Article Text

Abstract
Background Hepatocellular carcinoma (HCC) is a typical inflammation-associated cancer, but may also provoke antitumour immune responses whose significance and underlying mechanisms are incompletely understood.
Objective To characterise immune responses in the diethylnitrosamine (DEN)-liver cancer mouse model.
Design Tumour development and immune cell functions upon DEN treatment were compared between C57BL/6 wild-type (WT), chemokine scavenging receptor D6-deficient, B cell- (Igh6), CD4 T cell- (MHC-II) and T-/B cell-deficient (Rag1) mice. Relevance for human HCC was tested by comparing gene array results from 139 HCC tissues.
Results The induction of premalignant lesions after 24 weeks and of HCC-like tumours after 42 weeks by DEN in mice was accompanied by significant leucocyte infiltration in the liver and upregulation of distinct intrahepatic chemokines (CCL2, CCL5, CXCL9). Macrophages and CD8 (cytotoxic) T cells were most prominently enriched in tumour-bearing livers, similar to samples from human HCC. Myeloid-derived suppressor cells (MDSC) increased in extrahepatic compartments of DEN-treated mice (bone marrow, spleen). The contribution of immune cell subsets for DEN-induced hepatocarcinogenesis was functionally dissected. In D6−/− mice, which lack the chemokine scavenging receptor D6, hepatic macrophage infiltration was significantly increased, but tumour formation and progression did not differ from that of WT mice. In contrast, progression of hepatic tumours (numbers, diameters, tumour load) was strikingly enhanced in T-/B cell-deficient Rag1−/− mice upon DEN treatment. When mice deficient for B cells (Igh6−/−, μMT) or major histocompatibility complex II were used, the data indicated that T cells prevent initial tumour formation, while B cells critically limit growth of established tumours. Accordingly, in tumour-bearing mice antibody production against liver-related model antigen was enhanced, indicating tumour-associated B cell activation. In agreement, T and B cell pathways were differentially regulated in gene array analyses from 139 human HCC tissues and significantly associated with patients' survival.
Conclusions Distinct axes of the adaptive immune system, which are also prognostic in human HCC, actively suppress DEN-induced hepatocarcinogenesis by controlling tumour formation and progression.
- HCC
- macrophages
- adaptive immunity
- chemokine
- den
- hepatitis
- acute hepatitis
- antibody targeted therapy
- autoimmune liver disease
- gastric cancer
- acute liver failure
- cancer genetics
- carcinogenesis
- apoptosis
- hepatocellular carcinoma
- immune-mediated liver damage
- immunology in hepatology
- immunotherapy
- liver immunology
- liver
- molecular biology
- molecular carcinogenesis
- cell death
- chronic liver disease
- liver cirrhosis
- molecular genetics
- fibrosis
- linkage analysis
- macrophages
- monocytes
- IL-6
- hepatitis C
Statistics from Altmetric.com
- HCC
- macrophages
- adaptive immunity
- chemokine
- den
- hepatitis
- acute hepatitis
- antibody targeted therapy
- autoimmune liver disease
- gastric cancer
- acute liver failure
- cancer genetics
- carcinogenesis
- apoptosis
- hepatocellular carcinoma
- immune-mediated liver damage
- immunology in hepatology
- immunotherapy
- liver immunology
- liver
- molecular biology
- molecular carcinogenesis
- cell death
- chronic liver disease
- liver cirrhosis
- molecular genetics
- fibrosis
- linkage analysis
- macrophages
- monocytes
- IL-6
- hepatitis C
Significance of this study
What is already known about this subject?
-
Hepatocellular carcinoma (HCC) is an inflammation-associated cancer, as it typically arises in inflamed liver owing to hepatitis and fibrosis.
-
Antitumoural immune responses have been identified in many cancers, but their role in restricting hepatocarcinogenesis is not fully understood.
What are the new findings?
-
Using the well-established model of diethylnitrosamine (DEN)-induced hepatocarcinogenesis in mice, we show that liver tumour development provokes local and systemic inflammatory responses, characterised by intrahepatic induction of distinct proinflammatory (macrophage- and T cell-attracting) chemokines, intrahepatic accumulation of macrophages and CD8 cytotoxic T cells and extrahepatic expansion of myeloid-derived suppressor cells.
-
By employing several knockout mouse lines, including chemokine-scavenging-deficient D6−/− mice, T-/B cell-deficient Rag1−/− mice, B cell-deficient Igh6−/− and CD4 T-helper cell-deficient major histocompatibility complex II−/− mice, we further functionally dissected these tumour-associated immune responses and showed that adaptive immune cells critically control hepatocarcinogenesis, with B cells appearing essentially involved in limiting tumour growth and T cells controlling initial tumour formation.
-
Gene array based analyses from human HCC samples (n=139) support a crucial role of adaptive immunity for the progression of HCC and the survival of patients.
Significance of this study
How might it impact on clinical practice in the foreseeable future?
-
Because distinct axes of the adaptive immune system, which are also prognostic in human HCC, actively suppress DEN-induced hepatocarcinogenesis, future therapeutic approaches for HCC may be directed against tumour-promoting inflammatory reactions, and may also augment HCC-induced antitumour adaptive immune cell functions.
Introduction
Hepatocellular carcinoma (HCC) is among the most common malignancies, representing the fourth most common cause of cancer mortality worldwide. Typically, HCC occurs in patients with chronic inflammatory liver diseases, such as hepatitis B or C virus infections, autoimmune disorders and other conditions causing cirrhosis—for example, alcoholism.1 The inflammation leading to fibrosis and cirrhosis is regarded as a crucial factor in the carcinogenesis of liver tissue.1 ,2 However, the mechanisms by which chronic inflammation induces HCC are not well understood. High cell turn over with accumulation of mutations, reactive oxygen species and proliferation signals provided by immune cells seem to facilitate malignant transformation.3 Tumour necrosis factor production and NF-κB activation in hepatocytes are also importantly involved in hepatocarcinogenesis.2 ,4
The interactions between cancer and immune cells have been studied in various human tumour entities and animal models, providing insights into the crosstalk between malignant cells, peritumoural stroma and cells of innate and adaptive immunity.5 Tumours can be induced by malignant transformation through sustained inflammation, and once established, they use the immune system to their advantage by forcing immune cells to provide survival signals. Furthermore, they develop elaborate strategies to evade detection and destruction by immune cells—for instance, by favouring the generation of immunosuppressive cells such as regulatory T cells or myeloid-derived suppressor cells.6 Several studies have indicated that antitumoural immune reactions, but also tumour-triggered, immune-evasion mechanisms are present in HCC.7 While it has long been known that inflammation triggers HCC, the extent to which the tumour cells trigger antitumour immune responses and whether and how these in turn impact tumourigenesis is still debated.
We chose to examine this question by inducing liver tumours in mice through administration of the chemical carcinogen diethylnitrosamine (DEN) to 14-day-old mice. This leads to DNA alterations by ethylation and consequently to neoplastic foci.8 Eventually, male mice will develop HCCs within 40 weeks without sustained alterations of the remaining liver tissue.9 Although DEN administration is a well-established and the most widely used mouse model for hepatocarcinogenesis, it has distinct features compared with other primary liver tumour models, because tumours arise from chemically induced mutations without initial underlying hepatitis.8 ,10 We herein demonstrate that DEN-induced hepatocarcinogenesis triggered local and systemic inflammatory reactions. Moreover, using genetically modified mouse models and human HCC gene array datasets, we demonstrate previously unrecognised and distinct functions of adaptive immune cells (cytotoxic T cells and B cells) in suppressing HCC initiation and progression, respectively.
Materials and methods
Mice
C57BL/6 wild-type (WT), D6−/−,11 Rag1−/−, Igh6−/− and major histocompatibility complex (MHC)II−/− 12 mice (backcrossed to C57BL/6 background for more than eight generations) were maintained in our colony in a specific pathogen-free environment. All experiments were performed under ethical conditions approved by appropriate authorities according to German legal requirements. Male mice were injected intraperitoneally with 25 mg/kg body weight of DEN (Sigma-Aldrich, Munich, Germany) at 14 days of age.3 Mice were killed at 24 or 42 weeks after injection. Vehicle-injected (saline) 44-week old male mice served as controls. All animals were injected within a time period of 3 months with the same DEN lot, ensuring best comparability between treated groups.
Analysis of liver tumours
Each liver lobe was separately photographed with the Leica Z16 APO macroscope from every side. Tumour formations >1 mm in diameter were counted and measured using the calibrated Diskus software (Hilgers, Königswinter, Germany).13 Conventional haematoxylin–eosin stainings were evaluated for signs of malignancies by an experienced and blinded pathologist, using a modified score based on UICC grading for HCC (supplementary table 1).
FACS analysis of intrahepatic leucocytes, blood, spleen and bone marrow
Intrahepatic leukocytes were isolated and subjected to FACS analysis as described previously.14 ,15 Blood, bone marrow and spleen were also analysed by FACS as previously described.12 Six-colour staining was conducted using combinations of the following monoclonal antibodies: F4/80 (Serotec, Oxford, UK), CD115, CD4, CD11c, CD11b (all eBioscience, San Diego, CA, USA), CD45, Gr-1/Ly6C, Ly6G, CD19, NK1.1, CD8, CD3, or IgG isotype controls (all BD, Franklin Lakes, NJ, USA). Flow cytometric analysis was performed on a FACS-Canto II (BD) and analysed with FlowJo (Tree Star, Ashland, OR, USA). An example of a representative gating strategy is provided in supplementary figure 1. For comparability between livers and cell subsets, the total number of each population was normalised to the amount of total live liver cells, leucocytes or lymphocytes, as indicated.16
Immunohistochemistry
Immunohistological staining was performed on paraffin-embedded sections using standard protocols.17 Antibodies used were F4/80 (Abcam, Cambridge, UK) and Ki-67 (Dako, Glostrup, Denmark) for mice, and CD68, CD20, CD3, CD8 and MPO (Dako) for human tissue. Detection was performed using Anti-Ig HRP Detection Kit (BD).
Gene expression analysis
Liver tissue was shock-frozen in liquid nitrogen and stored at −80°C. RNA was purified by pegGOLD (peqLab, Erlangen, Germany) using standard protocols, and complementary DNA was generated from 1 μg RNA using a complementary DNA synthesis kit (Roche, Basel, Switzerland).18 Quantitative real-time PCR was performed using SYBR green reagent (Invitrogen, Darmstadt, Germany). Reactions were done twice in quintuples and β-actin values were used to normalise gene expression. Primer sequences are available upon request.
Cytokine and chemokine measurements
Liver tissue was shock-frozen in liquid nitrogen and stored at −80°C. Protein extracts were obtained using standard protocols. Protein concentrations were measured by extinction-based procedure. Aliquots of 50 μl were used for cytometric bead arrays (BD), according to the manufacturer's protocol.17
Transposon-mediated hepatic ovalbumin (Ova) expression
The pT/Caggs vector and transposase-encoding vector pPGK-SB13 have been described previously.19 To generate pT/Caggs-OVA the full-length sequence of ovalbumin (1.2 kb) was PCR amplified using the following primers: forward 5′-GGATCGAACGCGTATGGGCTCCATCGGCGCAGC-3′ (MluI overhang) and reverse 5′-GGATCGAACCGGTTTAAGGGGAAACACATCTGCCAAAGAAGAGAACG-3′ (AgeI overhang). The PCR-product obtained was subcloned into an AscI/AgeI digested pT/Caggs to obtain pT/Caggs-OVA. For transposon-mediated intrahepatic expression of ovalbumin, animals received a 5:1 molar ratio of transposon pT/Caggs-OVA to transposase-encoding plasmid pPGK-SB13 (12 μg total DNA). DNA was suspended in saline solution at a final volume of 10% of the animal's body weight and injected via the tail vein in <10 s.
Detection of ovalbumin-specific antibodies
An enzyme-linked immunosorbent assay (ELISA) was used for the detection of Ova-specific antibodies in the serum of mice. Briefly, 96-well flat-bottom microtitre plates were coated with 5 μg/ml of Ova (grade VII, Sigma). Plates were incubated overnight at 4°C, then washed and blocked with blocking solution (3% bovine serum albumin in phosphate-buffered saline) for 1 h at 37°C. Serum samples were diluted 1:100 in blocking solution and incubated for 1 h at 37°C. Control wells (blank) were filled with blocking solution only. After washing biotinylated anti-mouse IgM or anti-mouse IgG, respectively (both purchased from Sigma), were added to the wells and plates were incubated for 1 h at 37°C. After another wash, peroxidase-conjugated streptavidin (BD Pharmingen) was added to the wells for 30 min at 37°C. After a final wash, enzymatic reactions were developed using a substrate for peroxidase, 2,2′-azino-bis(3-ethylbenzthiazoline-6-sulphonic acid) (Roche Diagnostics), supplemented with 0.03% H2O2 (Sigma). After 30 min the reaction was terminated with a stop solution (1% sodium dodecyl sulphate). The absorbance was measured at 405 nm (OD405) using Infinite M200 reader (Tecan, Männedorf, Switzerland) and analysis was performed using MagellanTM (version 6.6) software (Tecan).
Statistical analysis from animal experiments
All experimental data are expressed as mean±SD. Differences between more than two groups were assessed by analysis of variance, and between two groups by two-tailed unpaired Student t test (GraphPad), except where indicated otherwise.
Impact of immune-related gene expression for survival of patients with HCC
To investigate the prognostic relevance of immune system related KEGG (Kyoto Encyclopedia of Genes and Genomes, http://www.genome.jp/kegg) signalling pathways in human HCC, we analysed a dataset containing 139 human genome-wide HCC tissue microarrays20 ,21 using BRB array tools (http://linus.nci.nih.gov/BRB-ArrayTools.html; version 3.8.0). Data were normalised using the median over the entire array. After normalisation, all genes with a percentage of missing data exceeding 5% were excluded. Then, the remaining genes were filtered for the individual KEGG signalling pathway to be analysed. Unsupervised hierarchical clustering using centred correlation and average linkage, split the human dataset into two subgroups A and B. For comparing the difference in survival between the two subgroups A and B, we performed Kaplan–Meier survival analysis and log-rank test using the MedCalc software packages (http://www.medcalc.be).
In addition, we applied the ‘Survival Analysis Tool for finding genes whose expression is correlated with survival time’ supplied by BRB Array Tools to the immune system-related KEGG signalling pathways. This tool is based on Cox proportional hazards regression analysis and identifies single genes whose actual log expression values correlate with survival time by parametric t test as well as by Kaplan–Meier survival analysis. The threshold was set to p<0.05 combined with 10 000 permutations to control for false discoveries (p<0.05). Furthermore, all genes associated with the particular KEGG pathway were included and genes were not subject to previous filtering. We only present genes for which the parametric p value as well as the Kaplan–Meier survival analysis showed significant differences in survival for that particular gene. For each gene the geometric mean expression ratio in the prognosis relevant subgroups A and B was calculated and their resulting fold change was used to rank the genes. The top five genes of each of the KEGG pathways analysed are presented, their degree of regulation has been colour coded.
Furthermore, the top five genes of each pathway were validated in an independent HCC patient population, using an open-source database.22 Associations between gene expression and clinical characteristics of HCC development and progression were obtained from the iCOD database.22 When multiple microarray spots in the database represented single genes, we selected those spots with the most significant clinical associations.
Results
Chemically induced hepatocarcinogenesis triggers liver inflammation with intrahepatic accumulation of macrophages and cytotoxic T cells
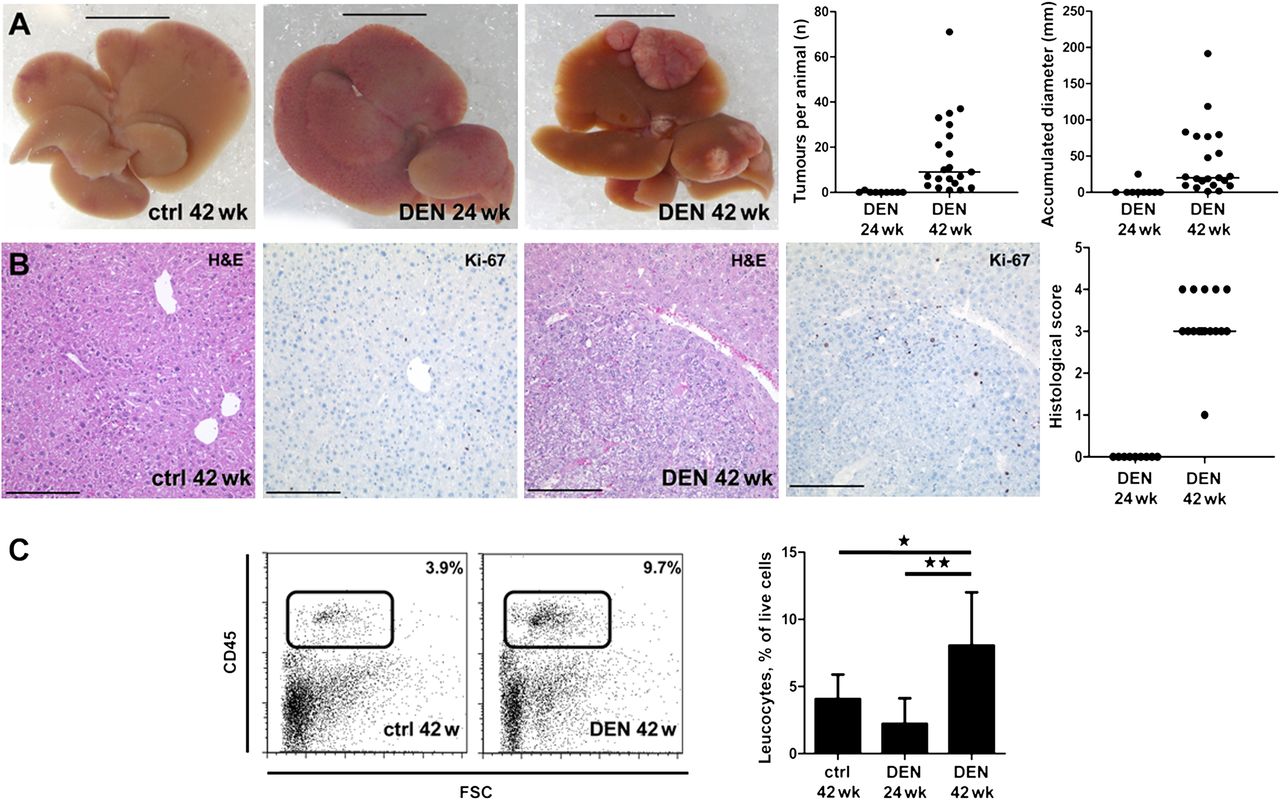
To investigate inflammatory responses induced by HCC, 14-day-old C57BL/6 male mice were injected with the chemical carcinogen DEN, and tumour formation was assessed 24 and 42 weeks later. Only one animal showed a visible nodule at the liver surface at 24 weeks after DEN administration, but the majority of mice had developed macroscopically detectable neoplasias at 42 weeks (figure 1A). Tumours were verified by Ki-67 staining and histopathological grading according to a UICC-adapted scale (figure 1B, supplementary table 1). Interestingly, DEN-induced hepatocarcinogenesis was associated with a significant increase in intrahepatic immune cells (figure 1C).

Chemically induced hepatocarcinogenesis triggers liver inflammation. Hepatocarcinogenesis was chemically induced by a single injection of diethylnitrosamine (DEN) intraperitoneally in C57BL/6 wild-type mice 14 days post partum. Livers were analysed 24 or 42 weeks after DEN. (A) Macroscopic appearance of liver tumour development (black bars: 10 mm). Nodules visible at the surface of each lobe were counted using a macroscope. The maximal diameter of each nodule was assessed with calibrated software and added to obtain the accumulated diameter (n=9 for 24 weeks, n=21 for 42 weeks). (B) Standard haematoxylin and eosin (H&E) stainings were scored by an expert pathologist, Ki-67 stainings indicated increased cell proliferation in tumour nodules. Black bars indicate 200 μm. (C) Intrahepatic infiltration of immune cells was assessed by flow cytometry for the pan-leucocyte marker CD45. *p<0.05, **p<0.01. Control animals (n=6) were age-matched (44 weeks old) for long-term DEN treatment.
We thus analysed the inflammatory influx from tumour-bearing and control liver tissue in more detail. By comprehensive FACS analyses, a clear accumulation of intrahepatic macrophages was seen (figure 2A,B). Immunohistochemical staining showed that these macrophages preferentially accumulated peritumourally (data not shown). Neutrophils and natural killer (NK) cells did not differ significantly between DEN-treated and control livers (figure 2). For the adaptive immune cells, a significant increment of CD8 cytotoxic T cells was detected in tumour-bearing livers, whereas intrahepatic CD4 T cells or CD19 B cells remained unaltered (figure 2). Similar results were obtained when not the relative leucocyte proportions, but absolute immune cell numbers were compared (data not shown). Moreover, DEN-induced hepatocarcinogenesis also triggered the accumulation of cancer-associated myeloid-derived suppressor cells (MDSCs) in bone marrow and spleen (supplementary figure 2), which are characterised by high expression of CD11b and Gr1 in mice23 and have been also reported in patients with HCC.24

Diethylnitrosamine (DEN)-induced hepatocarcinogenesis promotes intrahepatic accumulation of macrophages and cytotoxic T cells. Hepatocarcinogenesis was chemically induced by a single injection of DEN intraperitoneally in wild-type mice 14 days post partum. Livers were analysed 42 weeks after DEN (n=21) or in age-matched controls (n=6). (A) Representative FACS plots for different intrahepatic immune cell populations, pre-gated on CD45 and living (Hoechst-dye negative) cells. Staining compounds used for identification of immune cell populations were Ly6G−CD11bF4/80 for macrophages, SSChighLy6G for neutrophils, CD3CD4 or CD3CD8 for T lymphocytes, NK1.1CD3−CD4− for NK- and SSClowCD3−B220 for B cells. (B) Quantification of the different leucocyte subsets was based on FACS (see plots in A). Afterwards, relative immune cells counts by FACS were normalised to total intrahepatic cells or to total lymphocytes. *p<0.05.
The distinct intrahepatic accumulation of macrophages and CD8 T cells in tumour-bearing mice led us to hypothesise that the tumour or its surrounding tissue produced inflammatory mediators that would attract distinct immune cells. Indeed, we found a significant increase in protein levels of the macrophage attractant CCL2 (MCP-1) and of the T cell-attractive chemokines CCL3 (MIP1α), CCL4 (MIP1β), CCL5 (RANTES) and CXCL9 (MIG) in cancer-bearing livers (figure 3). Other chemokines simultaneously analysed by a multiplex assay were not differentially regulated at the protein level between DEN-treated and control livers (not shown). Interestingly, the T cell attractants CCL3, CCL4 and CCL5,25 in particular, were already strongly induced in livers at 24 weeks after DEN (figure 3B,D), in which transformed hepatocytes, but no tumours were seen (figure 1), corroborating the suggestion that the inflammatory response is induced early during hepatocarcinogenesis and is a highly regulated process.

Diethylnitrosamine (DEN)-induced hepatocarcinogenesis increases intrahepatic chemokine expression. Hepatocarcinogenesis was chemically induced by a single injection of diethylnitrosamine (DEN) intraperitoneally in wild-type mice 14 days post partum. Chemokine protein concentrations were measured from livers by multiplex cytometric bead assays at indicated time-points after DEN or in controls. The macrophage-attractive CCL2 (monocyte chemoattractant protein-1 (MCP-1) A) and the T cell-attractive chemokines CCL3 (MIP-1α, B), CCL4 (MIP-1β, C), CCL5 (RANTES, D) and CXCL9 (MIG, E) were significantly upregulated in DEN-treated livers on a protein level. *p<0.05, **p<0.01, ***p<0.001.
Finally, to test whether these immune cell compartments were also present in human HCC, we performed immunohistochemical analyses. Remarkably, and in line with current literature,26 liver tissue from patients with HCC showed pronounced inflammatory reactions compared with control livers, with innate as well as adaptive immune cells being found diffusely scattered throughout malignant and non-malignant hepatic tissue (supplementary figure 3). Of note, similar to DEN-induced murine hepatocarcinogenesis, macrophages (supplementary figure 3B) and CD8 cytotoxic T cells (supplementary figure 3E) were increased in human HCC as well, in direct contrast to healthy control liver. However, it has been previously noted that the distribution of immune cells in human HCC is heterogeneous between tumourous, peritumoural and non-tumourous areas.27 Collectively, our experiments demonstrated that DEN-induced hepatocarcinogenesis provoked distinct inflammatory reactions in the liver. Therefore, we next conducted experiments to elucidate the impact of those immune alterations on tumour progression.
Enhanced macrophage accumulation in chemokine scavenger receptor D6-deficient mice does not influence tumour progression
Based on the tumour-associated over-expression of inflammatory chemokines in the DEN model (figure 3), we next investigated the possible link between chemokine expression and tumour progression on a functional level. The atypical chemokine receptor D6 acts as a scavenger of inflammatory CC-chemokines (eg, CCL2, CCL3, CCL4 and CCL5), thereby limiting inflammatory responses in vivo.11 We used D6−/− mice in the DEN model to test if an enhanced chemokine response would promote or limit hepatocarcinogenesis. After hepatic tumour induction 42 weeks after DEN administration, D6−/− mice showed significantly higher numbers of intrahepatic macrophages than WT animals, while intrahepatic neutrophils were reduced in DEN-treated D6−/− mice (figure 4A,B). Macrophages were increased in number, and also showed significantly higher expression rates of Gr-1/Ly6C (figure 4A), which is indicative of a more proinflammatory phenotype.28 Additionally, CD4 T-helper cells were increased, whereas the relative (figure 4C) and absolute counts (not shown) of the other lymphocyte populations were unaltered. However, despite the pronounced macrophage accumulation, macroscopic tumour formation and tumour size were similar in DEN-treated D6−/− and WT mice (figure 4D). Also on histological analysis, hepatic tumours did not differ microscopically in D6−/− from WT mice (figure 4D,E), with the exception that some D6−/− mice displayed patches of mononuclear cells peritumourally (figure 4E, arrow). Of note, D6−/− mice showed significantly higher proportions of MDSCs in spleen and bone marrow under homoeostatic conditions than wild-type animals. But whereas in wild-type mice MDSCs would increase considerably and catch up with their knockout counterparts upon tumour growth, in D6−/− mice these populations only slightly expanded in the spleen and even decreased in the bone marrow (figure 4F,G). Taken together, these findings indicated that neither enhanced accumulation of intrahepatic macrophages in tumour-bearing livers nor an increased pre-existing extrahepatic MDSC compartment significantly influenced tumour formation or progression in DEN-induced hepatocarcinogenesis.

Enhanced macrophage accumulation and pronounced pre-existing extrahepatic myeloid-derived suppressor cell (MDSC) compartment in chemokine scavenger receptor D6-deficient mice does not influence tumour progression. Hepatocarcinogenesis was chemically induced by a single injection of diethylnitrosamine (DEN) intraperitoneally in wild-type (WT; n=21) or D6−/− mice (n=16) 14 days post partum. Livers were analysed 42 weeks after DEN. (A) Macrophage infiltration was analysed by immunohistochemistry for F4/80 (brown colour, left) and FACS (right), representative data are shown. The histogram shows Gr-1-expressing cells within the hepatic macrophage population. (B–C) Quantification of neutrophil, macrophage (B) and lymphocyte subset (C) accumulation in DEN-treated livers of WT and D6−/− mice. Tumour development was not shown to differ either by macroscopic or histological (D) analysis. In some DEN-treated D6−/− animals, inflammatory patches were occasionally noted (arrow) (E). (F–G) MDSCs were quantified in spleen (F) and bone marrow (G). *p<0.05, **p<0.01, ***p<0.001.
Distinct functions of T and B lymphocytes on tumour formation and progression in DEN-induced hepatocarcinogenesis
Based on our findings of increased intrahepatic leucocytes in tumour-bearing mice and unaltered tumour development in mice with enhanced innate immune cell accumulation, we next investigated the impact of adaptive immunity on liver cancer progression in the DEN model. To examine this in a broad manner, we applied the DEN model to Rag1-deficient mice, which lack both mature T and B cells (supplementary figure 4).29 Upon challenge with DEN, Rag1−/− mice developed strikingly more hepatic tumours (figure 5A). Macroscopically visible nodules were already detectable at 24 weeks after DEN in the majority of Rag1−/−, but not in WT mice (figure 5B). At 42 weeks after DEN administration, significantly more tumour nodules were detected in Rag1−/− mice (figure 5B). Tumours in Rag1−/− mice tended to grow more aggressively, as assessed by histological grading (figure 5B). Owing to the complete lack of T and B cells, the intrahepatic leucocyte infiltrate in Rag1−/− mice was significantly attenuated and, as expected, consisted mainly of neutrophils and macrophages (figure 5C). Nevertheless, the absolute number of intrahepatic neutrophils was significantly reduced in DEN-treated Rag1−/− in comparison with WT mice (figure 5C).

Adaptive immunity critically controls hepatic tumour formation and progression. (A) Macroscopic phenotype of the immune cell-deficient mouse strains, 42 weeks after diethylnitrosamine (DEN; black bars: 10 mm). (B) Rag1−/− mice show significantly earlier and more frequent tumour nodule formation than wild-type (WT) mice. Tumour grading tended to be more aggressive by pathologist's scoring. (C) Owing to a lack of T and B cells, intrahepatic leucocytes are reduced in Rag1−/− mice. Hepatic neutrophils are also lower, while macrophages are unaltered. (D) Tumour phenotype at 42 weeks after DEN in WT (n=21), Rag1−/− (n=10), Igh6−/− (n=5) and MHCII−/− (n=5) mice. (E) In Igh6−/− mice, tumours >10 mm in diameter were more frequently observed. *p<0.05, **p<0.01, ***p<0.001 (in E: two-sided Fisher's exact test).
To further dissect the functional contribution of T and B cells during cancer development, Igh6−/− mice and MHCII−/− mice were subjected to DEN-induced hepatocarcinogenesis. Igh6−/− mice, also called μMT mice, lack B cells, owing to a mutated IgM transmembrane domain preventing B cell maturation, whereas MHCII−/− mice have functional B cells and CD8 T cells, but an impaired CD4 T cell development and may also have impaired functions of M2 macrophages (supplementary figure 4).30 Comparing these different genotypes, Rag1−/− mice surpassed all other mouse strains with respect to the number of tumour nodules at 42 weeks after DEN (figure 5D). Interestingly, also Igh6−/−, but not MHCII−/− mice developed more tumours than WT animals in the DEN model (figure 5D). B cell-deficient Igh6−/− mice displayed, on average, larger tumours than WT, Rag1−/− or MHCII−/− mice (figure 5D). For the impact of CD4 T cells, MHCII−/− mice tended to develop slightly increased numbers of nodules while tumours remained rather small, but the differences in WT mice did not reach statistical significance. These experiments indicate that adaptive immune cells critically control hepatocarcinogenesis.
Increased detection of antibodies directed against antigens expressed in hepatocytes of DEN-treated mouse livers
The experiments using B cell-deficient mice indicated that B cells are involved in limiting tumour growth. We thus hypothesised that the tumour microenvironment leads to activated B cells that are specific for liver-derived antigens. This assumption was supported by an increased expression of CD81 and BLNK, two major functional mediators of B cell activation and immunoglobulin (Ig) production,31 ,32 in tumour-bearing livers, dependent on the individual tumour burden (figure 6A). To functionally test whether the DEN-related hepatic microenvironment allows for an increased induction of humoural immune responses against antigens in hepatocytes, we subjected DEN-treated and control mice to hydrodynamic injections with transposable elements encoding the Ova model antigen. This procedure allows for a stable delivery of the transposon-encoded transgene. On day 16 and subsequently on day 28 serum of mice was analysed for the presence of Ova-specific IgM and IgG antibodies. Strikingly, both Ova-specific Igs were increased in those mice which had been treated with DEN at an early age (figure 6B). Therefore our functional data shows that DEN-induced changes in the hepatic microenvironment and/or the inflammatory response allow for an increased efficiency in triggering humoural immune responses against antigens expressed in hepatocytes.

B cell activation and functionality is enhanced in tumour-bearing livers. Hepatocarcinogenesis was chemically induced by a single injection of diethylnitrosamine (DEN) intraperitoneally in wild-type (WT) mice 14 days post partum. (A) Expression levels of B cell activation-associated genes CD81 and BLNK were quantified by real-time qPCR in mice with low (<5 nodules) and high (>30 nodules) tumour (tu) burden. Data are normalised to β-actin and expressed as fold induction compared with matched healthy livers. (B) 20 Weeks after DEN or saline injection, DEN- and saline-treated control mice were subjected to hydrodynamic injection of ovalbumin-expressing transposon–transposase constructs, leading to continuous expression of the transgene (ovalbumin) by hepatocytes. Antigen-specific IgM and IgG were measured by ELISA in serum samples obtained 16 and 28 days thereafter; n=3 mice for control and n=4 mice for DEN injection were analysed at each time point.
Activation of adaptive immunity-related pathways affect survival of patients with HCC
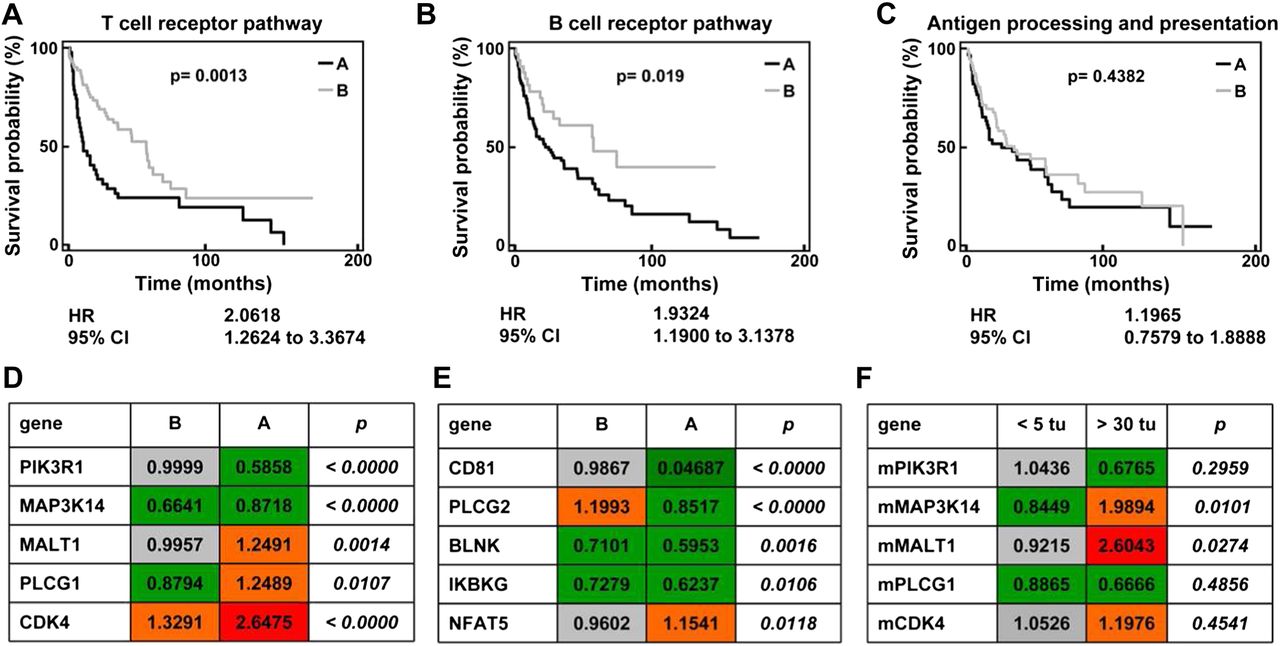
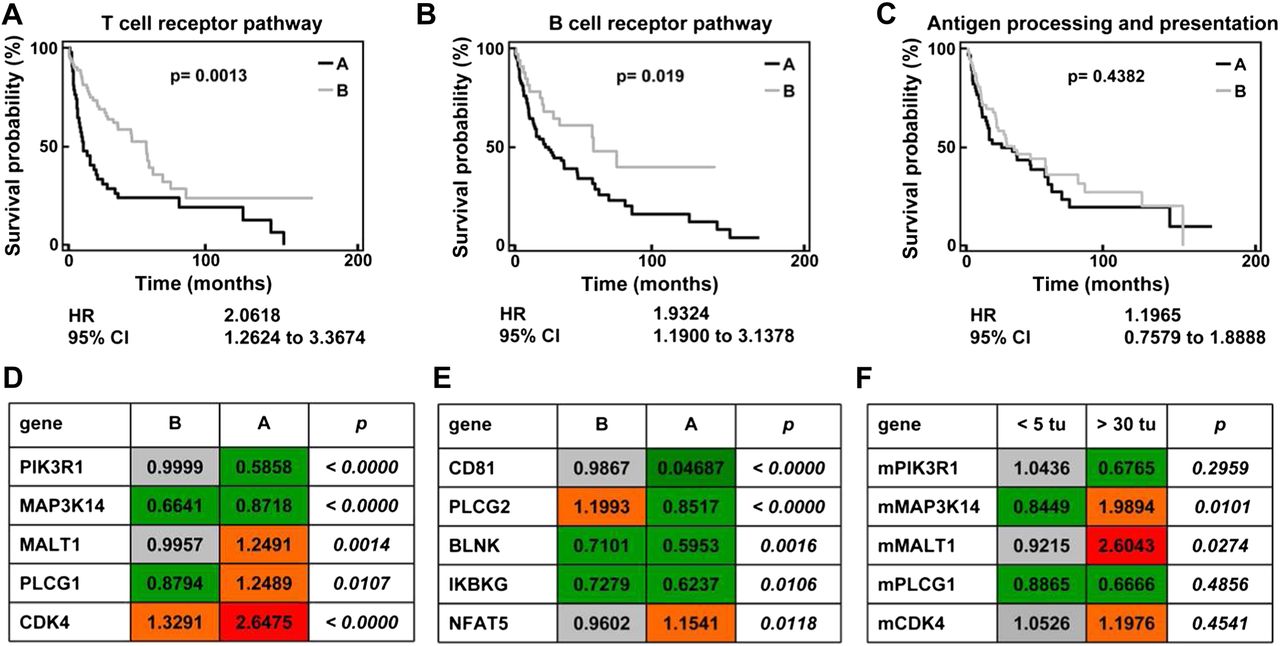
These experiments demonstrated that the activation of adaptive immunity in the liver is an essential protective mechanism in DEN-induced hepatocarcinogenesis. In order to translate the relevance of our findings into human HCC, we analysed gene array data from 139 HCC samples20 ,21 with respect to pathways of innate and adaptive immunity. Characteristic signalling pathways were chosen from the KEGG pathway maps (http://www.genome.jp.kegg). After basic filtering for genes with available expression data in >95% of HCC array datasets, 23 of 103 genes relevant to T cell receptor signalling and 18 of 67 genes relevant to B cell signalling were eligible for further analysis. Unsupervised clustering split the 139 HCC cases into two subgroups (A and B), which were then analysed by Kaplan–Meier plots for differences in survival. Differentially regulated T cell-related (figure 7A) and B cell-related genes (figure 7B) were found to be of prognostic relevance in HCC cases. The combined hazard ratio calculated from the gene clusters derived from the T cell or B cell KEGG pathway map was 2.06 for the T cell and 1.93 for the B cell pathway (figure 7A,B). On the other hand, 41 genes from the KEGG pathway ‘antigen processing and presentation’, which comprises genes involved in macrophage and dendritic cell functionality, could not significantly discriminate prognosis in human HCC (figure 7C).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Activation of adaptive immunity-related pathways affect survival of patients with hepatocellular carcinoma (HCC). Unsupervised clustering analysis of 139 cases of human HCC based on genes relevant to KEGG pathway ‘T cell receptor signalling’ (A), ‘B cell receptor signalling’ (B) or ‘Antigen processing and presentation’ (C) defined two subgroups of HCC: A (black line) and B (grey line). These subgroups were then analysed for differences in patient survival using Kaplan–Meier plots of 85 out of the 139 cases for which clinical data were available. (D–E) Advanced statistical analysis was performed to identify single genes among these KEGG pathways whose expression is correlated with survival time. For this analysis the whole set of genes per pathway as defined by KEGG was used. For 17/103 T cell receptor- and for 14/67 B cell receptor-associated genes a correlation with survival time was identified (parametric p value <0.05 and permutation p value ≤0.05). The tables provide the geometric mean of gene expression data of the top five most significantly and differentially regulated T cell receptor-associated (D) and B cell receptor-associated (E) genes whose single gene expression correlates with survival time. The p values describe the degree of significance with which the gene expression discriminates between HCC subgroup A and B (t test). (F) The regulation of the top five most significantly and differentially regulated T cell receptor-associated genes was tested by quantitative PCR in murine liver tissue after diethylnitrosamine treatment. Animals with high tumour burden (more than 30 nodules) were compared with mice with lower tumour burden (<5 nodules). Geometric mean of fold expression normalised to healthy age-matched control animals is presented (please compare with D).
In a more detailed analysis, which was based on Cox proportional hazards regression, single genes whose expression is correlated with survival time were identified. Among the unfiltered total group of 103 T cell receptor pathway genes, 17 were found to be associated with survival (parametric p value <0.05 and permutation p value ≤0.05). Among the unfiltered total group of 67 B cell pathway genes, 14 were found to be associated with survival (parametric p value <0.05 and permutation p value ≤0.05). We focused on the five most significantly and differentially regulated genes from each pathway between patients with good or poor prognosis (the full list of genes and their differential expression is provided as supplementary table 2). While some of the T cell-related genes were strongly upregulated in samples from patients with poor prognosis (figure 7D), a remarkable downregulation of important B cell activating genes was noted in patients with unfavourable outcome (figure 7E), indicating that B cells may also be important in human HCC to limit tumour progression.
In order to validate the importance of regulated T or B cell-associated genes, we tested these genes in an independent cohort of patients with HCC using an open-source database (supplementary table 3). However, survival data were not available for these patients with HCC,22 but clinical characteristics such as tumour stage and burden, biomarkers and invasiveness served as surrogate markers for prognosis. Furthermore, the T cell receptor pathway genes from human HCC arrays were also tested in tumour-bearing livers from DEN-treated mice. Interestingly, the genes from the T cell pathway were generally regulated in a similar direction in DEN-treated mice to that in human HCC database (figure 7F). For example, PIK3R1 tended to be downregulated in mice with more than 30 nodules compared with animals with fewer than five tumours, while downregulation of PIK3R1 was prognostic for poor survival in humans. Also, MALT1 was significantly upregulated in mice with higher tumour burden and humans with poor prognosis. Collectively, these data indicate that immunological mechanisms identified in the murine DEN model may be relevant also for progression of human liver cancer.
Discussion
HCC is regarded as a prototype of ‘inflammation-associated cancer’, because it regularly originates within chronically inflamed tissue.10 However, it is known from many other tumour entities that inflammation caused by malignant tissue is an important component of the tumour microenvironment. Thus, the occurrence of tumour-associated macrophages has been linked to immune suppression, angiogenesis and tumour progression,33 whereas adaptive immune cells may be important in cancer immunosurveillance, as transformed cells can be eliminated by immune-mediated actions.34
In the case of HCC, it is challenging to assess the specific role of tumour-associated inflammation due to the underlying inflamed hepatic environment. Several mouse models have been developed to unravel the link between inflammation and hepatocarcinogenesis.35–37 In these models, abrogation of the immune response—for example, by crossing lymphotoxin-expressing transgenic mice on a T/B cell-deficient (Rag1−/−) background, generally abrogates liver cancer formation,36 suggesting that the presence of adaptive immune cells is required for tumour progression. However, the major difference from the DEN model is that transgenic lymphotoxin expression in the liver results in a progressive hepatic inflammation, in which tumours arise secondarily.36 This tissue-destructive inflammation is undoubtedly not induced by or directed against the HCC. Thus, in order to investigate the inflammation induced by HCC, we chose the well-established and widely used murine model of HCC induction by DEN in male mice.8 By comparative genomic analyses, gene expression patterns seen in tumours in the DEN model were found to be similar to human HCC tissue of patients with poor prognosis.38 Of note, although considered a non-inflammatory liver cancer model,10 DEN-induced hepatocarcinogenesis has been perceived to be influenced by inflammatory signals. For instance, loss of IKKβ in hepatocytes, which prevents NF-κB activation, augments DEN-induced death, thereby potentiating carcinogenesis.3
Similar to our own analysis of human samples and observations reported in the literature from human HCC,26 DEN-induced liver cancer in mice was accompanied by a substantial leucocyte infiltration. DEN-induced hepatocarcinogenesis resulted, especially, in increasing numbers of hepatic macrophages and CD8 T cells. Tumour-associated macrophages have been linked to HCC invasiveness39 and with poor prognosis in various tumour entities probably owing to an immunosuppressive phenotype.33 ,40 HCC-associated lymphocytes have been described as comprising mainly T cells in human tissue sections.26 In patients with HCC, high amounts of CD8 T cells within the tumour may indicate a favourable prognosis.27 ,41 In this regard, the observed inflammatory influx of leucocytes induced by DEN-induced liver cancer itself, and the enhanced fractions of macrophages and CD8 T cells in our model, are well in accordance with observations in human tissue. DEN-induced tumour formation was further associated with substantial and marked induction of proinflammatory chemokines within the livers of mice. Interestingly, we found that both macrophage-associated (CCL2) and T cell-associated chemokines (CCL3-CCL5, CXCL9) were specifically induced in tumour-bearing livers.25 Moreover, tumour-bearing mice showed remarkable alterations of systemic immune cells—specifically, increased fractions of Gr-1CD11b MDSCs in bone marrow and spleen. MDSCs have been described in patients with liver cancer and are thought to exert tumour-promoting functions by inhibiting NK cell activity and induction of regulatory T cells.24 ,42
Based on the distinct hepatic accumulation of macrophages and T cells in DEN-treated animals, we conducted experiments to functionally dissect the role of these immune cell populations for tumour progression (table 1). Hypothesising that tumour-associated macrophages would promote tumour growth, we induced tumours in D6−/− mice, in which the scavenging of inflammatory chemokines was impaired.11 Of note, the atypical chemokine decoy receptor D6 has been found to suppress the development of chemically induced skin tumours, as D6−/− mice show an increased susceptibility to cutaneous tumour development in response to chemical carcinogenesis.43 However, this skin cancer model was associated with constant inflammation, in contrast to our model of chemically induced liver cancer. In D6−/− mice, we observed an enhanced macrophage infiltration upon DEN treatment, but no difference from WT mice in tumour development. Although we cannot exclude the possibility that the absence of this chemokine scavenging mechanism might functionally alter the observed macrophages, these experiments indicate that the increased numbers of macrophages in DEN-treated livers do not promote hepatic cancer progression by itself. Moreover, the increased levels of MDSCs seen in D6−/− mice under homoeostatic conditions did not significantly influence tumourigenesis, probably because WT mice expand this population rapidly upon tumour induction.
Macroscopic phenotype of the diethylnitrosamine liver tumour model in the different knockout mouse strains in comparison with wild-type animals
In order to assess the functional contribution of adaptive immunity in the DEN model, we investigated Rag1−/− mice, which are deficient for T and B cells. Rag1−/− mice showed a striking phenotype with markedly enhanced growth in the number and mass of tumours. Furthermore, tumours grew faster, forming visible nodules at an age of 24 weeks and were less differentiated. This is in accordance with the concept of equilibrium of occult cancer, which describes a T cell-dependent control of spots of very few transformed cells without eradicating them.34 Possibly, the adaptive immune cells are involved in clearing senescent ‘premalignant’ hepatocytes, as the clearance of senescent hepatocytes is an important mechanism of tumour surveillance in liver cancer models.19 ,44 However, we cannot exclude the possibility that additional alterations of the inflammatory reaction, as reflected by reduced intrahepatic neutrophil infiltration in Rag1−/− mice, also affect tumourigenesis in the absence of T cells.
Although immune suppression is widely accepted as tumour promoting in non-HCC cancers,5 our findings sharply contrast inflammatory liver cancer models, in which deletion of T and B cells abrogate tumour formation.36 To further elucidate the functional roles of the missing T and B cells in Rag1−/− mice, we also subjected Igh6−/− and MHCII−/− mice to treatment with DEN. Lack of B cells resulted in fewer but larger tumours than found for Rag1−/− mice, suggesting that B cells limit growth of established tumours, while T cells prevent initial nodule formation from transformed hepatocytes. Such T cell-dependent immune surveillance has been recently described in a different, oncogene-dependent liver cancer model.19 In contrast, no significantly increased tumourigenesis was found in MHCII−/− mice. In our study MHCII−/− mice were initially used as a model for impaired CD4 T cell function. However, interpretation of the obtained data is complicated by the fact that DEN-induced hepatocarcinogenesis depends on functional proinflammatory macrophages in the liver.3 As has been recently described, production of proinflammatory cytokines and interferon by macrophages of MHCII−/− mice is impaired,30 further DEN studies in CD4−/− mice are needed to examine the contribution of CD4 T cells for immune surveillance of HCC. Interestingly, recent work by Kang et al showed that the immune surveillance of precancerous senescent hepatocytes is impaired in CD4−/− mice in different HCC mouse models,19 thus further emphasising the need for additional DEN studies in CD4−/− mice.
In contrast to our results in Igh6−/− mice, tumour growth-promoting roles of B cells have been described previously, especially by producing lymphotoxins45 and by skewing the differentiation of tumour-surrounding macrophages towards an M2-like state through Fcγ receptor-dependent signalling.46 However, this B cell function was only demonstrated in inflammatory tumour models.46 Therefore we can hypothesise, that B cells exert a suppressive function on tumour growth in liver cancer, when a tumour is established and when inflammation is solely triggered by the tumour itself. Congruent with this assumption, cytotoxic antibodies and Fcγ receptor-dependent activation of leucocytes that exert antitumour activity have recently been studied.47 In line with this, we observed a tumour-associated B cell activation with antigen-specific immunoglobulin production during hepatocarcinogenesis, suggesting functionally relevant humoural antitumour immune responses in the DEN model.
Our experiments demonstrated that DEN-induced liver cancer led to an inflammatory hepatic microenvironment, in which adaptive immune cells essentially control early tumour formation as well as growth of established tumours. This led us to hypothesise that the activation of adaptive immune response might also be relevant in human HCC progression. In patients with liver cancer, gene profiling allows identification of prognostic patterns of gene regulation in hepatic tissue.21 ,48 We thus analysed earlier gene array datasets of regulated genes in 139 HCCs21 for genes relevant to the adaptive immune system using the KEGG pathway maps. We found that T and B cell-related genes from these pathways clustered in HCC samples and defined groups of distinct favourable or poor prognosis. Remarkably, antigen presentation-related pathways were not indicative of survival of patients with HCC in this analysis. Some of the gene regulations we observed, might have occurred within the malignant cells and not in infiltrating leucocytes—for instance, PIK3RI, which has been attributed with tumour-suppressive properties by regulating growth factor signalling in liver cells and liver cancer49 and may not be directly related to T cell functionality.50 However, other genes are clearly associated with leucocytes—for example, the T cell receptor associated gene MALT1 or CD81 and BLNK, which interact with B cell and T cell receptor complexes.32 ,51 In particular, the downregulation of B cell-activating genes was found to indicate unfavourable outcome and was in line with the observations of the murine DEN model. For example, phospholipase C (PLC)-γ2 is required for efficient formation of germinal centre and memory B cells.52 Given the hypothesis that PLC-γ2 conveys a survival signal to the germinal centre and memory B cells, required for a productive secondary immune response, the observed downregulated PLC-γ2 levels in patients with HCC with a poor prognosis might indicate an impaired memory B cell response in advanced HCC. However, PLC-γ2 might also represent a functional bridge between adaptive and innate immune responses due to PLC-γ2-dependent NK cell cytotoxicity, as shown in a mouse model of viral-infected and malignant cells.53
It was shown previously that the DEN tumour model genetically resembles human HCC of poor prognosis,38 but we nevertheless investigated these genes further and determined their regulation in tumour-bearing mice, which were divided into subgroups of higher and lower tumour burden. It was intriguing to find striking similarities in the regulation of T cell-associated genes between human HCC and the murine DEN model. Moreover, immunosuppressive medication has recently been associated with HCC progression in patients after re-transplant for hepatitis C,19 and lymphocyte-infiltration related genes such as chemokines (CCL2, CCL5, CXCL10) were identified as important indicators of favourable prognosis in human HCC in two independent cohorts.54 Nevertheless, the exact pathogenic contribution of the dysregulated genes in hepatocarcinogenesis remains to be elucidated by further functional studies.
Our study thereby emphasises that HCC arises within inflamed tissue, and also inherently triggers local hepatic and systemic inflammatory responses, which in consequence suppress tumourigenesis. Thus, future therapeutic approaches for HCC may be directed against tumour-promoting inflammatory reactions, and also towards augmenting HCC-induced antitumoural adaptive immune cell functions.
Acknowledgments
This article is dedicated to Professor Michael Manns (Hannover) on the occasion of his 60th birthday. The authors thank Aline Müller for excellent technical assistance and Professor Snorri S Thorgeirsson (National Cancer Institute, Bethesda, Maryland, USA) for providing the gene array datasets from patients with HCC. The HCC survival analyses were performed using BRB-ArrayTools developed by Dr Richard Simon and BRB-ArrayTools Development Team.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Download Supplementary Data (PDF) - Manuscript file of format pdf
- Download Supplementary Data (PDF) - Manuscript file of format pdf
- Download Supplementary Data (PDF) - Manuscript file of format pdf
- Download Supplementary Data (PDF) - Manuscript file of format pdf
- Download Supplementary Data (PDF) - Manuscript file of format pdf
- Download Supplementary Data (PDF) - Manuscript file of format pdf
Footnotes
-
Funding This work was supported by the German Research Foundation (DFG Ta434/2-1 to F.T., DFG SFB/TRR 57), by the Start-program and Interdisciplinary Centre for Clinical Research (IZKF) at RWTH-Aachen University.
-
Competing interests None.
-
Provenance and peer review Not commissioned; externally peer reviewed.
-
Data sharing statement All primary data are available upon request. SAL can be contacted about the D6-deficient mice, AH and LZ about the OVA-expressing plasmid.