Article Text

Abstract
Objective: In mice, a subpopulation of gut dendritic cells (DCs) expressing CD103 drives the development of regulatory T (Treg) cells. Further, it was recently described that the cross-talk between human intestinal epithelial cells (IECs) and DCs helps in maintaining gut immune homeostasis via the induction of non-inflammatory DCs. In this study, an analysis was carried out to determine whether IECs could promote the differentiation of CD103+ tolerogenic DCs, and the function of primary CD103+ DCs isolated from human mesenteric lymph nodes (MLNs) was evaluated.
Methods: Monocyte-derived DCs (MoDCs) and circulating CD1c+ DCs were conditioned or not with supernatants from Caco-2 cells or IECs isolated from healthy donors or donors with Crohn’s disease and analysed for their ability to induce Treg cell differentiation. In some cases, transforming growth factor β (TGFβ), retinoic acid (RA) or thymic stromal lymphopoietin (TSLP) were neutralised before conditioning. CD103+ and CD103− DCs were sorted by fluorescence-activated cell sorting (FACS) from MLNs and used in Treg cell differentiation experiments.
Results: It was found that human IECs promoted the differentiation of tolerogenic DCs able to drive the development of adaptive Foxp3+ Treg cells. This control was lost in patients with Crohn’s disease and paralleled a reduced expression of tolerogenic factors by primary IECs. MoDCs differentiated with RA or IEC supernatant upregulated the expression of CD103. Consistently, human primary CD103+ DCs isolated from MLNs were endowed with the ability to drive Treg cell differentiation. This subset of DCs expressed CCR7 and probably represents a lamina propria-derived migratory population.
Conclusions: A population of tolerogenic CD103+ DCs was identified in the human gut that probably differentiate in response to IEC-derived factors and drive Treg cell development.
Statistics from Altmetric.com
Recent evidence has suggested that tissue resident dendritic cells (DCs) are shaped by the local microenvironment to carry out specialised tissue-associated functions.1 A typical example is represented by DCs isolated from the gut.2 3 In mice, DCs from the lamina propria (LP), Peyer’s patches or mesenteric lymph nodes (MLNs) can confer gut tropism to T and B lymphocytes,4 5 6 7 8 can drive the differentiation of T regulatory (Treg) cells,9 10 and support the development of immunoglobulin A (IgA)-producing B cells.4 11 Furthermore, gut DCs have been shown to have reduced ability to release interleukin-12 (IL-12) and preferentially drive the T helper 2 (Th2)-type responses12 13 while a subset of LP-derived DCs can also drive the Th17 type of T cells.14 15 Little is known about DCs isolated from human tissues. We showed that, similarly to the mouse system, LP-derived DCs are unable to drive the Th1-type responses, even after a strong Th1 stimulus such as Salmonella.16 A subtype of MLN-derived DCs characterised by the expression of CD103 has been shown to confer gut tropism to CD8 T cells17 via a retinoic acid (RA)-dependent mechanism. It is possible that as in the mouse,8 17 these cells derive from the LP where they probably received tissue specialisations. It remains to be established which are the cells that might influence DC function and that release immunomodulatory mediators such as RA.
Epithelial cells (ECs) are ideal candidates to confer mucosal DC specialisations because of their proximity to DCs that occasionally extend protrusions for direct luminal sampling.18 19 20 We recently showed that intestinal ECs (IECs) release thymic stromal lymphopoietin (TSLP) that inhibits IL-12 production by DCs in response to bacteria and drives Th2-polarising cells, inhibiting the inflammatory potential of DCs.16 Notably, IECs isolated from 70% of patients with Crohn’s disease (CD) do not express TSLP and fail to control the DC proinflammatory response.16 In addition, TSLP promotes the release of BAFF and APRIL by EC-conditioned DCs and supports IgA class switching directly in the LP21 22 in humans. TSLP is also released by ECs in the Hassall’s corpuscles and drives Treg cell-inducing tolerogenic DCs23 in human thymuses, while TSLP-conditioned DCs drive Treg cells in the mouse.24
In this study we analysed whether IECs were involved in driving the development of tolerogenic DCs in the human system and whether deregulations of this function correlated with CD pathology. We additionally identified a population of CD103-expressing DCs in human MLNs that were potent inducers of Treg cell differentiation. This mechanism might play a crucial role in the maintenance of gut tolerance.
Materials and methods
DC cultures and peripheral blood DC purification
Human monocyte-derived DCs (MoDCs) were generated from CD14+ monocytes of healthy donors, as previously described.16 For the differentiation of CD103+ DCs from CD14+ monocytes, monocyte cultures were supplemented with supernatants derived from Caco-2 cells grown on Transwell filters as previously described,16 or with 1 μM RA (Sigma-Aldrich, Taufkirchen, Germany). Human peripheral blood CD1c+ DCs were purified from peripheral blood mononuclear cells (PBMCs) of buffy coats of volunteers by positive selection using the CD1c (BDCA-1) DC isolation kit (Miltenyi, Bergisch Gladbach, Germany).
MLN DC and IEC isolation
CD103+ and CD103− DCs were isolated from MLNs of patients undergoing surgery for colon cancer. MLNs were cut into pieces and incubated in digestion medium consisting of RPMI 1640, 5% fetal calf serum (FCS), 0.5 mg/ml collagenase type VIII (Sigma-Aldrich), 5 U/ml DNase (Roche Diagnostics, Nutley, New Jersey, USA), 100 IU/ml penicillin and 100 μg/ml streptomycin for 30 min at 37°C by gentle shaking. Cells were passed through a mesh, centrifuged and stained with lineage markers (anti-CD3, anti-CD14, anti-CD16, anti-CD19 and anti-CD56), anti-CD11c, anti-human leucocyte antigen (HLA)-DR, anti-CCR7 and anti-CD103 antibodies (BD Biosciences, Bedford, Massachusetts, USA). CD11c+HALDR+ DCs and, in some experiments, Lin−CD11c+HALDR+ DCs were sorted by fluorescence-activated cell sorting (FACS) (FACSAria; BD Biosciences) into CD103+ and CD103− DC populations. Human IECs were isolated from healthy tissue of patients with colon carcinoma (>7 cm from the neoplasm) or from patients with CD, according to a protocol established in our laboratory.16 All experimental procedures are in compliance with guidelines set by our review board after informed consent of the patients (European Institute of Oncology, Milan). Caco-2 and CL2 (a Caco-2 cell line treated with TSLP small interfering RNA (siRNA)16) were grown in Iscove’s modified Dulbecco’s medium (IMDM) supplemented with 10% FCS, 2 mmol/l glutamine, 100 IU/ml penicillin, 100 μg/ml streptomycin, 50 μmol/l 2-mercaptoethanol or in Advanced Dulbecco’s modified Eagle’s medium (DMEM; Gibco, Gaithersburg, Maryland, USA).
DC conditioning and T cell differentiation
MoDCs were incubated for 24 h with primary IECs, Caco-2 (neutralised or not for transforming growth factor β (TGFβ) or RA) or CL2 cell supernatants. Cells were extensively washed and then co-cultured in a mixed leukocyte reaction (MLR) for 5 days with naive CD4+CD45RA+CD25− T cells. Cells were analysed for Foxp3 expression by FACS. The suppressive capacity of newly developed Treg cells was examined in a suppression assay. The expression of different genes in DCs and IECs was analysed by quantitive PCR. A more detailed description of these methods is provided as supplementary information.
Statistical analysis
Student paired t test was used to determine the statistical significance of the data. Significance was defined at p<0.05 (two-tailed test and two-sample equal variance parameters). Statistic calculations were performed by JMP 5.1 software (SAS, Cary, North Carolina, USA).
Results
EC-conditioned DCs convert naïve T cells into Foxp3+ Treg cells
We previously showed that human gut resident DCs are non-inflammatory and that IECs with which they closely interact16 confer this property. Recent reports have demonstrated that DCs isolated from mouse gut can induce the development of Treg cell differentiation.9 10 25 Thus, we analysed whether MoDCs conditioned with IEC supernatants could also drive Treg cell differentiation. MoDCs were incubated with the supernatant of either Caco-2 or primary IECs isolated from the healthy tissue of patients with colon cancer and used for T cell activation assays in allogeneic MLRs. Notably, MoDCs derived from different donors when incubated with Caco-2 supernatants led to a greater conversion of naïve CD4+CD45RA+CD25−Foxp3− T cells into Foxp3+ Treg cells, compared with non-conditioned MoDCs, as analysed by flow cytometry (fig 1A) and by quantitative reverse transcription–PCR (RT–PCR) for FOXP3 gene expression (fig 1B). This was not a peculiarity of the Caco-2 cell line as a similar feature was observed when primary IECs were used to “educate” DCs (fig 1). Additionally, the increase in the percentage (fig 1C) and in the total number (fig 1D) of Foxp3-converted Treg cells was statistically significant when EC-conditioned DCs were used as antigen-presenting cells (APCs) compared with DCs. In order to exclude the expansion of a small population of already existing Treg cells, the experiments were conducted using highly purified CD4+CD45RA+CD25−Foxp3− T cells (Supplementary fig 1). Together, these results indicate that IECs can “educate” tolerogenic DCs to be able to induce the differentiation of Treg cells.

Epithelial cell (EC)-conditioned dendritic cells (DCs) promote T regulatory (Treg) cell differentiation in humans. Human monocyte-derived DCs (MoDCs) were incubated or not in the presence of Caco-2 supernatants or primary intestinal EC (IEC) supernatants for 24 h. DCs were co-cultured with naïve CD4+CD45RA+CD25−Foxp3− T cells in an allogenic mixed leukocyte reaction (MLR) for 5 days. (A) Caco-2 (middle, Caco2 sn) and primary IEC supernatants (right, EC sn) were equally potent in their ability to condition MoDCs to promote Treg cell development. Dot plots show CD4+Foxp3+ Treg cells. Numbers show the percentage of double-positive cells in the quadrant. Representative dot plots are shown. (B) Quantitative reverse transcription–PCR (RT–PCR) of FOXP3 expression in CD4+ T cells isolated from the MLR. FOXP3 was normalised to GAPDH (glyceraldehyde phosphate dehydrogenase) mRNA. (C) MoDCs derived from different donors show a similar response to IEC conditioning. The graph shows the percentage of Foxp3+ T cells among the CD4+ T cell population. Each symbol corresponds to an individual donor; lines represent the mean value. Two experiments were performed with MoDCs from five different donors. (D) Absolute numbers of CD4+Foxp3+ Treg cells in the cultures are shown. (B, D, error bars show the SD; B, C, D, *p<0.05; **p<0.01).
Peripheral blood DCs become tolerogenic after EC conditioning
To understand if other types of DCs could also be converted into being tolerogenic by IECs, CD1c+ cells were isolated from PBMCs and incubated with Caco-2 supernatants, as described above. Subsequently, naïve T cells were co-cultured with EC-primed DCs and after 5 days the number of newly developed Treg cells was evaluated. We found that similarly to MoDCs, peripheral blood CD1c+ DCs were able to induce CD4+CD25+Foxp3+ Treg cell differentiation (fig 2A,B). Together these data indicate that it is possible to drive the differentiation of tolerogenic DCs starting from different DC populations (MoDCs and CD1c+ blood DCs) by incubation in IEC supernatant.
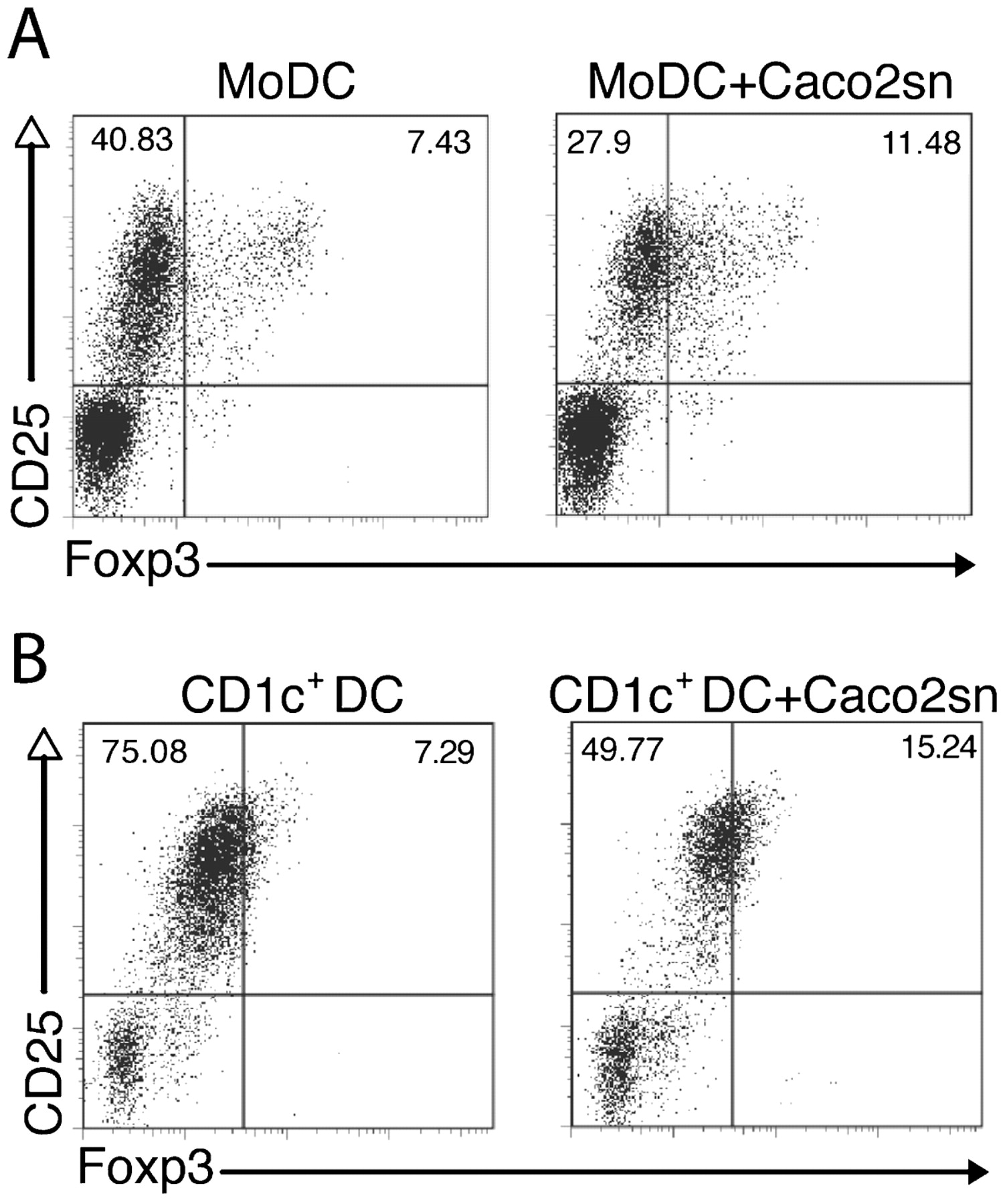
Peripheral blood dendritic cells (DCs) become tolerogenic after epithelial cell (EC) conditioning. Human monocyte-derived DCs (MoDCs) or CD1c+ cells purified from blood mononuclear cells were conditioned or not with Caco-2 supernatant (Caco2sn) and co-cultured with naïve T cells for 5 days. Dot plots show CD25+ and Foxp3+ T cells arising from 5 day cultures with conditioned or non-conditioned MoDCs (A) or CD1c+ cells (B). Numbers indicate the percentage of positive cells in the quadrants.
EC-conditioned DC-activated T cells are suppressive
We observed that in EC-conditioned DC–T cell co-cultures the amount of CD4+CD25+Foxp3− T cells was greatly reduced (fig 2A). As this population also includes activated T cells, we analysed whether EC conditioning reduced the ability of DCs to promote T cell proliferation. In the cultures where carboxyfluorescein succinimidyl ester (CFSE)-labelled T cells were incubated with EC-conditioned DCs, there was a correlation between Treg cell development and decreased T cell proliferation (fig 3A, lower panels). Confirming previous data,9 Foxp3+ cells were proliferating cells as they diluted the CFSE staining (fig 3A, upper panels).

Epithelial cell (EC)-conditioned dendritic cells (DCs) induce less T cell proliferation and drive the development of suppressor CD25+Foxp3+ T cells. EC-conditioned DCs promote reduced proliferation of naïve T cells. (A) CD4+CD45RA+CD25− naïve T cells were stained with carboxyfluorescein succinimidyl ester (CFSE) and incubated with monocyte-derived DCs (MoDCs) or EC-conditioned MoDCs for 5 days. Dot plots illustrate CFSE+ and Foxp3+ T cells (upper panels); histograms show CFSE staining for the total CD4+ cells (lower panels). Numbers indicate the percentage of positive cells in the gates. (B) EC-conditioned DCs promote the development of highly suppressive T regulatory (Treg) cells. CD4+CD25+ T cells were purified from the primary mixed lymphocyte reactions (MLRs) where MoDCs were incubated or not with Caco-2 supernatant (Caco2sn) and were added at different ratios (1:5, 1:2.5 and 1:1) to naive T cells in a second MLR using lipopolysaccharide (LPS)-activated DCs as APCs. CD4+CD25+ T cells isolated from blood were used as positive controls of suppression. T cell effectors (CD4+CD25−) incubated with LPS-activated DCs without CD25+ cells are also shown to assess maximal proliferation. Incorporation of [3H]thymidine during the last 16 h is shown. Error bars show the SD. Data are representative of three independent experiments. PBMC, peripheral blood mononuclear cell.
In order to confirm that EC-conditioned DC-converted cells were functionally active Treg cells, we performed suppressor assays in which CD4+CD25+ T cells isolated from the primary MLRs were added to a second proliferating DC–T cell co-culture. CD4+CD25+ and CD4+CD25− T cells isolated from PBMCs were used as controls. As shown in fig 3B, CD4+CD25+ Treg cells generated by EC-conditioned DCs were as potent as CD4+CD25+ T cells isolated from PBMCs in suppressing T cell proliferation in the second co-culture. The effect was already detectable at a low Treg:Teff cell ratio (1:5), confirming strong suppressor activity. Together, these results indicate that EC-conditioned DCs induce the development of highly suppressive Treg cells.
IECs from patients with CD show impaired ability to induce tolerogenic DCs
Defects in IEC function have been proposed to contribute to the pathology of CD.26 Further, reduced numbers of Treg cells have been described in patients with inflammatory bowel disease.27 To understand whether the erroneous behaviour of IECs is directly responsible for a decrease in Treg cell conversion in patients with CD, we compared the ability of healthy primary IECs versus IECs isolated from the non-inflamed region of resected colons of patients with CD to promote the development of tolerogenic DCs. Interestingly, IECs isolated from patients with CD displayed highly reduced ability to drive Treg cell-promoting DCs (fig 4A,B). This correlated with significantly reduced expression of TGFB1, TSLP and ALDH1A1 (aldehyde dehydrogenase 1A1) genes in IECs isolated from colons of patients with CD compared with healthy controls (fig 4C), suggesting a possible involvement of their products in DC conditioning.
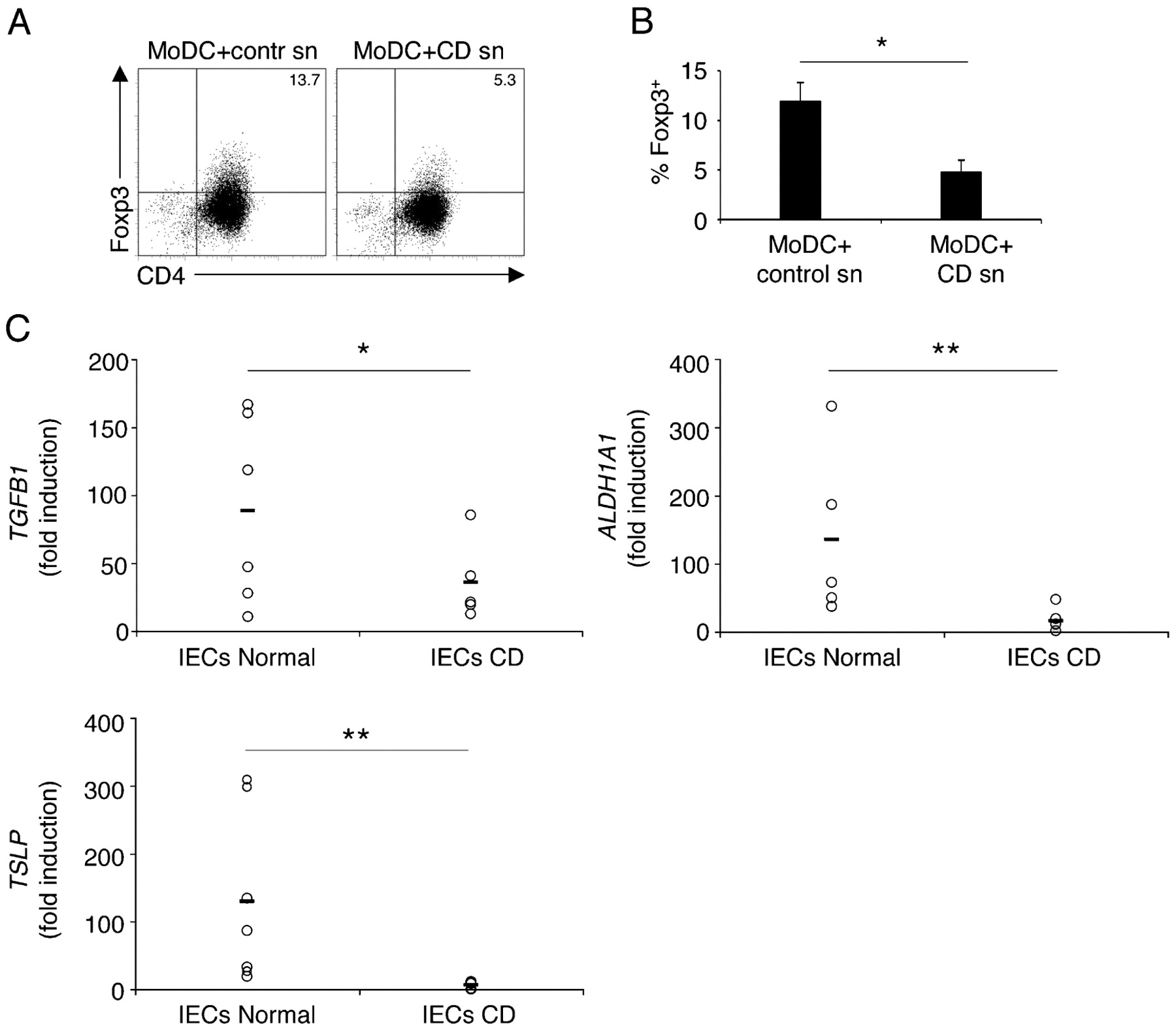
Intestinal epithelial cells (IECs) from patients with Crohn’s disease (CD) show impaired ability to drive tolerogenic dendritic cells (DCs). (A) Human monocyte-derived dendritic cells (MoDCs) were incubated with supernatants (sn) of primary IECs isolated from healthy tissue of patients with colon cancer or from non-inflamed regions of resected colons of patients with CD. Conditioned DCs were co-cultured with naïve T cells, as described in fig 1. Dot plots represent CD4+Foxp3+ T regulatory (Treg) cells and numbers show the percentage of double-positive cells in the quadrant. Two experiments were performed with IECs from five different patients from each group. Representative dot plots are shown. (B) The graph represents pooled data from the experiments in A. (C) Quantitative reverse transcription–PCR (RT–PCR) of expression of the genes encoding transformig growth factor β1 (TGFB1), aldehyde dehydrogenase 1A1 (ALDH1A1) and thymic stromal lymphopoietin (TSLP) in primary IECs isolated from healthy tissue of patients with colon cancer (IECs Normal) or from resected colons of patients with CD (IECs CD). Each symbol corresponds to an individual patient; lines represent the mean value (B. error bars show the SD; B, C, *p<0.05; **p<0.01).
EC-derived TGFβ drives DC-dependent Treg cell conversion
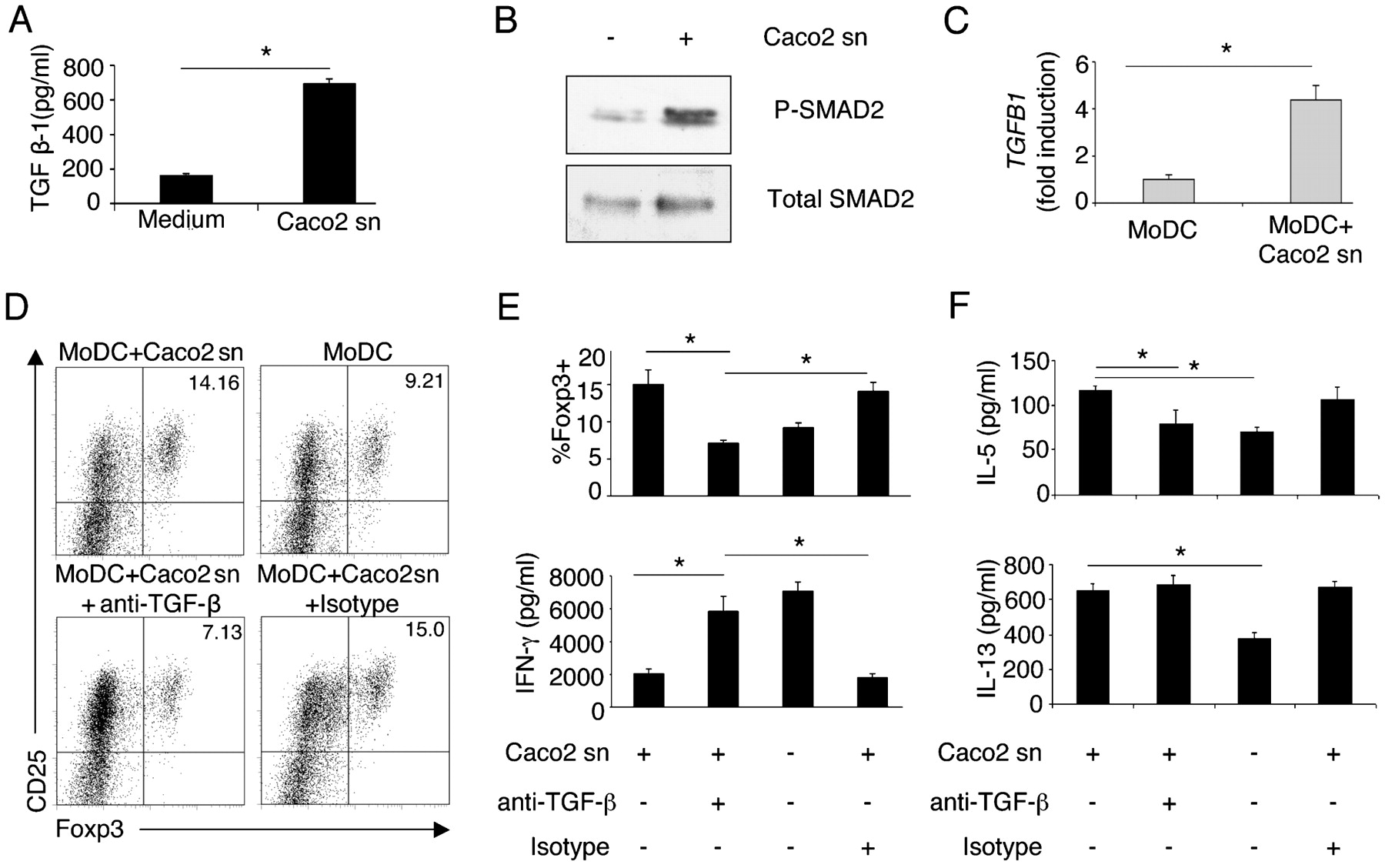
TGFβ is a regulatory cytokine that has a direct effect on gut APCs, including DCs,28 29 and is important for the generation and maintenance of Treg cells.9 10 25 30 31 We found decreased TGFB1 expression in IECs isolated from the colons of patients with CD (fig 4C), hence we evaluated whether EC-derived TGFβ might be acting on the DCs, which upon stimulation would induce Treg cell differentiation. To test this hypothesis we examined TGFβ production by IECs. We observed that Caco-2 cells secreted TGFβ (fig 5A) which directly acted on DCs by switching on SMAD signalling and inducing SMAD2 phosphorylation (fig 5B). Notably, DCs acquired TGFβ-producing ability after incubation in the IEC supernatants as assessed by TGFB1 expression (fig 5C). DCs conditioned with IEC supernatants neutralised with anti-TGFβ antibodies displayed reduced ability to induce Treg cell differentiation (fig 5D,E; Supplementary fig 2A). This correlated with enhanced interferon γ (IFNγ) and decreased IL-5 production in the MLRs (fig 5E, F). To distinguish between the need for serum-derived or cell-derived TGFβ, we used supernatants of Caco-2 cells or primary IECs grown in TGFβ-free medium. We observed a trend of tolerogenic DC induction similar to that of cells grown in medium supplemented with serum, suggesting that cell-derived TGFβ is sufficient for DC conditioning (Supplementary fig 2A). To discriminate between the requirement for TGFβ produced by IECs and that produced by DCs in the development of Treg cells, we depleted the IEC supernatant of TGFβ using a microbead-based immunoprecipitation technique (Supplementary fig 2B). This procedure allows depletion of TGFβ but also allows unbound antibody that could bind to TGFβ produced by DCs to be disposed of. TGFβ-depleted supernatants (Supplementary fig 2B) were then used for conditioning of DCs. As shown in Supplementary fig 2C, TGFβ released by IECs is the one required to condition tolerogenic DCs properly.

Epithelial cell (EC)-derived transforming growth factor β (TGFβ) is required to educate dendritic cells (DCs) to promote regulatory T (Treg) cell differentiation. (A) Caco-2 cells were grown in advanced Dulbecco’s modified Eagle’s medium (DMEM) and the amount of released TGFβ1 was measured by ELISA. (B) SMAD2 phosphorylation in monocyte-derived DCs (MoDCs) without (–) or with (+) Caco-2 supernatant (Caco2 sn) conditioning was detected by western blotting. Total SMAD2 is also shown. (C) Quantitative reverse transcription–PCR (RT–PCR) of TGFB1 expression in MoDCs incubated or not with Caco-2 supernatants (sn). TGFB1 was normalised to glyceraldehyde phosphate dehydrogenase (GAPDH) mRNA. (D–F) Neutralisation of TGFβ inhibits Treg cell differentiation, restores interferon γ (IFNγ) production and reduces interleukin-5 (IL-5) production. (D) Dot plots show CD25+Foxp3+ Treg cells in cultures of MoDCs incubated (MoDC+Caco2 sn) or not (MoDC) with Caco-2 supernatant neutralised with anti-TGFβ antibody (MoDC+Caco2 sn+anti-TGF-β) or treated with isotype control (MoDC+Caco2 sn+isotype). Numbers show the percentage of positive cells in the quadrant. One of two representative experiments is shown. (E) Pooled data representing the percentage of Foxp3+ cells (upper graph) and IFNγ production in mixed leucoyte reactions (MLRs) (lower graph) from two experiments. (F) IL-5 and IL-13 production in culture supernatants from the experiments in D–E. A, C, E, F; error bars show the SD. *p<0.05.
IEC-derived RA and TSLP are also involved in the induction of tolerogenic DCs
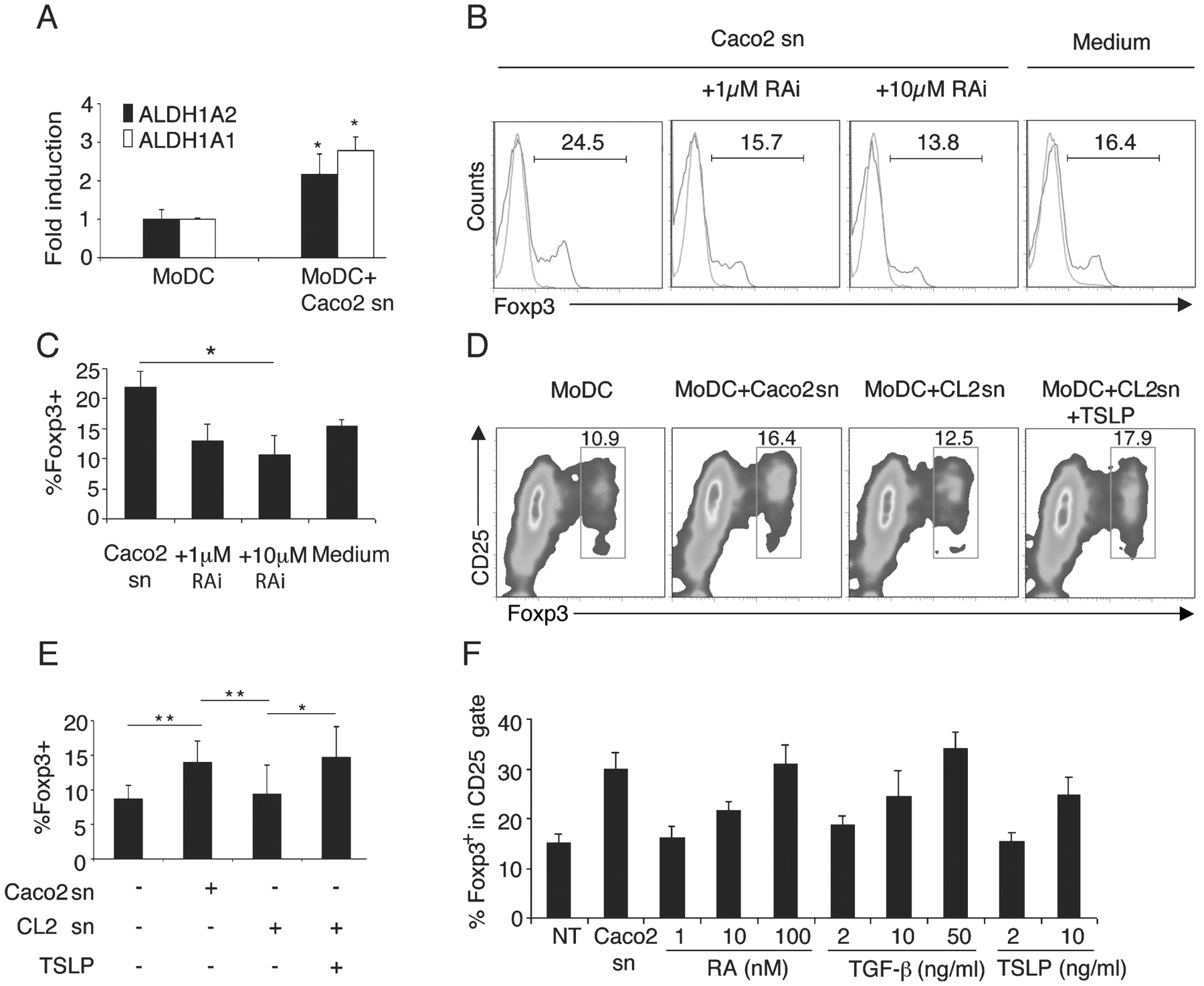
All-trans RA has been shown to be a cofactor of TGFβ in the induction of Treg cells.9 10 We tested whether RA was also required for the observed differentiation of tolerogenic DCs because we found that EC-conditioned DCs upregulated the expression of ALDH1A2 and ALDH1A1 after conditioning (fig 6A). These retinal hydrogenases are involved in the conversion of retinal to RA. IEC supernatants were incubated with an RA receptor inhibitor (LE540) for 1 h before conditioning. Twenty-four hours later DCs were washed and incubated with allogeneic naïve T cells, as above. We found that inhibition of RA signalling on DCs significantly affected the induction of Treg cells (fig 6B,C). We could not detect any synergistic effect in Treg cell differentiation by inhibiting both TGFβ and RA (data not shown).

Retinoic acid (RA) and thymic stromal lymphopoietin (TSLP) are also necessary to drive tolerogenic dendritic cells (DCs). (A) Quantitative reverse transcription–PCR (RT–PCR) of the expression of the genes encoding aldehyde dehydrogenase 1A1 and 1A2 (ALDH1A1 and ALDH1A2) in monocyte-derived DCs (MoDCs) incubated or not in Caco-2 supernatant (sn) and normalised to glyceraldehyde phosphate dehydrogenase (GAPDH) mRNA is shown. (B) Inhibition of RA signalling blocks regulatory T (Treg) cell differentiation. MoDCs were conditioned with Caco-2 supernatants preincubated with the RA receptor inhibitor LE540 (RAi), and treated with lipopolysaccharide (LPS). After extensive washing, DCs were co-cultured with naïve T cells for 5 days. Histograms show Foxp3+ T cells in the CD4+ gate (red histograms) in the co-cultures as analysed by flow cytometry overlaid on the isotype control (green histograms). Numbers indicate the percentage of Foxp3+ T cells in the marker. (C) Statistics of the percentage of Foxp3+ cells after RA receptor inhibition from pooled data of two experiments as in B. (D) TSLP is required for the induction of tolerogenic DCs. MoDCs were incubated with Caco-2 supernatants (MoDC+Caco2 sn), CL2 supernatants (a Caco-2 cell line treated with TSLP small interfering RNA (siRNA); MoDC+CL2 sn) or CL2 supernatants supplemented with recombinant TSLP (MoDC+CL2 sn+TSLP) for 24 h, treated with LPS and co-cultured with naïve T cells in a mixed leucocyte reaction (MLR) for 5 days. Dot plots of CD25+Foxp3+ cells are shown. Numbers represent the percentage of cells in the red gate. Pooled data from two independent experiments are shown in E. (F) DCs were incubated (Caco2 sn) or not (NT) for 24 h with Caco-2 supernatant or with the stated concentrations of either RA, transforming growth factor β (TGFβ) or TSLP and co-cultured with naïve T cells as described in fig 1A. The graph depicts the percentage of Foxp3+ T cells in the CD25+ gate. Data are representative of two independent experiments. A, C, E, F, error bars show the SD; *p<0.05; **p<0.01.
TSLP, which is released by the Hassal’s corpuscles in the thymus, activates thymic DCs to induce the development of Treg cells.23 We recently showed that human IECs constitutively express TSLP16 and we found decreased TSLP expression in IECs isolated from the colons of patients with CD (fig 4C). Thus we tested whether EC-derived TSLP was also involved in the induction of tolerogenic DCs. To address this issue we used a clone of Caco-2 cells treated with TSLP siRNA (CL2), as already described.16 In the absence of TSLP, the ability of DCs to drive Treg cell differentiation was also impaired (it dropped from 16.4% to 12.5%) and could be restored by adding exogenous TSLP (fig 6D,E).
Next, we evaluated whether each factor could individually induce the tolerogenic phenotype of DCs. We thus incubated DCs with different concentrations of either RA (1–100 nM), TGFβ (2–50 ng/ml) or TSLP (2–10 ng/ml). We found that only at the highest concentration was each factor able to drive the development of Foxp3+ T cell-inducing DCs (fig 6F). At low concentrations, resembling those present in the IEC supernatants, the factors were unable to drive tolerogenic DCs. It is likely that these factors act in a concerted fashion and this would explain why inhibition of any one of them in the IEC supernatant abolished the development of Treg cells. Thus, EC-derived factors, including TGFβ, RA and TSLP, are required for the generation of tolerogenic DCs.
CD103+ DCs isolated from human MLNs induce Treg cell differentiation
It has been recently shown that murine LP and MLN CD103+ DCs can drive the differentiation of Treg cells.9 10 This population represents about 40% of total MLN DCs and has been shown to imprint gut homing properties to T cells via the upregulation of CCR9 and α4β7 expression on responding T cells.8 17 However, the tolerogenic properties of human CD103+ DCs have not been analysed. Thus, we stained for Lin− MLN cells and sorted the CD11c+HLADR+ population of MLN DCs into CD103+ and CD103− fractions (fig 7A). These cells were also distinguishable in the expression of CCR7, a chemokine receptor involved in DC migration to draining LNs. CD103+ cells expressed CCR7 (fig 7B), suggesting that they may migrate from the LP. We then analysed the ability of these two subpopulations to convert naïve CD4+CD45RA+CD25−Foxp3− T cells into Foxp3+ Treg cells. As shown in fig 7C and D, FACS-sorted CD103+ cells displayed a >4 four times higher ability to drive Treg cell development than CD103− DCs, confirming that CD103 is also a marker of tolerogenic DCs in humans. In addition, we found that CD103+ DCs suppressed the induction of the Th1 responses, as measured by IFNγ production in the MLRs (fig 7E). Thus our results suggest that also in humans CD103+ DCs might represent tolerogenic LP-derived migratory cells.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Human mesenteric lymph node (MLN) CD103+ dendritic cells (DCs) drive regulatory T (Treg) cell development and intestinal epithelial cells (IECs) can induce CD103 expression on differentiating monocyte-derived DCs (MoDCs). (A) Human MLN CD11c+HLADR+ DCs were sorted by flow cytometry into CD103+ or CD103− fractions (representative plots are shown). (B) CCR7 expression on CD103+ (upper histogram) or CD103− MLN DCs (lower histogram) is shown. Grey histograms represents isotype control; black histograms show the expression of CCR7. (C) CD103+ or CD103− DCs were co-cultured with naïve CD4+CD45RA+CD25−Foxp3− T cells in an allogeneic mixed leucocyte reaction (MLR) for 6 days. Dot plots show CD4+Foxp3+ Treg cells. Numbers show the percentage of double-positive cells in the quadrant. Representative dot plots are shown. (D) The graph shows the percentage of Foxp3+ T cells in the CD4+ T cell population. Each symbol corresponds to an individual experiment using CD103+ and CD103− DCs isolated from MLNs of individual donors; lines represent the mean value. (E) Interferon γ (IFNγ) production in culture supernatants from the experiments in (D) was measured by ELISA. Results show the mean of five independent experiments. Error bars show the SD; *p<0.05. (F) CD14+ monocytes were cultured in differentiation medium containing granulocyte–macrophage colony-stimulating factor (GM-CSF) and interleukin-4 (IL-4), in the presence or absence of Caco-2 supernatants (sn), retinoic acid (RA; 1 μM) or recombinant transforming growth factor β (TGFβ; 2 ng/ml). After 6 days of culture, cells were collected and stained for CD11c, human leucocyte antigen (HLA)-DR and CD103. Histograms represent CD103+ cells within the CD11c-positive gate. Grey histograms represent the basal level of CD103 expression in conventional MoDCs; dark histograms show CD103 expression by MoDCs differentiated as described (Caco2 sn, RA or TGFβ). Data are representative of two independent experiments.
IECs upregulate CD103 expression on DCs differentiating from monocytes
A recent report has suggested that CD103+ DCs in mouse LP differentiate from blood-derived DC precursors.8 17 Due to the close proximity between IECs and DCs in the LP,18 19 20 we hypothesised that EC–DC interactions might be involved in inducing CD103 expression during their differentiation from blood precursors. Thus, we exposed circulating CD14+ monocytes to supernatants derived from the Caco-2 cell line in the presence of granulocyte–macrophage colony-stimulating factor (GM–CSF) and IL-4. After 6 days of differentiation, cells were collected and stained for CD11c, HLA-DR and CD103. Exogenous RA and TGFβ were used in independent experiments. Interestingly, a high percentage of MoDCs upregulated CD103 when differentiated in the presence of Caco-2 supernatants (fig 7F). In contrast, MoDCs that were differentiated in standard conditions (with GM-CSF and IL-4 but without Caco-2 supernatants) did not upregulate CD103 (fig 7F). Interestingly, incubation with Caco-2 supernatants could not induce CD103 upregulation on already differentiated MoDCs (data not shown). CD103 expression on MoDCs was strongly promoted by exogenous RA, but not by TGFβ (fig 7F), suggesting that RA may be involved in CD103 upregulation in the gut, where it is abundantly expressed and accumulated in different cell types, including IECs. These results suggest that ECs in the intestine might be involved in the induction of DCs with a full tolerogenic phenotype.
Discussion
Gut DCs confer “mucosal” properties and tropism on effector immune cells, but still very little is known about what drives the “mucosal” phenotype of the DCs. Our results indicate that IECs participate in the development of “tolerogenic mucosal” DCs. EC-conditioned DCs drive the de novo generation of CD4+CD25+Foxp3+ T cells that are able to inhibit T cell proliferation. Soluble factors constitutively released by IECs are responsible for the observed effect. We found that EC-derived TGFβ and RA are necessary to drive the tolerogenic phenotype of DCs, because neutralisation of either one was sufficient to inhibit Treg cell differentiation. This is not surprising as RA signalling synergises with TGFβ to favour Treg cell conversion.9 10 We found that similarly to the thymus,23 TSLP was also involved in driving tolerogenic DCs. This suggests that in humans, central and peripheral naturally occurring Treg cells are generated via similar mechanisms. Notably, the addition of each factor separately (TGFβ, RA or TSLP) to human MoDCs led to tolerogenic DCs only at a very high concentration. At concentrations resembling those present in the supernatant, the factors were unable to induce tolerogenic DCs, suggesting that a concerted action is needed.
LP and MLN CD103+ DCs from both humans and mice have been shown to induce CCR9 and α4β7 expression on responding T cells, and thus to be involved in imprinting T cells with gut homing properties.8 17 Our results extend these findings and show that human CD103+ DCs are also involved in the induction of Treg cell differentiation. Similarly to murine CD103+ DCs,8 17 25 human MLN-derived CD103+ DCs express CCR7 and might represent a migratory population from the LP. A recent report suggests that in the mouse CD103+ DCs develop from circulating precursors,8 17 and not by already differentiated CD103− DCs. It can be hypothesised that monocytes arriving in the intestine from the bloodstream can develop into tolerogenic DCs after their exposure to the specific gut microenvironment. Because of the close proximity of IECs and DCs in the LP,18 19 20 an EC–DC interaction might contribute to confer a full tolerogenic phenotype to DCs. Indeed our results suggest that this might be the case and here we showed that MoDCs differentiated in the presence of IEC-derived factors became tolerogenic and upregulated CD103. This phenomenon took place only during the differentiation process from monocytes, since MoDCs that were already differentiated did not upregulate CD103 after additional incubation with Caco-2 supernatants. We additionally showed that the vitamin A metabolite RA, which is abundantly present in the gut and accumulates in different cellular types, including IECs, seem to be critically involved in this process as it strongly induced CD103 expression on differentiating MoDCs.
Thus, we describe that IECs are conductors of DC function, and disrupting this balance could result in inflammatory disorders. In agreement with this, IECs isolated from patients with CD showed decreased expression of genes encoding TGFβ, ALDH1A1 and TSLP, and were unable to imprint DCs with the ability to induce Treg cells. It remains to be elucidated whether this could contribute to the onset of the disease or could simply reflect a secondary effect of the ongoing inflammation.
We describe that IECs participate in controlling DC function by conferring on them both a non-inflammatory and a tolerogenic “mucosal” phenotype. This allows the DCs to avoid over-reacting to intestinal bacteria, but presumably also to initiate tolerance towards luminal antigens. We have preliminary evidence indicating that “educated” DCs are irreversibly blocked in their tolerogenic phenotype, suggesting that if an immunogenic response needs to be initiated new non-conditioned cells should be recruited from nearby areas or from blood. Given the difficulty in isolating DCs from the gut, one could consider using these irreversibly “tolerogenic” DCs in the treatment of inflammatory intestinal disorders.
Another important therapeutic implication of our study is that the local microenvironment could be a target of immunomodulatory drugs aimed at restoring the potential to educate resident DCs. In this regard, recent evidence has shown that incubation of IECs with bacteria resulted in different levels of TSLP and TGFβ1 upregulation depending on the nature of the bacteria encountered in vitro, with Gram-negative species being the most effective.32 Probiotics, described as bacteria able to confer a benefit to human health, have been shown to protect from experimental colitis by increasing the number of Treg cells.33 We identified some strains of probiotics able to increase the levels of TGFβ and TSLP in IECs and to favour the development of non-inflammatory DCs.34 Together, this evidence could provide a rationale for a probiotic-based dietary regimen to help in restoring immune homeostasis and presumably to retard recurrences in patients with inflammatory bowel disease.
In conclusion, our results not only shed light on the requirements to maintain a tolerogenic gut microenvironment, but set the basis for future development of cell-based therapies for the treatment of intestinal inflammatory disorders.
Acknowledgments
We would like to thank Simona Ronzoni for the valuable technical assistance with FACS.
REFERENCES
Supplementary materials
Web only appendices 58;11:1481
Files in this Data Supplement:
Footnotes
Funding This study was supported by the Crohn’s and Colitis Foundation of America (CCFA to MR); Italian Association for Cancer Research (AIRC to MR); European Research Council (ERC to MR); and the EC 7th Framework Program (IBDase, METAhit to MR).
Competing interests None.
Provenance and Peer review Not commissioned; externally peer reviewed.
Ethics approval The local ethics committee approved the informed consent specifying that material not needed for diagnostic purposes could be used for research purposes.
▸ A more detailed description of the methods and additional figures are published online only at http://gut.bmj.com/content/vol58/issue11
Linked Articles
- Digest