Article Text

Abstract
Objective Therapeutically targeting lymphocyte adhesion is of increasing relevance in IBD. Yet, central aspects of the action of antiadhesion compounds are incompletely understood. We investigated the role of αEβ7 and α4β7 integrins and their blockade by vedolizumab and etrolizumab for trafficking of IBD T lymphocytes in an in vivo model of homing to and retention in the inflamed gut.
Design We explored integrin expression in patients with IBD by flow cytometry and immunohistochemistry, while regulation of integrins was studied in T cell cultures. The functional relevance of integrins was assessed by adhesion assays and a recently established humanised mouse model in dextran sodium sulfate-treated immunodeficient mice.
Results High expression of αEβ7 was noted on CD8+ and CD4+ Th9 cells, while α4β7 was expressed on CD8+, Th2 and Th17 cells. T cell receptor stimulation and transforming growth factor β were key inducers of αEβ7 on human T cells, while butyric acid suppressed αEβ7. In comparison to α4β7 blockade via vedolizumab, blockade of β7 via etrolizumab surrogate antibody superiorly reduced colonic numbers of CD8+ and Th9 cells in vivo after 3 hours, while no difference was noted after 0.5 hours. AEβ7 expression was higher on CD8+ T cells from patients with IBD under vedolizumab therapy.
Conclusions AEβ7 is of key relevance for gut trafficking of IBD CD8+ T cells and CD4+ Th9 cells in vivo and mainly retention might account for this effect. These findings indicate that blockade of αEβ7 in addition to α4β7 may be particularly effective in intestinal disorders with expansion of CD8+ and Th9 cells such as IBD.
- INFLAMMATORY BOWEL DISEASE
- INTEGRINS
- INTESTINAL T CELLS
- ADHESION MOLECULES
- ANTIBODY TARGETED THERAPY
Statistics from Altmetric.com
Significance of this study
What is already known on this subject?
The α4β7 integrin antibody vedolizumab blocks gut homing of regulatory and effector CD4+ T cells and is successfully used for clinical therapy in IBD.
αEβ7 integrin is believed to mediate retention of lymphocytes in or near the epithelium via interaction with E-cadherin.
The β7 integrin antibody etrolizumab targets α4β7 and αEβ7 integrins and is currently evaluated in phase III trials.
What are the new findings?
αEβ7 and α4β7 are differentially expressed on human T lymphocyte effector subsets both in the peripheral blood and the intestine.
T cell receptor stimulation and transforming growth factor β treatment increase αEβ7 expression especially on CD8+ lymphocytes.
In a recently established humanised mouse model, etrolizumab surrogate antibody is more potent than vedolizumab in reducing accumulation of human CD8+ and CD4+ Th9 cells in the inflamed gut, probably due to additional inhibition of αEβ7-mediated retention.
How might it impact on clinical practice in the foreseeable future?
β7 inhibition by antibodies such as etrolizumab might offer additional benefits for the treatment of IBD compared with α4β7 inhibition.
Introduction
A dysguided inflammatory response within the intestinal lamina propria is believed to be a cornerstone of the pathogenesis of IBD such as ulcerative colitis (UC) and Crohn's disease (CD).1–3 In particular, CD4+ and CD8+ T lymphocytes are critically involved in the responsible immunological network by secreting pro-inflammatory cytokines, orchestrating the function of other immune cells and causing direct damage to cellular or extracellular elements of the intestinal wall.4–6
Naive T lymphocytes are primed to become gut homing lymphocytes in the mucosa-associated lymphoid tissue. Here, dendritic cells (DCs) not only function as antigen-presenting cells, but also induce the expression of α4β7 integrin via production of retinoic acid.7 ,8 After re-entering the circulation, thus primed lymphocytes are able to access the intestinal lamina propria in a complex homing process9 that critically involves the interaction of α4β7 integrin with its endothelial receptor mucosal vascular addressin cell adhesion molecule 1 (MAdCAM-1), which is exclusively expressed on postcapillary venules in the intestine.10
Blockade of α4β7 integrin-mediated gut homing with vedolizumab has greatly increased the options in clinical therapy of patients with IBD.11 ,12 Mechanistically, we have recently shown that vedolizumab reduces colonic homing of both effector and regulatory CD4+ T lymphocytes.13 However, the impact of vedolizumab on CD8+ T cells and cytokine-producing CD4+ T lymphocyte subsets has not been addressed so far. Moreover, lymphocyte homing can potentially be mediated by other homing molecules like α4β1 integrin as well,14 suggesting that vedolizumab may not suppress homing of all lymphocyte subsets. Consistent with this concept, vedolizumab does not induce or maintain remission in a significant portion of patients.11 ,12
Several other compounds interfering with the gut homing process are currently under development. One of them is the monoclonal humanised rat anti-β7 integrin antibody etrolizumab, which has recently successfully been tested in a phase II study.15 As anti-β7 antibody, it not only interferes with the interaction of α4β7 with MAdCAM-1, but also inhibits binding of αEβ7 to its receptor E-cadherin. As E-cadherin is mainly expressed on epithelial cells, its ligand αEβ7 is believed to mediate epithelial retention of homed gut lymphocytes16 such as intraepithelial lymphocytes.17 Moreover, a decrease in the number of αE+ cells in the intestinal crypt epithelium was associated with a therapeutic response to etrolizumab treatment.15 Yet, functional in vivo studies assessing the importance of αEβ7 for trafficking of lymphocytes are lacking and the abilities of α4β7 versus β7 blockade to reduce lymphocyte accumulation in IBD have not been compared to date. Here, we explored homing and retention of human IBD CD4+ and CD8+ T cells upon α4β7 blockade via vedolizumab and β7 blockade via etrolizumab surrogate (etro-s) antibody in a humanised mouse model of colitis. Our findings indicate that etrolizumab has marked additional effects on CD8+ and Th9 trafficking compared with vedolizumab and might be a useful tool for future clinical therapy in IBD.
Methods
Materials and methods are available as online supplementary file. Online supplementary table S1 summarises the patients' clinical data.
Supplementary methods
Supplementary table
Results
AEβ7 and α4β7 integrins are differentially expressed on T cell subsets in the peripheral blood of patients with IBD
Although it is known that αEβ7 integrin is expressed on a subset of peripheral T cells,17 no quantitative analysis of αEβ7 expression in IBD has been performed so far. Moreover, αEβ7 and α4β7 expression on cytokine-producing lymphocyte subsets is largely unknown. We therefore measured the expression of αEβ7 and α4β7 on CD4+ and CD8+ T cells in patients with UC, CD and control donors. We found a significantly higher expression of αEβ7 and α4β7 on CD8+ than on CD4+ T cells regardless of the group analysed (figure 1A and online supplementary figure S1A,B). Furthermore, α4β7 levels on UC CD8+ T cells were found to be significantly higher than on CD CD8+ T cells, suggesting a particular relevance of this integrin for CD8+ T cell homing in the former disease. To define the potential differential target cells for vedolizumab and etrolizumab in the peripheral blood more clearly, we analysed α4 expression on αEβ7+ cells and found that many of these cells also bore α4 (see online supplementary figure S1C), thus formally expressing both α4β7 and αEβ7, although coexpression of α4β1 and αEβ7 might also lead to this picture. Therefore, we directly labelled vedolizumab (vedo) and an etro-s antibody posessing the identical antigen recognition site as etrolizumab (etro) and used these antibodies in flow cytometry. While the majority of etro-s+ cells was also stained by vedo, we were able to detect a small subset of T lymphocytes that was targeted by etro-s but not vedo and this subset was significantly larger in CD8+ than in CD4+ cells (see online supplementary figure S1D).

Differential expression of αEβ7 and α4β7 on T cell subsets in the peripheral blood. (A) Left panels: representative flow cytometric analyses of αEβ7 (upper panels) and α4β7 expression (lower panels) on CD4+ and CD8+ blood lymphocytes from patients with UC, Crohn's disease (CD) and control donors along with isotype control stainings. Numbers indicate the percentage of αEβ7+ and α4β7+ cells. Right panels: pooled statistics of αEβ7 expression (upper panel) and α4β7 expression (lower panel) on peripheral blood CD4+ and CD8+ T cells from patients with UC (n=13), CD (n=24) and control donors (n=13). (B) Flow cytometry of αEβ7 and α4β7 expression on CD4+Foxp3+ (Treg), CD4+Foxp3− (Teff), CD4+IFN-γ+ (Th1), CD4+IL-4+ (Th2), CD4+IL-9+ (Th9) and CD4+IL-17A+ (Th17) cells from the peripheral blood of patients with UC (n=6–15), CD (n=30–44) and control donors (n=11–21). Peripheral blood mononuclear cells were stimulated overnight to induce cytokine production. IFN, interferon; IL, interleukin.
Supplementary figures
Next, we quantified the expression of αEβ7 on CD4+ T cell subsets (figure 1B). While Treg cells and overall Teff cells had low levels of αEβ7, the expression was markedly increased on Th17 and Th9 cells with the highest levels on Th9 cells. With regard to the expression of α4β7, Th2 and Th17 cells showed particularly high expression, while levels on Treg, Th1 and Th9 cells were lower.
In CD8+ T cell subsets,18 interleukin (IL)-9 secretion was correlated with a high expression of αEβ7, while Tc2 cells expressed particularly low levels of αEβ7 (see online supplementary figure S1E). Taken together, these findings supported the notion that αEβ7 and α4β7 are differentially expressed on various T cell subsets, suggesting that antiadhesive therapies may not equally block adhesion of all lymphocyte subsets in IBD.
High expression of αEβ7 on intestinal CD4+ and CD8+ T lymphocytes in IBD
In subsequent studies, we explored the expression of αEβ7 on intestinal lymphocytes in IBD by immunohistochemistry. We observed that both in CD and UC the percentage expression of αEβ7 on CD4+ T cells is higher than in controls and exceeds 10%, therefore being markedly higher than in the peripheral blood (figures 2A, D). However, the expression of αEβ7 on Foxp3+ cells in the lamina propria was unchanged in samples from CD and UC compared with controls (around 5%; figure 2B,D), suggesting that mucosal Treg cells might be a population with particularly low expression of αEβ7.

Differential expression of αEβ7 and α4β7 on IBD T cell subsets in the lamina propria. (A–C) Representative Immunofluorescent stainings of cryosections from patients with UC, Crohn's disease (CD) and control donors (n=15–21) for CD4 (A), Foxp3 (B), CD8 (C) and CD103 (αE integrin, A–C) along with control stainings (A). Scale bars: 25 µm (upper rows), 10 µm (lower rows). (D) Quantitative analysis of the frequency of αEβ7 expression on intestinal CD4+, Foxp3+ and total, lamina propria and intraepithelial CD8+ T cells as indicated.
The αEβ7 expression on CD8+ cells in the gut markedly differed between CD8+ cells in the lamina propria and the epithelium (figure 2C, D). However, the expression did not significantly vary between controls, UC and CD. Overall, αEβ7 was expressed by around 40% of the CD8+ lymphocytes, thus on a remarkably greater fraction than in the peripheral blood. Again, the differential expression of αEβ7 on CD4+ and CD8+ lymphocytes highlighted the idea that therapeutically blocking this integrin might have different effects on lymphocyte subpopulations.
To further explore the nature of these CD8+αEβ7+ cells in the intestine, we performed additional stainings with the T resident memory (TRM) cell marker CD69.19 We found that around 40% of the CD103+ cells also expressed CD69, roughly equalling the frequency of CD8 expression among CD103+ cells (see online supplementary figure S2A) and thus suggesting that the majority of CD8+αEβ7+ T lymphocytes are TRM cells. Moreover, we sought to exclude that DCs add to the CD8+αEβ7+ population as both CD8 and αEβ7 are potential markers of DC subsets.20 As expected, we identified αEβ7+ DCs using the pan-DC marker CD11c,21 but almost no CD11c+ cell coexpressed CD8 and coexpression of CD123 (plasmacytoid DCs) and CD141 (type II classical DCs) with αEβ7+ was very low (data not shown).
The high expression of αEβ7 on CD4+ and CD8+ lymphocytes in the intestinal mucosa compared with the peripheral blood was consistent with the idea that αEβ7 might be induced on both CD4+ and especially CD8+ lymphocytes upon reaching the intestinal mucosa. However, we also considered that this finding might be due to accumulation of these cells following preferential homing. To address this, we performed another series of stainings with vedo and etro-s (see online supplementary figure S2B). As expected, virtually all vedo+ cells were also etro-s+ but in contrast to the peripheral blood a relevant single etro-s+ population could be identified. As this was markedly different from the pattern in the peripheral blood (see online supplementary figure S1D), it strongly suggested that indeed regulation of integrin expression is responsible for the differential integrin levels in the peripheral blood and the intestine. Moreover, these experiments proved that targeting β7 instead of α4β7 increases the number of target cells not only in the blood but also in the gut.
TGF-β and butyric acid are key regulators of αEβ7 expression on CD4+ and CD8+ T lymphocytes
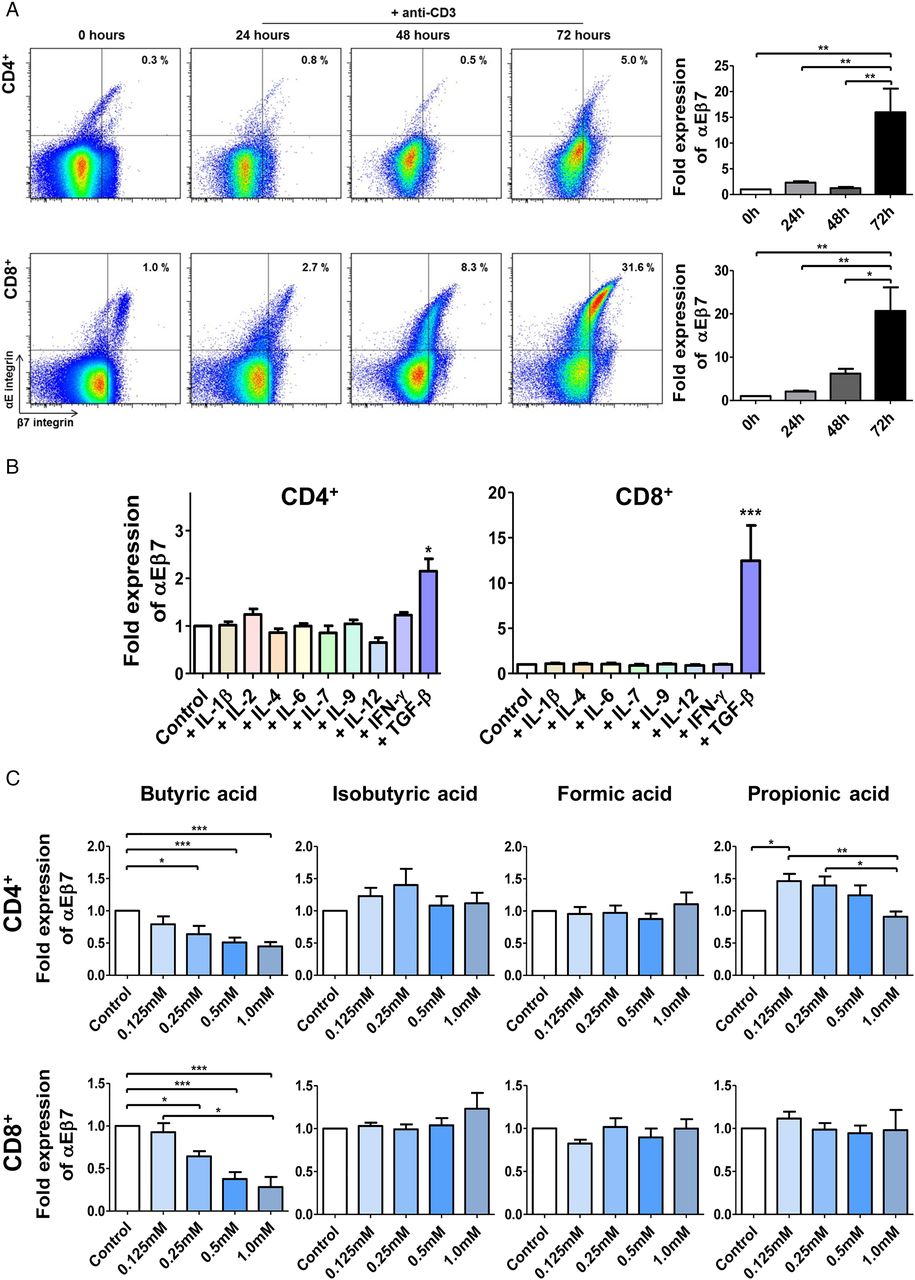
To further elucidate the mechanisms controlling αEβ7 upregulation in the intestinal mucosa, we studied αEβ7 expression in response to various stimuli in T cell cultures. Here, we noted that T cell receptor (TCR) stimulation caused a marked upregulation of αEβ7 on CD4+ and CD8+ T cells (figure 3A).

Expression of αEβ7 is differentially regulated in T cell subsets. (A) Left panels: representative flow cytometric analyses of αEβ7 expression on CD4+ and CD8+ T cells from the peripheral blood upon stimulation with anti-CD3 antibodies. Right panels: flow-cytometric quantification of αEβ7 expression on CD4+ and CD8+ cells upon stimulation with anti-CD3 antibodies. Values were normalised to the respective value before stimulation. (B) Flow cytometry of αEβ7 expression on CD4+ and CD8+ T cells upon incubation with the indicated cytokines for 72 hours together with anti-CD3/anti-CD28 antibodies (n=5–15). (C) Flow cytometry of αEβ7 expression on CD4+ and CD8+ T cells upon treatment with short chain fatty acids for 72 hours together with anti-CD3/anti-CD28 antibodies (n=8–37). IFN, interferon; IL, interleukin; TGFβ, transforming growth factor β.
In the following experiments, we assessed whether cytokines might affect αEβ7 expression. With the exception of TGF-β, however, none of the cytokines tested had any effect on αEβ7 levels. Interestingly, the potential of TGF-β to induce αEβ7 was clearly higher in CD8+ than in CD4+ T cells (figure 3B). When CD4+ lymphocyte subsets were analysed, we observed that TGF-β was not able to induce αEβ7 expression on Th9 cells (see online supplementary figure S3A), the subset with the highest intrinsic αEβ7 levels. This might be explained by the role of TGF-β in Th9 differentiation22 in the way that high levels of αEβ7 on Th9 cells are a result of prior contact with TGF-β and can therefore not be further boosted.
As earlier murine studies reported that TGF-β simultaneously downregulates α4β723 and the effect of α4β7-inducing retinoic acid on αEβ7 expression has not been tested so far, we further dissected the impact of these stimuli on the expression of both markers on human T cells (see online supplementary figure S3B–E). We could demonstrate that TGF-β indeed reduces α4β7 expression on CD8+ but not CD4+ cells, while retinoic acid had no significant effect on αEβ7 levels.
Finally, we incubated CD4+ and CD8+ T cells with short-chain fatty acids derived from microbes in the human gut24 (figure 3C). While isobutyric acid and formic acid had no effects, low concentrations of propionic acid significantly increased αEβ7 expression on CD4+ but not CD8+ T cells. However, we detected a significant and dose-dependent downregulation of αEβ7 expression on both CD4+ and CD8+ T cells upon treatment with butyric acid, a key inducer of anti-inflammatory Treg cells.
Collectively, these results showed that stimulation of T cells via the TCR, butyric acid and TGF-β are key regulators of αEβ7 expression. Moreover, TGF-β and TCR stimulation trigger a much stronger induction of αEβ7 expression on CD8+ than on CD4+ T lymphocytes. This is further supporting the notion that αEβ7 is especially important for the adhesion of CD8+ cells.
Etro-s antibody blocks adhesion of IBD T cells to E-cadherin and MAdCAM-1 more effectively than vedo
To investigate the interaction of α4β7 and αEβ7 with their ligands in vitro, we performed a series of adhesion assays with plates coated with E-Cadherin and/or MAdCAM-1. First, we employed etro-s, which blocks β7 integrin, in two concentrations corresponding to the reported effective etrolizumab levels in human serum.25 This antibody blocked the adhesion of both CD4+ and CD8+ T cells from patients with IBD and control donors to E-cadherin (figure 4A and online supplementary figure S4E). Moreover, we tested an anti-αE antibody and observed that this antibody was also able to block the adhesion of CD4+ and CD8+ cells to E-cadherin (see online supplementary figure S4A).
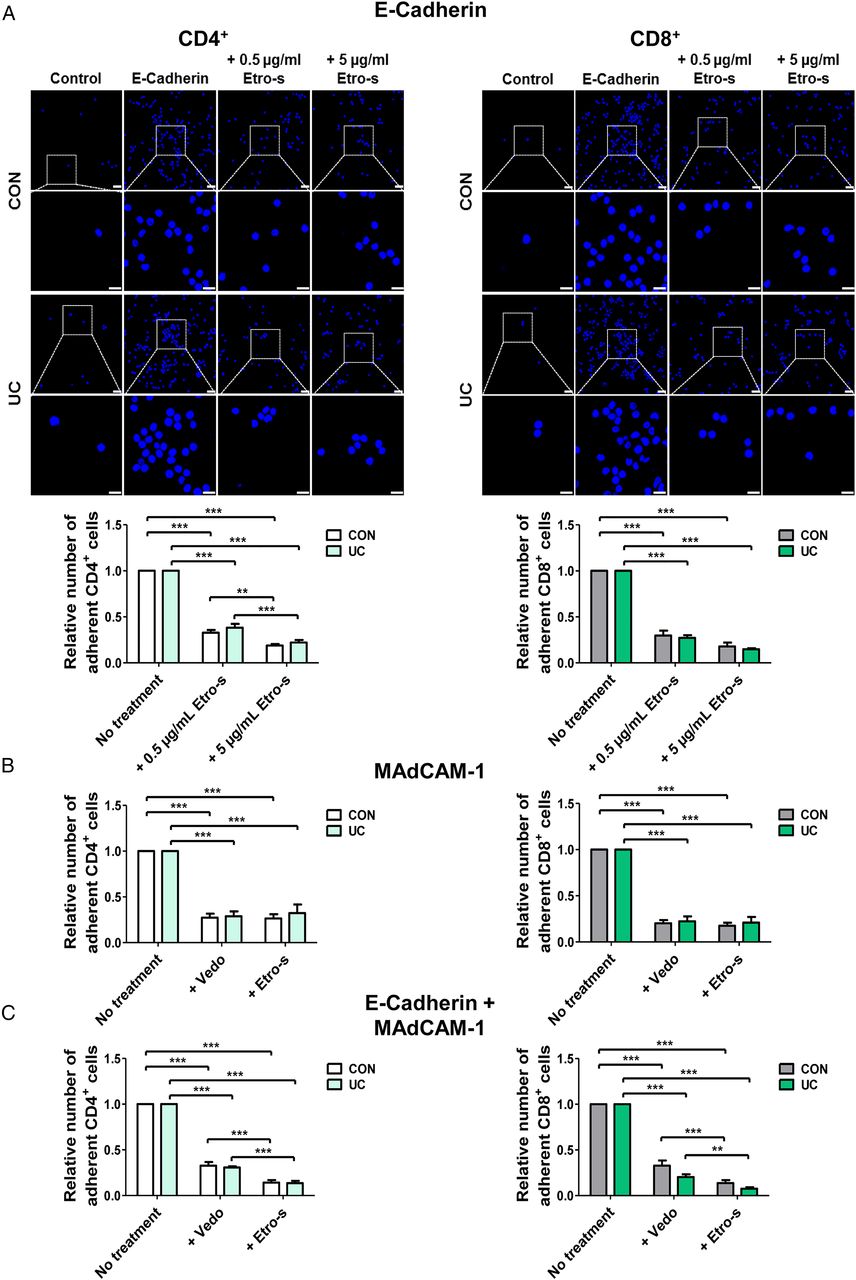
Adhesion of CD4+ and CD8+ IBD T cells to mucosal vascular addressin cell adhesion molecule 1 (MAdCAM-1) and E-cadherin is blocked by etrolizumab surrogate (etro-s) antibody. (A) Upper panels: representative adhesion assays showing the adhesion of UC and control CD4+ and CD8+ T cells treated with or without etro-s to slides coated with E-cadherin and control conditions without ligand. Scale bars: 25 and 10 µm (inserts). Lower panels: quantitative analysis of the adhesion of CD4+ and CD8+ T cells from patients with UC and control donors to E-cadherin upon treatment with different concentrations of etro-s. Quantitative analysis of the adhesion of CD4+ and CD8+ T cells from patients with UC and control donors to MAdCAM-1 (B) and E-cadherin+MAdCAM-1 (C) upon treatment with vedolizumab (vedo) or etro-s.
Next, we compared how vedo and etro-s antibodies block the adhesion of lymphocytes to MAdCAM-1. Adhesion of CD4+ as well as CD8+ T cells from patients with IBD and control donors was similarly blocked by both antibodies (see figure 4B and online supplementary figure S4E). Additionally, more untreated CD8+ T cells bound to E-cadherin and MAdCAM-1 compared with CD4+ cells matching which preferential αEβ7 and α4β7 expression on CD8+ T cells (see online supplementary figure S4A, B). Subsequently, we coated slides with E-cadherin together with MAdCAM-1. The number of adherent CD4+ and CD8+ cells was significantly reduced after treatment with etro-s compared with vedo (see figure 4C and online supplementary figure S4E). In preparation of our in vivo experiments, we then coated slides with human and murine E-cadherin. Both CD4+ and CD8+ cells from UC and controls bound to murine E-cadherin in lower numbers compared with human E-cadherin. However, binding still occurred to a considerable extent of around 70% (see online supplementary figure S4D) showing a marked interaction between human receptor and murine ligand. In conclusion, our data show that etro-s blocks the adhesion of human lymphocytes to both MAdCAM-1 and E-cadherin and suggest that cell adhesion is reduced upon etro-s treatment compared with vedo when both ligands are present.
Superior reduction of colonic IBD CD8+ and Th9 cell accumulation in vivo upon treatment with etro-s compared with vedo by additional inhibition of retention
We next explored the functional relevance of the above-mentioned interactions in vivo by using a humanised mouse model for analysis of T cell accumulation in the inflamed gut.13 We thus induced dextran sodium sulfate (DSS) colitis in immunodeficient mice prior to adoptive transfer of human T cells to the ileocolic artery (figure 5A). Intravital confocal microscopy showed that carboxyfluorescein succinimidyl ester (CFSE)-labelled CD4+ and CD8+ UC and control cells could be detected in the murine lamina propria and near the epithelium (see online supplementary figure S5A). Moreover, z-stacks demonstrated the positioning of human T cells in close proximity to epithelial cells and confirmed extravasation (see figure 5B and online supplementary figure S5B).

Etrolizumab surrogate (etro-s) antibody is more potent than vedolizumab (vedo) in blocking the in vivo accumulation of UC CD8+ and Th9 cells. (A) Illustration of adoptive transfer. After anaesthesia, laparotomy and exposure of the ileocecal region, the ileocolic artery was punctured (left image) and carboxyfluorescein succinimidyl ester (CFSE)-labelled human T cells were injected. Here, ink is injected to demonstrate successful puncture (right image). (B) 3D reconstruction from a representative z-stack (cf. online supplementary figure S5B) obtained during intravital confocal microscopy highlighting an extravasated human cell (green in white circle). Blue: murine cells. Red: murine vessels. (C, E) Upper panels: representative confocal in vivo microscopy images of murine colon after adoptive transfer of UC CD4+ (C) and CD8+ T cells (E) to the ileocolic artery. Treatment with blocking antibodies is indicated. Scale bars: 25 µm (upper row) and 10 µm (lower row). Left lower panels: representative flow cytometry of murine lamina propria mononuclear cells 3 hours after adoptive transfer of UC CD4+ (C) and CD8+ T cells (E). Numbers denote the CFSE+ cell fraction. Right lower panels: flow-cytometric quantification of CFSE+ cells in the murine lamina propria 3 hours after adoptive transfer of UC CD4+ (C) or CD8+ T cells (E) in the presence or absence of integrin blockers, as indicated. Data were pooled from five independent experiments. (D) Left panel: flow-cytometric quantification of CFSE+ cells in the murine lamina propria 3 hours after adoptive transfer of UC CD8+ cells upon treatment with vedo and anti-CD103 as indicated (n=7). Right panel: flow-cytometric comparison of the colonic accumulation of untreated UC CD4+ and CD8+ T cells. (F) Flow-cytometric comparison of the relative portion of Th1 and Th9 cells among CFSE+ purified CD4+ T cells before and after transfer (n=5–6). (G) Flow-cytometric comparison of the relative portion of Th1 and Th9 cells among CFSE+ purified CD4+ T cells in the murine lamina propria upon vedo or etro-s treatment (n=4–5). AT, after transfer; BT, before transfer; SSC, sideward scatter.

Continued.

Continued.
Further studies verified that E-cadherin is present throughout the colon of DSS-treated mice in a pattern comparable to patients with IBD (see online supplementary figure S5C). We continued our studies by treating CD4+ T cells from UC patients with vedo, anti-CD103 or both before adoptive transfer to DSS-treated mice. As determined by in vivo confocal microscopy and flow cytometry of lamina propria mononuclear cells (LPMCs) isolated 3 hours after transfer, we found that anti-CD103 treatment alone was not able to reduce colonic accumulation of UC CD4+ cells and the combination of anti-CD103 with vedo and vedo alone yielded an equal reduction of colonic UC CD4+ cells (see online supplementary figure S6A). Yet, in a similar series of experiments with CD8+ cells, combined blockade of α4β7 and αEβ7 through vedo and anti-CD103 was followed by a decrease of colonic UC CD8+ cells compared with sole α4β7 inhibition by vedo (see figure 5D and online supplementary figure S6B). Consistently, anti-CD103 treatment also had significant effects. These results suggested that, while additional blockade of αEβ7 together with α4β7 might not affect the colonic localisation of peripheral UC CD4+ cells due to their low αEβ7 expression, higher levels of αEβ7 on UC CD8+ cells might go along with a further reduction of colonic accumulation after combined blockade of both adhesion molecules.
Subsequently, we found that both vedo and etro-s similarly decreased the number of colonic UC CD4+ T cells (figure 5C). CD8+ T cells, however, were found in lower numbers upon etro-s treatment compared with vedo treatment, supporting the notion that combined α4β7 and αEβ7 blockade might be of special relevance for CD8+ T cells (figure 5E).
As αEβ7 expression is not uniformly distributed among different CD4+ T subsets, we reasoned that etro-s might still additionally affect certain CD4+ T subsets. Hence, we stained LPMCs for αEβ7 and markers of Th1, Th9 and Th17 cells, respectively (see figure 5F, G and online supplementary figure S6C, D). Analysis of these markers on CFSE+ cells demonstrated that αEβ7+ cells as well as Th1, Th9 and Th17 cells were enriched among untreated human cells that had entered the lamina propria compared with aliquots from the respective donor before transfer. When comparing vedo and etro-s treated cells in the lamina propria, we found that the proportion of αEβ7+ cells was reduced upon etro-s treatment matching with its additional blockade of αEβ7. Concerning the CD4+ subsets, numbers of Th9 but not Th1 and Th17 cells were reduced with etro-s compared with vedo. Although limited by the low number of cells analysed, these findings prompted the conclusion that while the overall CD4+ T cell infiltrate was not reduced after etro-s compared with vedo treatment, etro-s reduced the accumulation of Th9 cells.
To analyse the effect that is accountable for these findings in more detail, we compared colonic accumulation of UC CD8+ cells 0.5 and 3 hours after adoptive transfer (figure 6). Interestingly, vedo and etro-s had comparable effects after 0.5 hours. After 3 hours, however, we observed similar differences as above. This is in line with the view that both compounds similarly inhibit α4β7-mediated homing, while—subsequently—etro-s possesses an additional and longer-lasting mechanism, which is easily reconcilable with the concept of inhibition of retention.

Time course of vedolizumab (vedo) and etrolizumab surrogate (etro-s) effects—evidence for in vivo blockade of retention. Upper panels: representative intravital confocal images of murine colon at the indicated time points after adoptive transfer of UC CD8+ T cells treated with vedo and etro-s as indicated. Scale bars: 50 µm. Lower panels: flow-cytometric quantification of carboxyfluorescein succinimidyl ester (CFSE+) cells in the murine lamina propria 0.5 hours (left) and 3 hours (right) after adoptive transfer of UC CD8+ T cells in the presence or absence of integrin blockers, as indicated (n=3).
Taken together, these results suggested that combined α4β7 and αEβ7 blockade in UC T cells through etro therapy results in a superior reduction of distinct lymphocyte subsets in the colon compared with sole α4β7 blockade through vedo.
To get an estimate of the potential clinical relevance of this finding, we studied the expression of αEβ7 on peripheral blood lymphocytes from patients with IBD treated with vedo (figure 7A–C). Interestingly, we observed that no changes in the expression of αEβ7 on CD4+ Teff and Treg cells occurred when comparing samples obtained during the induction phase with samples obtained during the maintenance phase of treatment. However, we noticed a significant increase in the expression of αEβ7 but not α4β7 (figure 7D) on CD8+ T cells in the maintenance compared with the induction phase suggesting that these T cells might possibly develop αEβ7-dependent compensatory strategies to ensure their colonic positioning despite suppression of gut homing via vedo-induced blockade of α4β7.
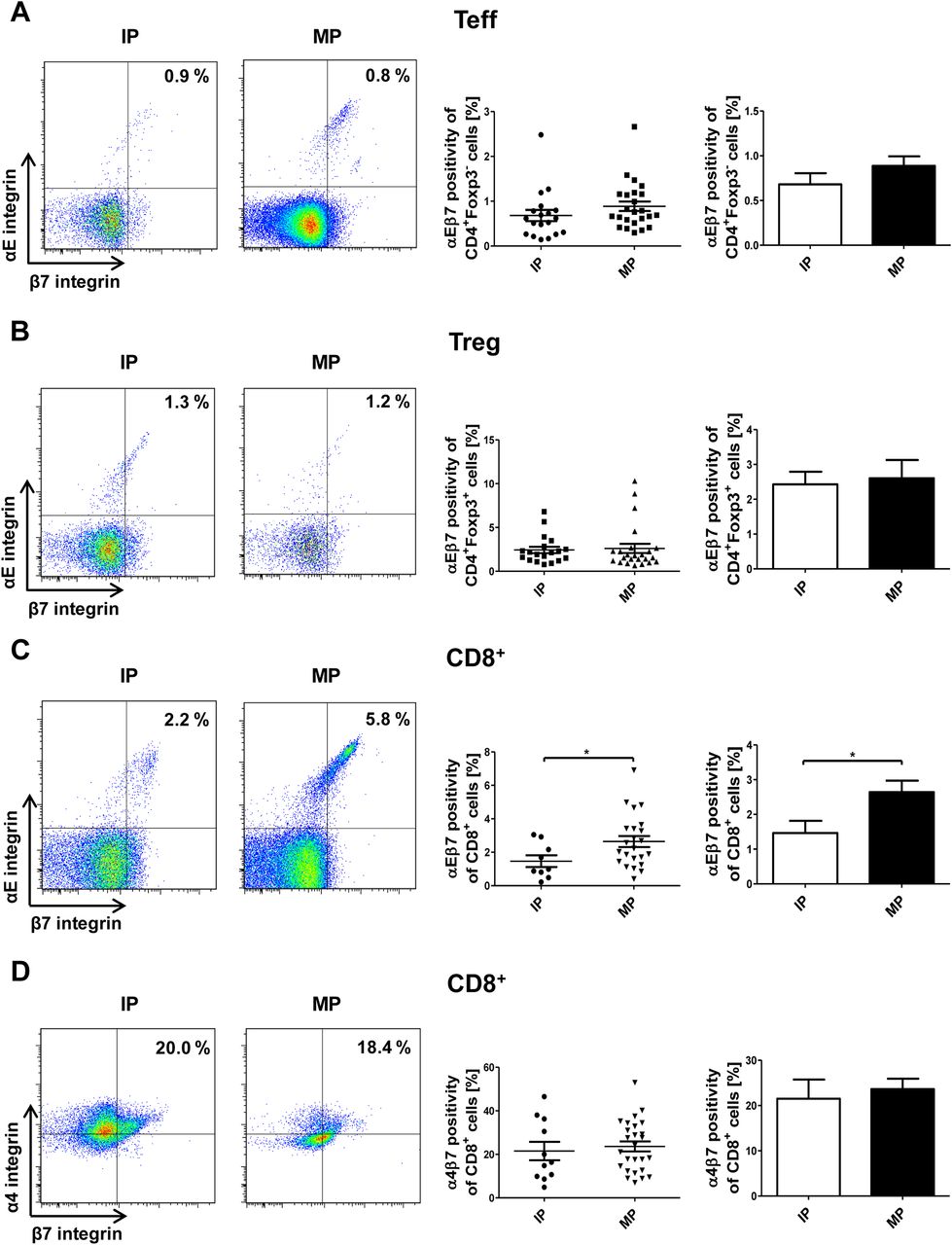
Vedolizumab treatment leads to higher αEβ7 expression on blood IBD CD8+ cells. (A–C) Representative flow cytometric data and quantitative analyses of αEβ7 expression on peripheral CD4+Foxp3− (A), CD4+Foxp3+ (B) and CD8+ (C) T cells during the induction phase (IP, i.e. treatment 1–3) and maintenance phase (MP, i.e. treatment 4 and later) of vedolizumab therapy as indicated. (D) Representative flow cytometric data and quantitative analyses of α4β7 on peripheral CD8+ T cells during the IP and MP of vedolizumab therapy.
Discussion
Although clinical treatment with the monoclonal anti-α4β7 antibody vedo is available for therapy of patients with IBD for some years now, the knowledge about the factors that control the colonic homing and retention of lymphocytes is still fragmentary.26 In particular, this is the case for CD8+ T cells, a lymphocyte subset that has been largely overlooked in the past due to the prominent role of CD4+ T cells.4 ,27 However, CD8+ T cells are thought to be not only important effector cells but also crucial mediators of pathological immune responses in a number of immunologically mediated diseases.18 ,28 Moreover, there is evidence that CD8+ T cells are also implicated in the events leading to human and murine colitis.5 ,6 Using an in vivo model for IBD T cell trafficking in humanised mice, we provide evidence here that blockade of β7 is more effective in suppressing colonic accumulation of CD8+ T cells than α4β7 blockade, suggesting a crucial regulatory role of αEβ7. Similar findings were obtained for Th9 cells, a recently identified CD4+ T cell subset with important pathogenic function in UC,29 but not for total CD4+ T cells. These results provide new insights into the molecular factors that control CD4+ and CD8+ T cell homing and retention in patients with IBD and suggest new avenues for therapy.
Interestingly, little attention has been paid to αEβ7 in the framework of IBD since its discovery16 and functional in vivo data for human cells have not been reported so far. We uncovered that both αEβ7 and α4β7 are differentially expressed on blood and intestinal CD4+ and CD8+ T cells in patients with IBD with higher expression on CD8+ T cells. The potential of TGF-β and TCR stimulation to induce αEβ7 expression was much higher in CD8+ than in CD4+ T cells, possibly leading to this finding. This observation led to the hypothesis that differential expression of adhesion markers might also concern CD4+ T lymphocyte subsets in IBD. Indeed, according studies revealed that Th9 cells bear particularly high levels of αEβ7, while Th2 and Th17 cells had the highest expression of α4β7. Similarly, Tc9 cells expressed the highest levels of αEβ7 among CD8+ T cells. Collectively, these results were consistent with the idea that clinical therapy with α4β7 or β7 blockers differentially affects distinct T lymphocyte subsets and might therefore have specific strengths and weaknesses in targeting the adhesion of these populations. This could be of potential therapeutic relevance as there is fundamental evidence that different T cell subsets and their signature cytokines are of varying importance in individual patients.4 ,6 ,29–32 Compatibly, it is postulated that therapy will prospectively be assigned on the basis of biomarkers reflecting such immunological profiles.33
Differential expression of αEβ7 on CD4+ and CD8+ cells was demonstrated in the peripheral blood and the intestine of controls and patients with IBD. Moreover, we show that both in the blood and the gut more cells can be targeted with anti-β7 than with anti-α4β7 antibodies. Matching with earlier reports,34 the expression of αEβ7 was found to be higher in the intestine than in the peripheral blood and we provide further evidence35 that this is majorly due to regulation of expression and not frequency shifts following homing. Stimulation experiments demonstrated that mainly TGF-β and TCR stimulation may be responsible for this increase of αEβ7 on intestinal lymphocytes. In conclusion, these stimuli might induce an integrin profile that allows targeting of these cells with etro but not vedo. Moreover, butyric acid, a key inducer of anti-inflammatory Treg cells in the colon,24 markedly suppressed αEβ7 expression on both CD4+ and CD8+ T cells. Thus, it seems possible that commensal bacteria in close proximity to intestinal epithelial cells might affect αEβ7 levels via butyric acid production.
Further stainings indicated that the majority of CD8+αEβ7+ cells belong to the compartment of CD69+ TRM cells. As several authors have highlighted,19 ,36–38 a subset of mainly CD8+ lymphocytes develops into resident cells that may immediately react to translocated antigens with cytokine secretion and thus coordinate prompt defence against infectious agents but might also contribute to the dysregulated immune response in IBD.39 In conclusion, it is likely that TRM cells are a main target of anti-β7 antibodies in the intestine.
To investigate the functional relevance of αEβ7 and α4β7 for colonic positioning in vivo, we used a recently described humanised mouse model13 where DSS colitis is induced in immunodeficient mice prior to adoptive transfer of labelled human lymphocytes to the ileocolic artery. Preliminary experiments showed that human αEβ7 mediates adhesion to murine E-cadherin in spite of reduced adhesion compared with human E-cadherin. Moreover, confocal in vivo imaging after adoptive transfer of human cells demonstrated that UC T cells can be found in close proximity of the murine epithelium, thus allowing an interaction of human αEβ7 with murine E-cadherin in vivo.
In humanised mice, we showed that the number of UC CD4+αEβ7+ and Th9 cells is reduced upon β7 blockade with the etro-s antibody FIB504, which shares the identical antigen recognition site with etrolizumab, in comparison with the α4β7 blocker vedo. Thus, β7 blockade via the former antibody targeted CD4+ subsets with high αEβ7 expression in UC, although the overall CD4+ T cell population was not affected. This is particularly interesting as we found that the expression of α4β7 on Th9 cells is relatively low, suggesting that a considerable portion of these cells might be able to access the intestine despite α4β7 blockade with vedo. Additionally targeting the epithelial retention of Th9 cells by etrolizumab might therefore narrow this ‘gap’ and may be clinically relevant, as an expansion of these cells has been demonstrated in patients with IBD and as IL-9 blockade was effective in a murine model of colitis induced by the hapten reagent oxazolone.29
Studies using cell transfer of human T cells to DSS-treated mice showed that α4β7 blockade via vedo not only reduces the colonic homing of UC CD4+ but also CD8+ T cells. In addition, β7 blockade with etro-s or treatment with vedo, together with an anti-CD103 antibody, further reduced the number of CD8+ T cells but not CD4+ T cells. Given our data on the expression and regulation of αEβ7 on peripheral CD4+ and CD8+ cells, this may reflect the higher expression of αEβ7 on CD8+ T cells. Consistently, anti-CD103 antibody treatment had significant effects on T cell accumulation in UC CD8+ T cells rather than CD4+ cells, highlighting the functional relevance of αEβ7 for the former cells.
This is in line with data from two murine studies: While Annacker et al40 reported that αE plays no role for the pathogenic and beneficial potential of effector and regulatory CD4+ lymphocytes, respectively, but rather has an important regulatory role in DCs in the T cell transfer model of colitis, Lúdvíksson et al41 have shown that pan-lymphocyte inhibition of αE significantly ameliorated disease in a T cell-dependent colitis model.
Different suggestions have been made regarding the concrete function of αEβ7 for cell trafficking. While it is known that αEβ7 interacts with the epithelial ligand E-cadherin and many authors have thus proposed that αEβ7 is responsible for retention of homed lymphocytes within or near the epithelium,16 ,42 αEβ7 has also been shown to bind to intestinal endothelial cell lines independently of E-cadherin.43 Moreover, the number of lamina propria lymphocytes is reduced in αE-deficient mice,44 leading to the possibility of additional direct functions for αEβ7 in gut homing through an alternative, yet unknown, ligand.45 However, our findings do not provide evidence for alternative homing via αEβ7. On the contrary, although limited by the restricted observation period achievable in our model, our data provide for the first time functional in vivo evidence that αEβ7 is indeed implicated in the retention of human UC T cells.
Interestingly, clinical data from patients with UC treated with vedo showed that more αEβ7, but not α4β7, is found on blood CD8+ T cells after the induction phase of vedo therapy. This suggests that lymphocytes might use alternative molecules such as αEβ7 to ensure their localisation within the intestine despite α4β7 inhibition by vedo, further supporting the concept that blocking αEβ7 together with α4β7 may increase the effects of antiadhesion therapy.
Taken together, our data suggest a key regulatory role of αEβ7 for CD8+ rather than global CD4+ T cell trafficking. Nevertheless, αEβ7 seems to be important in specified CD4+ subsets, namely Th9 cells (figure 8). Additional therapeutic targeting of αEβ7 on CD8+ and Th9 cells with antibodies such as etrolizumab might therefore open new avenues for clinical treatment of IBD by increasing and extending the impact of sole α4β7 inhibition.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
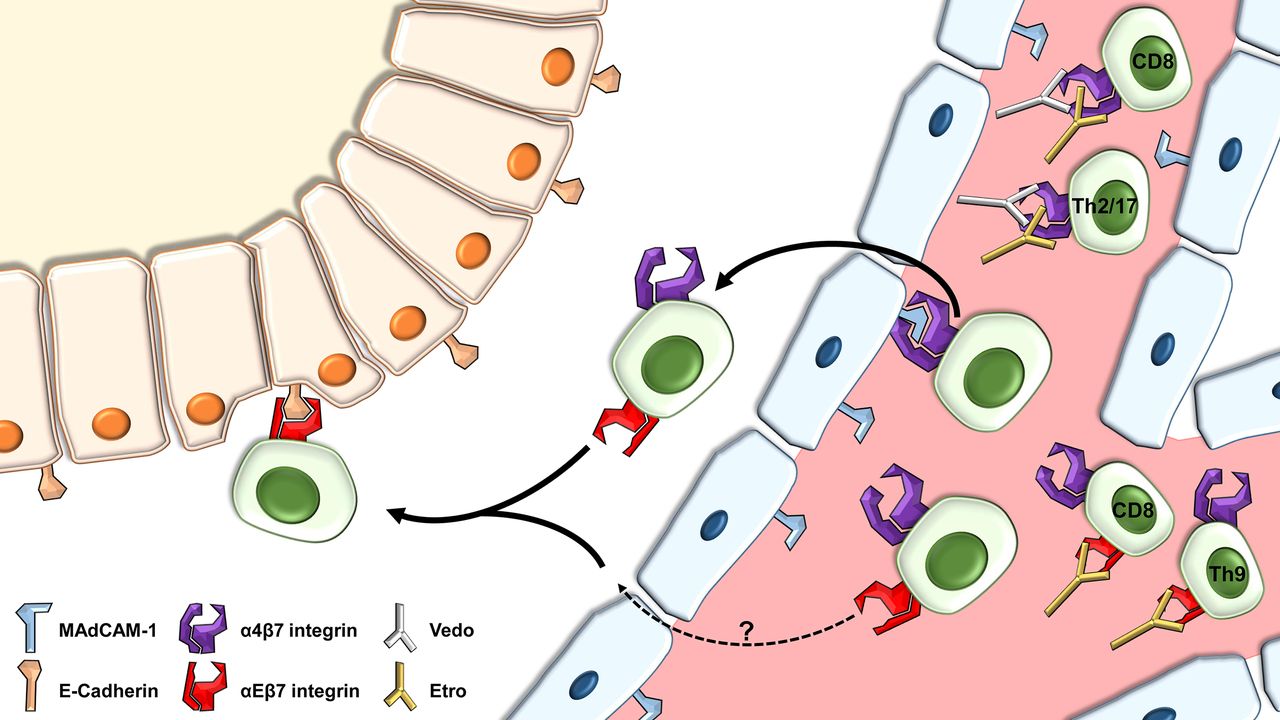
Model for the control of colonic localisation of IBD T lymphocyte subsets by α4β7 and αEβ7. Cells may extravasate to the lamina propria via interaction of α4β7 with mucosal vascular addressin cell adhesion molecule 1 (MAdCAM-1). Possibly, an alternative αEβ7-dependent pathway via an unknown ligand might exist. After homing to the intestine, interaction of αEβ7 with epithelial E-cadherin allows retention of T lymphocytes in or near the epithelium. Vedolizumab (vedo) blocks the interaction of α4β7 with MAdCAM-1, which particularly affects CD8+, Th2 and Th17 cells, while inhibition of αEβ7 by etrolizumab (etro) is of special relevance for CD8+ and Th9 cells.
Acknowledgments
The authors thank Dr Simon Völkl, Florentine Koppitz (both Core Facility Immune Monitoring of the University Erlangen-Nuremberg), Karin Enderle, Tatyana Kisseleva and Michael Dobrönti for excellent scientific technical support. The research of SZ, RA, RLP, CN, IA and MFN was supported by the Interdisciplinary Center for Clinical Research (IZKF), the Clinical Research Group CEDER of the German Research Council (DFG), the DFG topic programme on Microbiota, the Emerging Field Initiative, the ELAN programme of the University Erlangen-Nuremberg and the DFG Collaborative Research Centers 643, 796 and 1181. AW is supported by BB/J004529/1: The Gut Health and Food Safety ISP.
References
Footnotes
Contributors SZ, DS and AF performed experiments. SZ, RA, RLP, AW, CN, IA and MFN provided clinical samples, protocols, reagents or designed experiments. SZ, CN, IA and MFN analysed and interpreted the data. SZ and MFN drafted the manuscript. All authors critically revised the manuscript for important intellectual content.
Funding This work was supported by the Clinical Research Group 257 CEDER of the Deutsche Forschungsgemeinschaft and the Interdisciplinary Center for Clinical Research of the Friedrich Alexander Universität Erlangen-Nürnberg.
Competing interests The etrolizumab surrogate antibody FIB504 was provided by Genentech, San Francisco, California, USA. The company was neither involved in the conception and conduction of the study nor in the analysis and interpretation of the results. MFN has served as an advisor for Abbvie, MSD, Boehringer, Takeda, Janssen, Pentax and Giuliani. A research project of MFN and SZ is supported by Takeda.
Ethics approval Ethics Committee of the University Erlangen-Nuremberg.
Provenance and peer review Not commissioned; externally peer reviewed.