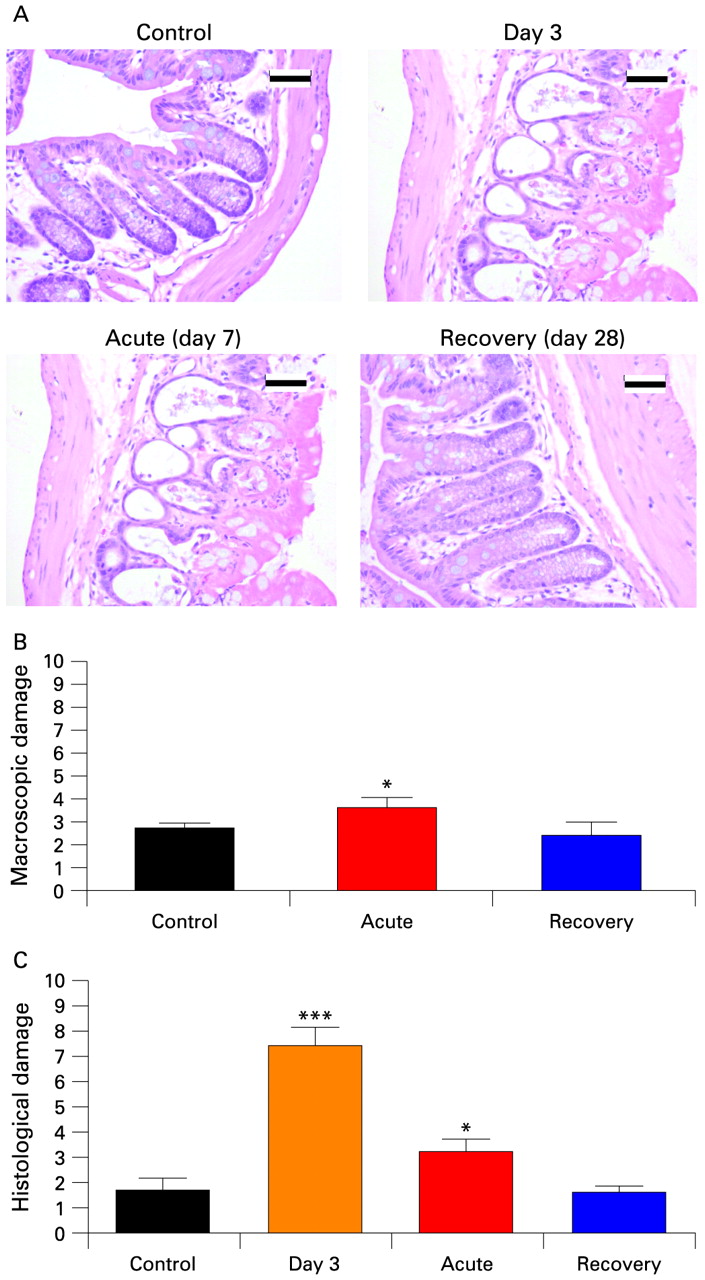
Assessment of trinitrobenzene sulfonic acid (TNBS) colitis induced inflammation. (A) Representative examples of the most damaged sections of colon in control and TNBS treated mice at day 3, acute (day 7) and recovery (day 28). (B) Macroscopic assessment indicating a small but significant (*p<0.05, unpaired t test, n = 5) difference in appearance of the colon between control and acute stages, which had normalised by the recovery stage. Degree of damage was scored according to degree of hyperaemia, ulceration and presence of diarrhoea, stricture or adhesions for a total score of 13. (C) Histological scoring indicating damage induced by day 3 has repaired by day 7 and recovered by day 28. Degree of inflammation scored according to appearance of mucosa and musculature, and the presence of cellular infiltrate, crypt abscesses and goblet cells with a maximum total of 11. *p<0.05 vs Control; ***p<0.001 vs Control. Scale bar, 20 μm.
Colonic afferents
Population findings
TNBS-induced colitis did not affect the proportion or distribution of pelvic or splanchnic afferent subclasses that we encountered during nerve recording protocols in preparations from either acute or recovered mice. This indicates that afferent endings were unlikely to be present in different layers of gut than in controls. This was assessed by recording the type of afferent observed in each electrophysiological experiment and plotting the location of its main receptive field (fig 2). Thus, four classes of fibres were recorded in each pathway with distinct mechanosensory properties, as we have observed previously:11 in the pelvic nerve, serosal, muscular, mucosal and muscular/mucosal afferents; in the splanchnic nerve, serosal, muscular, mucosal and mesenteric afferents. Whilst extraluminal (serosal and mesenteric) afferents predominate and others are rare in the splanchnic innervation, there is more of a balance of intra- and extraluminal afferents in the pelvic innervation. TNBS treatment did not alter the total number of afferents identified in each experiment. An average of 4.0 (lumbar splanchnic nerve (LSN)) and 4.7 (pelvic nerve (PN)) individual afferent endings were recorded in each control experiment, 3.9 (LSN) and 6.0 (PN) in each experiment on acute animals and 3.8 (LSN) and 4.3 (PN) afferents per mouse after recovery. This would suggest that recruitment of afferent fibres that were normally silent was unlikely. Spontaneous discharge was rarely observed in any class of pelvic or splanchnic afferents, as previously reported,11 and no induction of activity occurred in acute or post-inflammatory preparations.
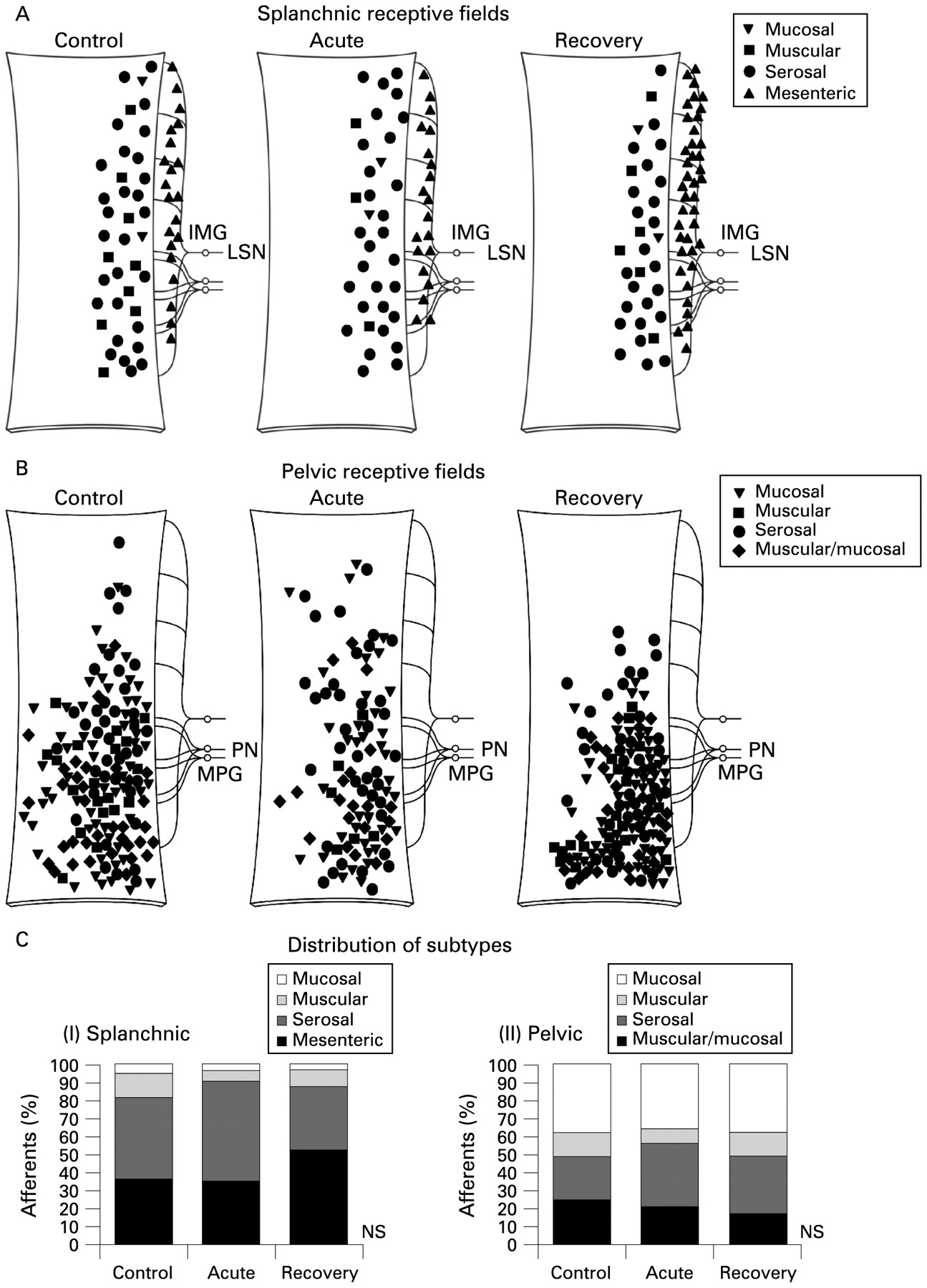
Distribution and proportions of colonic afferent subclasses. Location of receptive fields of (A) splanchnic and (B) pelvic afferents encountered during experiments at control, acute and recovery states. Preparations are 5–6 cm in length, beginning directly adjacent to the anal canal. Proximal is shown uppermost. Subtypes are denoted by specific symbols; the same ones are used in subsequent figures. (C) The frequency of occurrence of each subtype plotted as a percentage of the total number. NS = none of the proportions were significantly different between stages of inflammation for (1) splanchnic or (2) pelvic afferents (p>0.05, χ2 test). IMG, inferior mesenteric ganglion; LSN, lumbar splanchnic nerve; MPG, major pelvic ganglion; PN, pelvic nerve.
Mechanical sensitivity: pelvic afferents
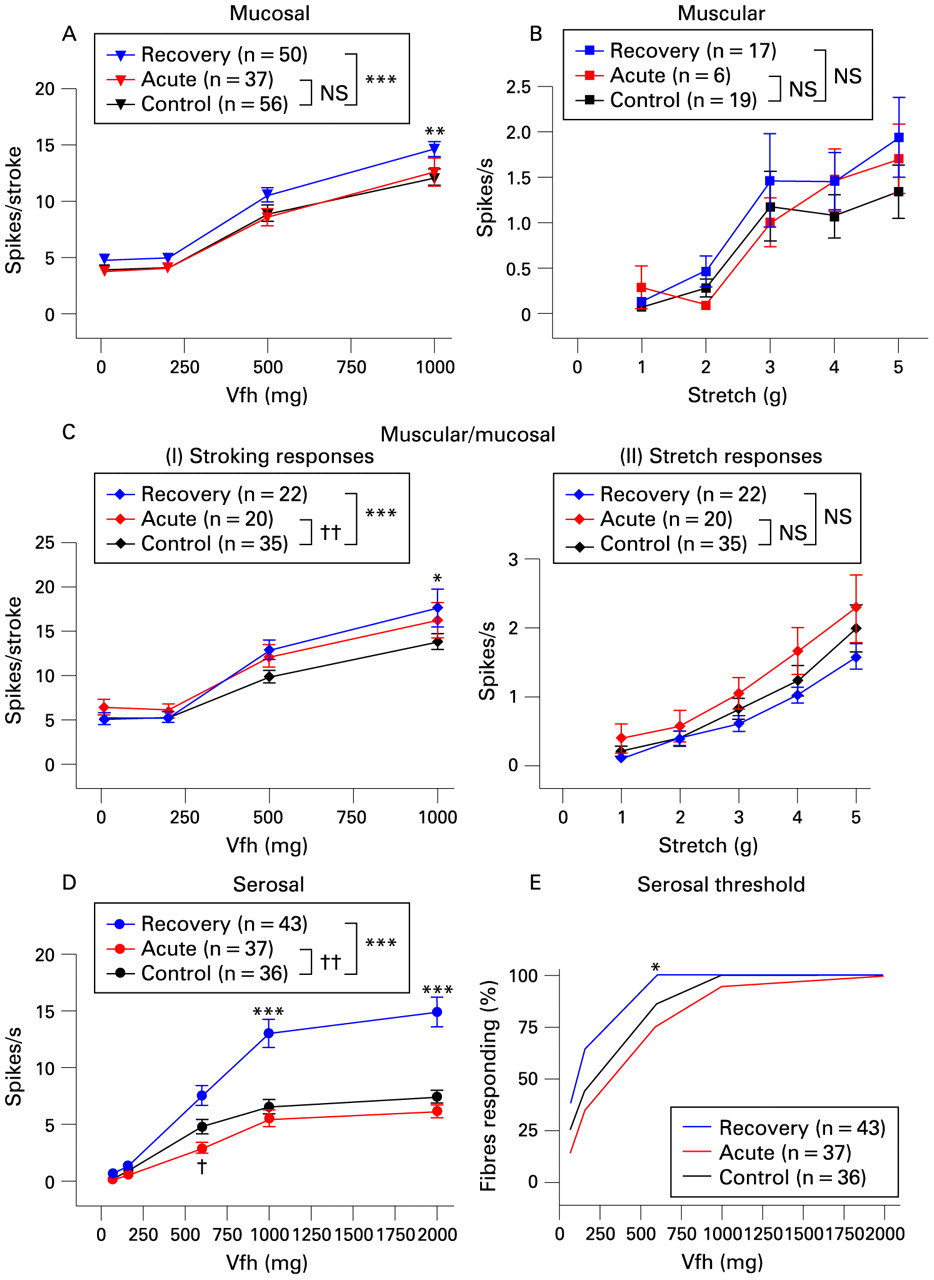
Changes in pelvic afferent mechanical sensitivity were minor or absent in the acute group when compared to controls (red vs black in fig 3). A slight, but significant, decrease was seen in the stimulus–response function of serosal afferents, with an individual effect significant only at 600 mg force (fig 3D).Their mean thresholds to von Frey probing were unaffected in the acute stage (437 (SEM 54) mg control (n = 36); 584 (SEM 77) mg acute (n = 37)). A small change was evident in the mucosal stroking response of muscular/mucosal afferents (fig 3Ci), with an increase in the curve. Responses to muscular stretch, both in muscular afferents and muscular/mucosal afferents, were unchanged (fig 3 B,Cii), as were responses of mucosal afferents to stroking (fig 3A).

Mechanosensitivity of pelvic colonic afferents. Stimulus–response functions of (A) mucosal, (B) muscular, (C) muscular/mucosal and (D) serosal afferents, with responses of muscular/mucosal afferents shown to both mucosal stroking (Ci) and circular stretch (Cii). Significance of two-way ANOVA comparison between control and each stage is shown (***p<0.001 recovery vs control; ††p<0.01 acute vs control) Significance of Bonferroni post-hoc tests is shown above or below individual points (***p<0.001 recovery vs control, †p<0.05 acute vs control). Differences between thresholds of serosal afferents (E) are shown only when significant (*p<0.05 recovery vs control, Fisher’s exact test). Vfh, von Frey hair.
In contrast to the minor changes seen in the acute phase, after recovery from inflammation, major increases in mechanosensitivity were observed (blue vs black in fig 3). These increases were most evident in serosal afferents, with a 2-fold increase in the stimulus–response function (fig 3D). In addition, these fibres showed a decrease in von Frey threshold, significant p<0.05) at 600 mg force (fig 3E). The average probing activation threshold was also reduced in recovery (control 437 (SEM 54) mg vs recovery 280 (SEM 39) mg, p<0.05 t test). Increases in mucosal afferent mechanosensitivity were also seen, but were more moderate, yet still significant (maximal 21% increase, fig 3A), as was the increase in the responses to mucosal stroking in muscular/mucosal afferents (maximal 28% increase, fig 3Ci)). The stretch responses of both muscular and muscular/mucosal afferents were unchanged in recovered mice when compared to control mice (fig 3B,Cii).
Mechanical sensitivity: splanchnic afferents
The pattern of changes we observed in splanchnic afferents differed markedly from those in pelvic afferents. In the two predominant classes of splanchnic afferents we encountered, large increases in mechanosensitivity were seen at both acute and recovery stages (fig 4A,B). Accompanying the increase in mechanosensory responses were decreased von Frey thresholds (fig 4C,D). In particular, the mean probing activation thresholds of serosal afferents (control 638 (SEM 108) mg) were significantly reduced in acute (308 (SEM 36) mg, p<0.05) and recovery states (270 (SEM 46) mg, p<0.05).

Mechanosensitivity of splanchnic colonic afferents to von Frey hair probing. Stimulus–response functions of (A) serosal and (B) mesenteric afferents. Significance of two-way ANOVA comparison between control and each stage is shown (***p<0.001 recovery vs control; †p<0.05 acute vs control †††p<0.001 acute vs control). Significance of Bonferroni post-hoc tests is shown above or below individual points (***p<0.001 recovery vs control, **p<0.01 recovery vs control, †††p<0.001 acute vs control, †p<0.05 acute vs control). Differences between thresholds of serosal (C) and mesenteric (D) afferents are shown only when significant (*p<0.05 recovery vs control, ††p<0.01 acute vs control, Fisher’s exact test). Vfh, von Frey hair.
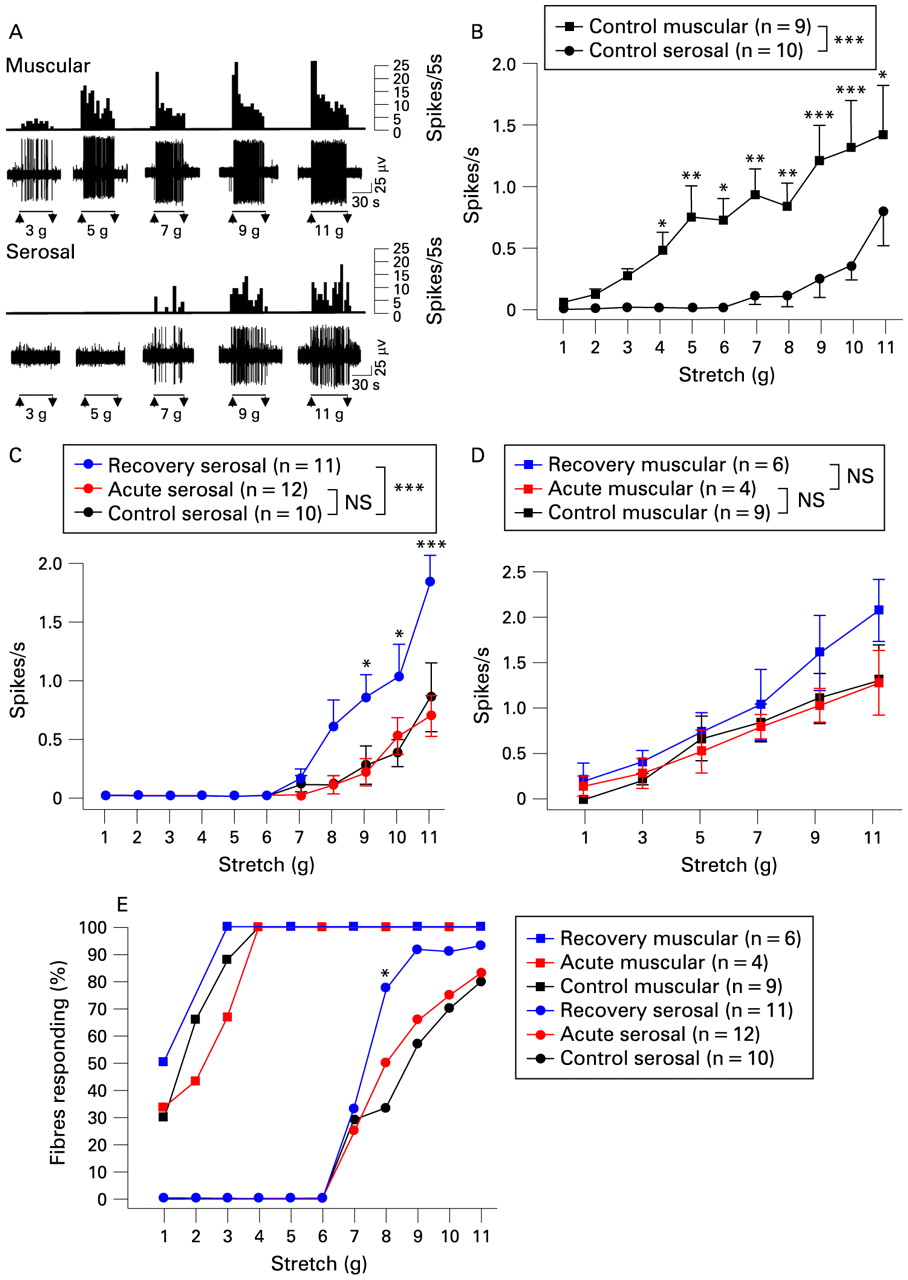
Although von Frey probes provide highly reproducible stimuli necessary for accurate quantification of mechanosensory responses, they do not relate directly to the situation that would be encountered during contraction and distension in vivo. We hypothesised that increases in responses of splanchnic extraluminal (serosal and mesenteric) afferents would translate to increased responses to distension, which are observed in the clinical situation.4 5 17 We addressed this by first comparing the responsiveness to circular stretch of serosal afferents with that of muscular afferents (which were encountered rarely in splanchnic nerve recordings). It was clear that muscular afferents respond at low thresholds (<3 g stretch, mean stretch activation threshold: 2.11 (SEM 0.30) g, fig 5A,B), with 100% of them responding at 4 g stretch (fig 5E). Serosal afferents, on the other hand, showed no response until at least 7 g stretch (fig 5A,B, mean stretch activation threshold: 9.22 (SEM 0.46) g), and were not all recruited even at 11 g stretch (fig 5B,E). In separate preparations that were maintained as a tube instead of opening as a sheet (n = 8), the threshold to balloon distension was reached at a pressure of 53 (SEM 4) mm Hg applied with the centre of the balloon as close as possible to the position of the receptive field. Similar thresholds were found for mesenteric afferents (56 (SEM 4) mm Hg, n = 4) even though their receptive fields were clearly away from the colonic wall, probably indicating that longitudinal forces are transferred through the mesenteric attachments.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Distension sensitivity of splanchnic colonic afferents. (A) Representative traces of muscular and serosal afferent responses to graded stretch across the receptive field, shown in grams, between each pair of arrows. In each case upper traces show integrated records of afferent discharge in spikes/5 s of the afferent fibre of which the raw electrical signal is show on the lower traces. (B) Group data showing stimulus–response functions of splanchnic muscular (n = 9) and serosal afferents (n = 10) to low, through to high intensity stretch. The difference between these curves was significant when assessed with two-way ANOVA (p<0.001). (C) Serosal afferent stimulus–response functions to stretch in controls (n = 10), acute (n = 12) and recovery (n = 11) stages of inflammation, compared at increments of 1 g. (D) Muscular afferent responses to stretch at each stage of inflammation, compared at increments of 2 g (n = 9 (control), n = 4 (acute), n = 6 (recovery) respectively). Differences between curves, assessed by two-way ANOVA, are show in the legend (***p<0.001), and differences at individual stimulus strengths, assessed by Bonferroni post-hoc tests, are shown next to each point (***p<0.001, **p<0.01, *p<0.05). (E) Thresholds of muscular and serosal afferents to stretch at each stage, shown as percentage of the total number fibres recorded responding, with no significant differences between threshold of muscular afferents at each stage, but with a significant increase in the proportion of serosal afferents responding at 8 g after recovery (*p<0.05, Fisher’s exact test).
We then compared responses to stretch after acute inflammation. Interestingly, no acute increase was seen in serosal afferent responses to stretch (fig 5C), whereas they showed clear increases in responses to von Frey hair stimuli (fig 4A), suggesting that more intense focal stimuli reveal changes more readily than evenly distributed stimuli at lower intensities. After recovery from inflammation, both distension and von Frey responses were significantly augmented, such that maximal amplitude of distension responses were comparable with those of muscular afferents (fig 5C,D). Despite this increased responsiveness, the threshold of serosal afferents to stretch was still 7 g or above (mean stretch activation threshold: control: 9.22 (SEM 0.46) g vs recovery: 8.27 (SEM 0.40) g), although significantly more of them responded at 8 g in recovery states (fig 5E). Muscular afferent responsiveness (fig 5D) and threshold to stretch (fig 5E) were unchanged in both acute and recovery stages, similar to what we observed in pelvic muscular afferents (fig 3B). Mucosal afferents were too rare in all cases for any statistical comparison to be made.
Discussion
We have demonstrated clear differences between the roles of the two colonic sensory pathways in the way they are sensitised by a single inflammatory event. These differences relate to the afferent subpopulations affected and the time course of sensitisation. There was a predominant involvement of splanchnic afferents in visceral hypersensitivity 7 days after TNBS treatment, whereas pelvic afferent mechanosensitivity was mainly unaffected in all afferent classes at this stage. However, a contribution is made by both splanchnic and pelvic afferent pathways to delayed post-inflammatory hypersensitivity. Although clear decreases in threshold and increases in maximal responses were evident, no significant change in the proportions of subtypes was observed, suggesting afferents did not change their phenotype, although there may be a trend towards an increase in the occurrence of mesenteric afferents after recovery. We did not see induction of spontaneous discharge, indicating no major increase in excitability at the endings of sensory neurones. Importantly, no evidence of increased yield of afferent recordings was seen, suggesting that previously insensitive, or “silent” afferents18 were not recruited by inflammation. Post-inflammatory hypersensitivity can therefore be explained based on existing populations of afferents.
Several groups have investigated changes in afferent sensitivity over different time courses: after direct application of individual or multiple inflammatory mediators, during acute inflammation or infection, and after recovery of mucosal lesions. These changes have been studied in vitro and in vivo, in different regions of gut, from different pathways, using a variety of species and a variety of infections or inflammatory agents. They have also been conducted using different recording methods: from single fibres, from multi-unit strands, and from cell bodies in the dorsal root or nodose ganglia after retrograde labelling. There is consensus that inflammation does change afferent function, but there are discrepancies in the details. It is interesting to compare the findings of the current study, in which several subtypes in two pathways were studied, with those already published where individual subtypes were investigated or general observations on mechanosensory function were made. From this comparison it becomes evident that many of the apparent discrepancies in the literature may be explained by the concept of “different subtypes, different pathways and different time courses”. For simplicity and comparability, the discussion here is restricted to studies of colorectal afferents, and their changes after mucosal inflammation, but it is clear that much has been learned from studies of small intestinal and gastric afferents,19 20 and studies of direct effects of inflammatory mediators,13 14 21 22 all of which can probably be extrapolated to apply to colorectal afferents during and after the full process of inflammation.
Our previous study in the rat focussed on changes in chemosensitivity of splanchnic serosal and mesenteric afferents after dextran sodium sulfate (DSS) colitis.23 We saw no changes in mechanosensitivity in the rat model at either acute or recovery stages, whereas responsiveness to serotonin was markedly augmented. This suggests that in order to observe upregulation of mechanosensory mechanisms, the choice of species and/or the inflammatory agent is important. Our recent study in the guinea-pig24 showed, in a TNBS model, that rectal muscular afferents were sensitised to distension in the acute phase of inflammation, but unaffected after recovery. In this case our stimulus is the same but the species is different, but there is agreement that long-term changes in mechanosensitivity are absent in low-threshold distension-sensitive subtypes of afferents.
The majority of studies in the literature have focussed on the responses of afferents to colorectal distension, and how this is altered by inflammation. A recent in vivo recording study25 showed that acute TNBS colitis in rats increased the response to colorectal distension of pelvic afferent C-fibres, but had no effect on A-delta-fibres, suggesting differences between subtypes. However, all afferents in mouse colon are C-fibres,16 indicating that differences we observed are not linked with conduction velocity. Another study of rat TNBS colitis, where recordings were performed in vitro, confirmed hypersensitivity to distension, indicating differences are not linked to the setting of recordings.26 An earlier study27 using an identical in vivo rat TNBS protocol, recording from the same population of afferents, did not reveal any changes in mechanosensitivity, so clearly there is controversy in this area. In agreement with the latter study, and later observations in a mouse zymosan colitis model,21 we saw no change in low-threshold distension-sensitive afferents in the pelvic nerve. We can add that no change occurs either in splanchnic low-threshold distension-sensitive (muscular) afferents. The pattern that seems to be emerging from these and our studies is one of inconsistent or absent effects of inflammation on low-threshold distension-sensitive subtypes of afferents. Importantly, what we have revealed are much more consistent effects on high-threshold afferents, and a predominant contribution of splanchnic afferents towards hypersensitivity, neither of which have previously been investigated. Our data on the sensitivity to colorectal distension of serosal and mesenteric afferents indicate that although these fibres do not reduce their minimum threshold to distension after inflammation, more of them are recruited above threshold and they generate larger responses. The levels of balloon distension at which they are activated correlate with stimuli that evoke pain in humans and animal models, placing them in a key role for clinically relevant visceral pain.28 29 Coupled with the fact that these afferents are particularly chemosensitive to inflammatory and enteroendocrine mediators, this places them in a central role for mediating symptoms in conditions such as PI-IBS.
It is important to note that in our original paper categorising mouse colonic afferents,11 we defined serosal afferents as those that had receptive fields located in the colonic wall that were reproducibly activated only by probing and did not respond to circular stretch (1–5 g) or fine mucosal stroking with a 10 mg von Frey hair. We did not investigate their responses to higher intensities of stretch, but speculated that they may be capable of responding to such stimuli in vivo. We later found that serosal afferents fitting these criteria responded also to noxious levels of distension of the intact colon by balloon, or circumferential stretch of the flat preparation in excess of 10 g.16 We confirmed and quantified these findings in the current study. Since our original definition of serosal afferents was based on their insensitivity to stretch, this suggested a location of their endings outside the muscular layers. In the light of our more recent data showing that these fibres do in fact respond to stretch at high intensity, it may be argued that their endings could penetrate more deeply within the gut wall, and that the term “serosal” may be inappropriate. Indeed, we have found endings on submucosal vessels in guinea-pig that respond to high intensity stretch.12 We would propose, therefore, that the current nomenclature of serosal afferents is retained until a detailed anatomical characterisation is made in mouse, because it is useful in discriminating these endings from bona fide muscular afferents and mesenteric afferents, whose anatomical location is better defined. From a functional point of view, however, it is clear we have described major post-inflammatory changes in a distinct population of afferents with high thresholds to distension.
The increase in pelvic mucosal afferent mechanosensitivity that we observed, mainly at the recovery stage, is of possible clinical significance. However, these changes were small in comparison with serosal and mesenteric afferents, and were only observed at high intensities of mucosal stroking stimuli, which these afferents are unlikely to encounter in situ. Their major role is considered to be in detection of stool consistency, by responding to shear forces during transit of solid stool or pellet.11
Although our study is concerned with changes at the level of the afferent ending, there are changes in excitability that are observable in their cell bodies that may be reflected in altered peripheral activation. These are reviewed by Beyak and Vanner,30 who describe in acute rat and mouse TNBS models decreased voltage-dependent K+ channel function and increased Na+ channel function, both of which lead to increased excitability. We did not observe appreciable levels of spontaneous firing in our study, nor was this revealed in the acute or post-inflammatory state. Therefore contributions of these channels toward baseline excitability would be masked. They may, however, be manifested in increased mechanosensory responses and reduced thresholds. There are numerous other candidates for mediating the altered mechanosensitivity and thresholds that we observed, including transient receptor potential (TRP) channels,16 25 31 32 and acid-sensing ion channels,22 33 but their precise roles will require study in the context of the specific subtypes we have identified here.
Little is currently known of the events leading to sensitisation of visceral afferents in post-inflammatory environments. However, even mild inflammatory events that do not cause acute sensitisation can induce chronic hypersensitivity.6 21 Our results demonstrate that mild inflammation results in an acute sensitisation of splanchnic but not pelvic afferents, adding to the growing literature describing differences between these pathways.11 13 14 16 Furthermore, we also demonstrate that, of all afferent classes, the most affected are those most distant from the inflammatory damage, such that serosal and mesenteric afferents are more affected than muscular or mucosal afferents. These afferents are implicated in detection of nociceptive events, and are in close contact with larger blood vessels, which may place them in a better position to detect circulating factors.16 23 34 Therefore, either blood-borne inflammatory mediators induce plasticity in extrinsic afferents, or factors released locally by the mucosa do so after diffusion through to the serosa and mesentery. Both are possible in the acute setting; however, in the post-inflammatory setting, the mucosa is healed and less likely to continue release of inflammatory mediators. The reason for hypersensitivity appearing only after considerable delay may reflect the recruitment of anti-nociceptive processes initially that wane over time. In this respect we have evidence for T-cell mediators that inhibit afferent activation, probably via an opioidergic mechanism,35 36 which serve to balance the influence of pro-algesic mediators that may be co-released.1 Whether such a mechanism is active within our preparation is the subject of continued detailed investigation. We have shown that mouse splanchnic afferents are more sensitive than pelvic afferents to mediators released in response to colonic inflammatory damage, including bradykinin and ATP,13 14 which may explain the earlier manifestation of hypersensitivity in the splanchnic pathway. It is also probable that the chronic hypersensitivity observed in the current study is due to circulating inflammatory mediators. Patients suffering post-infectious IBS have increased interleukin 1β and tumour necrosis factor α levels in peripheral blood mononuclear cells compared to healthy subjects, and immune cell supernatants from these patients sensitise both splanchnic and pelvic afferent mechanosensitivity in our mouse model.1 35 Little is otherwise known in animal models of post infectious IBS about changes in circulating cytokines such as interleukin 1β and tumour necrosis factor α or other mediators of inflammation.
In conclusion, we have described a model that shares many of the features of post-infectious IBS whereby mild inflammatory events sensitise colonic sensory pathways – initially only splanchnic afferents – but after healing, both splanchnic and pelvic afferents are hypersensitive. Furthermore, we have identified high threshold serosal and mesenteric afferent classes as the drivers of this mechanical hypersensitivity. This provides a complete picture of how the mechanosensory signal from the colon is amplified in conditions of visceral hypersensitivity. Our future studies will concentrate on mechanisms underlying this effect in order to produce targets for treatment of post-infectious IBS.
REFERENCES
Footnotes
Funding This work was supported by a National Health and Medical Research Council Senior Research Fellowship to LAB, a NHMRC Australian Biomedical Research Fellowship to SMB, and by NHMRC project grant #399324.
Competing interests None.
Provenance and Peer review Not commissioned; externally peer reviewed.
See Commentary, p 1317
Ethics approval Studies were performed with the approval of the Animal Ethics Committees of the Institute for Medical and Veterinary Science, and University of Adelaide.