Article Text

Abstract
BACKGROUND Intestinal metaplasia is considered a risk factor for the development of gastric adenocarcinomas of the intestinal type and is found in approximately 20% of gastric biopsies. Conventional histology only detects advanced stages of intestinal metaplasia.
AIMS To study expression of the enterocyte specific adhesion molecule liver-intestinal (LI)-cadherin in intestinal metaplasia as well as in gastric cancer, and to evaluate its use as a diagnostic marker molecule.
PATIENTS Gastric biopsies (n=77) from 30 consecutive patients (n=30; aged 28–90 years) as well as surgically resected tissue samples (n=24) of all types of gastric carcinomas were analysed.
METHODS Single and double label immunofluorescence detection on cryosections of gastric biopsies; alkaline phosphatase antialkaline phosphatase method on paraffin embedded carcinoma tissue sections.
RESULTS Of 77 biopsies (from 30 patients), 12 (from 10 patients) stained positive for LI-cadherin. LI-cadherin staining correlated with the presence of intestinal metaplasia. Conventional histological diagnosis however failed to detect subtle gastric intestinal metaplasia (three of 10 patients). In contrast, only LI-cadherin and villin were positive in these cases whereas sucrase-isomaltase also failed to detect intestinal metaplasia in four of 10 patients. Well differentiated gastric carcinomas showed intense staining for LI-cadherin while undifferentiated carcinomas showed only weak diffuse cytoplasmic staining.
CONCLUSIONS To detect early metaplastic changes in the gastric mucosa, LI-cadherin has a sensitivity superior to sucrase-isomaltase and conventional histology and comparable with that of villin. Its specificity exceeds that of villin. Thus LI-cadherin represents a new, reliable, and powerful marker molecule for early detection of gastric intestinal metaplasia and well differentiated adenocarcinomas.
- stomach
- intestinal metaplasia
- cadherins
- carcinogenesis
Abbreviations used in this paper
- BSA
- bovine serum albumin
- IgG
- immunoglobulin G
- LI-cadherin
- liver-intestinal cadherin
- PBS
- phosphate buffered saline
- TBS
- Tris buffered saline
Statistics from Altmetric.com
Gastric carcinomas rank among the most common malignant diseases worldwide. While the incidence of gastric carcinomas has remained unchanged in China, Japan, Eastern Europe, and South America, its incidence has decreased in Western Europe and the USA, probably due to the influence of nutritional and environmental factors.1 The majority of gastric carcinomas (97%) belong to one of four subgroups of adenocarcinomas (papillary, tubular, mucinous, and signet ring cell-type). The remaining 3% of gastric carcinomas are adenosquamous, squamous, or undifferentiated (anaplastic) carcinomas. As most gastric carcinomas (80%) are diagnosed at an advanced stage leading to a five year survival rate of only 17%,1 it is important to identify early indicators and mucosal lesions associated with the development of these carcinomas.
Metaplastic epithelium in the stomach is considered to be a risk factor for the development of gastric adenocarcinomas of the intestinal-type. Morson described the possible transition from intestinal metaplasia to stomach cancer as far back as 1955.2 ,3 Since then, numerous studies have demonstrated that both metaplastic epithelium and gastric carcinoma exhibit similar expression patterns for various proteins and mucins commonly not expressed in the stomach.4-7 It is believed that normal gastric epithelium develops successively to adenocarcinoma via chronic atrophic gastritis, intestinal metaplasia, and dysplasia.8
Intestinal metaplasia of the stomach represents an alteration of the gastric mucosa into an epithelium with morphological and biological characteristics of the intestine.9 Intestinal metaplasia is found in approximately 20% of gastric biopsies examined10 and is more frequent in older patients.11
While the expression patterns of various cytoskeletal proteins, enzymes, and mucins in intestinal metaplasia are well documented, the role of the important class of cadherin cell adhesion molecules in the process of metaplastic transition has not been studied to date. Cadherins play an essential role in the structural and functional organisation of cells in tissues and organs of multicellular organisms. They represent a large family of transmembrane glycoproteins responsible for Ca2+ dependent intercellular adhesion and interaction. Cadherins are essential for animal morphogenesis,12-14 formation of intercellular junctions,15 cell polarity, and signal transduction.12 ,16
E-cadherin, the best known member of the classical cadherins, represents the most intensively examined cadherin with respect to cancer. The reduction in functional E-cadherin expression is thought to be a key event for the dysfunction of the cell-cell adhesion system, triggering invasive cell behaviour and the formation of metastases. Therefore, normal E-cadherin expression is considered to act as an “tumour invasion suppressor system”.17-19 With regard to analysing tumour inducing mechanisms, cadherins are in the forefront of medical research. Thus the correlation of cancer associated morphological changes with modulation of cadherin expression patterns is very important. As described for many other epithelial tumours, reduction in the cell adhesion molecule E-cadherin has also been associated with adenocarcinomas of the stomach.20-24Apart from the well documented tumour associated somatic alterations of cadherin function, Guilford and colleagues25 recently reported E-cadherin germline mutations predisposing to familial gastric cancer.
LI-cadherin was discovered as a structurally different cadherin specifically expressed in polarised epithelia of the liver and intestine of the rat.26 In contrast with classical cadherins, LI-cadherin is composed of seven instead of five extracellular cadherin repeats. In addition, the highly conserved cytoplasmic region of the classical cadherins containing 150–160 amino acids is replaced in LI-cadherin by a much shorter domain consisting of only 21 amino acids. This domain does not interact with catenins or actin cytoskeleton and is not essential for the adhesive function of LI-cadherin. Nevertheless, LI-cadherin mediates homotypic Ca2+ dependent cell-cell adhesion when expressed in L cells.27 Human LI-cadherin is exclusively found in the intestinal epithelium but not in the liver (Zitt et al, submitted). The stomach is also negative for LI-cadherin. Within the human intestinal mucosa, LI-cadherin is concentrated in the lateral domain of the plasma membrane residing between adjacent enterocytes but spared from adherens junctions whereas E-cadherin is concentrated in these very structures.
Since intestine specific markers such as villin, sucrase-isomaltase, and aminopeptidase N are known to be expressed in intestinal metaplastic cells and gastric adenocarcinomas28 while missing in the normal gastric mucosa, we studied expression of the intestine specific cell adhesion molecule LI-cadherin in the gastric epithelium undergoing neoplastic transformation.
Materials and methods
GASTRIC BIOPSIES AND HISTOPATHOLOGY
After obtaining informed consent, fresh gastric biopsies were taken during routine gastroscopy from randomly selected patients with upper gastrointestinal symptoms. Several endoscopists in the department were involved in the study. Biopsy specimens were generally obtained from the antrum, corpus, and fundus. The number of samples taken from each patient varied from two to six. All endoscopically removed biopsies were taken from gastric mucosa which appeared macroscopically normal. For histopathology, samples were fixed in formalin and paraffin embedded. For immunofluorescence microscopy, corresponding samples were first fixed in 9% sucrose at 4°C for one hour, then in 15% sucrose at 4°C for four hours, and were finally snap frozen in isopentane cooled to −196°C in liquid nitrogen. Several sections were prepared from both paraffin embedded and shock frozen biopsies. For histopathological evaluation, sections were stained with haematoxylin-eosin. Intestinal metaplasia was diagnosed based on the presence of goblet cells and occasional enterocytes within the foveolar epithelium.29
ANTIBODIES
LI-cadherin was detected using a polyclonal antiserum raised in rabbits against overexpressed and purified extracellular fragments of human LI-cadherin (Zitt et al, submitted). Polyclonal antibody to β-catenin was purchased from Sigma (Deisenhofen, Germany). Monoclonal antibody to E-cadherin was from Transduction Laboratories (Hamburg, Germany) and monoclonal antivillin ID2C3 was obtained from Coulter-Immunodiagnostics (Krefeld, Germany). The monoclonal antibody to sucrase-isomaltase was a kind gift from Dr H-P Hauri (Biozentrum Basel, Switzerland). Appropriate antibody dilutions were determined by serial dilutions in the presence of positive and negative controls. Final dilutions for immunostaining were as follows: anti-LI-cadherin, 1:500; anti-β-catenin, 1:1000; anti-E-cadherin, 1:400; antivillin, 1:50; and antisucrase-isomaltase, 1:1000. As secondary antibodies, affinity purified Texas Red conjugated goat antirabbit IgG (Dianova, Hamburg, Germany) and Cy2 conjugated goat antimouse IgG (Dako, Hamburg, Germany) were used at a 1:200 dilution.
For western blot analysis, anti-LI-cadherin antiserum was diluted 1:200 and horseradish peroxidase conjugated swine antirabbit antibodies (Dako, Hamburg, Germany) were used as secondary antibodies at a 1:2000 dilution.
GEL ELECTROPHORESIS
Shock frozen biopsies were homogenised in the presence of protease inhibitors and separated on a standard 8% sodium dodecyl sulphate-polyacrylamide gel.
IMMUNOBLOTTING
Proteins separated on the gel were transferred electrophoretically onto a Hybond C nitrocellulose membrane (Amersham-Pharmacia, Freiburg, Germany). The membrane was incubated for 30 minutes with Tris buffered saline (TBS) containing 5% skim milk and 0.1% Tween 20. The membrane was washed four times with 0.1% Tween 20 in TBS at intervals of 10 minutes and incubated for one hour with the primary antibody (diluted in TBS containing 5% skim milk and 0.1% Tween 20). The membrane was washed again as described above and incubated with the secondary antibody under identical conditions. After a final wash step, bound antibodies were detected by chemoluminescence (ECL-kit; Amersham-Pharmacia).
IMMUNOFLUORESCENCE MICROSCOPY
For indirect immunofluorescence, gastric biopsies were embedded in optimum cutting temperature compound, and frozen sections (4 μm) were cut on a cryostat and mounted on poly-D-lysine-coated slides. Two sections of each specimen were stained with haematoxylin-eosin for histological evaluation in parallel. The sections were postfixed with Carnoy reagent (60% ethanol, 30% chloroform, 10% acetic acid) for 20 minutes and washed in 96% ethanol for 10 minutes. To avoid non-specific binding of antibodies, sections were pretreated with 2% skim milk in phosphate buffered saline (PBS) for 20 minutes. After extensive washing with PBS, the slides were incubated with a primary antibody (diluted in PBS containing 0.1% bovine serum albumin (BSA)) in a humidified chamber at 4°C overnight. The sections were rinsed in PBS and incubated with the secondary antibody (diluted in PBS) at room temperature for 45 minutes. The sections were first washed in PBS, then in distilled water for five minutes, and dehydrated in 96% ethanol before mounting in Elvanol. For double label immunofluorescence microscopy, both primary and secondary antibodies were applied simultaneously. For negative controls, primary antibody was replaced by PBS. The immunostained samples were analysed using Zeiss Axiophot fluorescence microscope and a Zeiss LSM 410 confocal laser scanning microscope. For detection of positively stained cells, the whole section was carefully screened at least five times.
IMMUNOHISTOCHEMISTRY
Paraffin embedded samples of adenocarcinomas of the stomach (n=25) were analysed. Of 25 cases, one was graded as G1, six as G2, and the majority (n=18) as G3, in accordance with the WHO classification of 1990. The histopathological subclassification included papillotubular growth patterns, tubular carcinomas, and signet ring cell variants. Sections (5 μm) were mounted on slides and dried for at least 12 hours at 40°C. The sections were deparaffinised in xylol and subsequently submerged twice for five minutes in 100% and twice in 50% ethanol in TBS. Finally, the slides were incubated for 30 minutes at 100°C in 10 mM sodium citrate, pH 6.0. Staining was performed using the peroxidase/AEC rabbit/mouse Dako ChemMate detection kit (Dako, Hamburg, Germany). This involved a mild 10 minute proteinase K digestion followed by 30 minute incubations with the primary polyclonal anti-LI-cadherin antibody, the secondary streptavidin coupled antirabbit immunoglobulin antibody, peroxidase blocking solution, and biotinylated peroxidase. Staining was performed with the supplied chromogenic substrate and haematoxylin.
Results
LI-CADHERIN EXPRESSION IN BIOPSIES OF THE HUMAN STOMACH
In the gastrointestinal tract of healthy individuals, LI-cadherin is expressed in the small and large bowel but is completely absent from the oesophagus and stomach (fig 1). As expression of other intestinal proteins (for example, villin and sucrase-isomaltase) is known to be induced in the gastric mucosa under pathological conditions such as metaplasia and malignant transformation, we speculated that LI-cadherin may also be expressed under these circumstances. To test this hypothesis, we analysed 77 gastric biopsies from 30 consecutive patients, aged 28–90 years. At least two independent samples from every patient were evaluated by a histopathologist. Conventional histological analysis led to a diagnosis of intestinal metaplasia in seven of 30 patients (table 1).

Western blot analysis of LI-cadherin expression in the upper gastrointestinal tract. Protein extracts of biopsies (oesophagus, stomach, duodenum, ileum, and jejunum) and of the human colon carcinoma cell line Caco-2 were separated by sodium dodecyl sulphate-polyacrylamide gel electrophoresis and immunoblotted with polyclonal antiserum against human LI-cadherin.
Summary of patient data, diagnostic parameters, and immunoreactivity for LI-cadherin, villin, and sucrase-isomaltase in gastric biopsies
All gastric biopsies were subsequently analysed for expression of LI-cadherin by immunofluorescence microscopy. Of 77 biopsies (from 30 patients), 12 (from 10 patients) stained positive for LI-cadherin. These included all samples from patients diagnosed histopathologically to be affected by intestinal metaplasia as well as biopsies from three more patients initially rated negative.
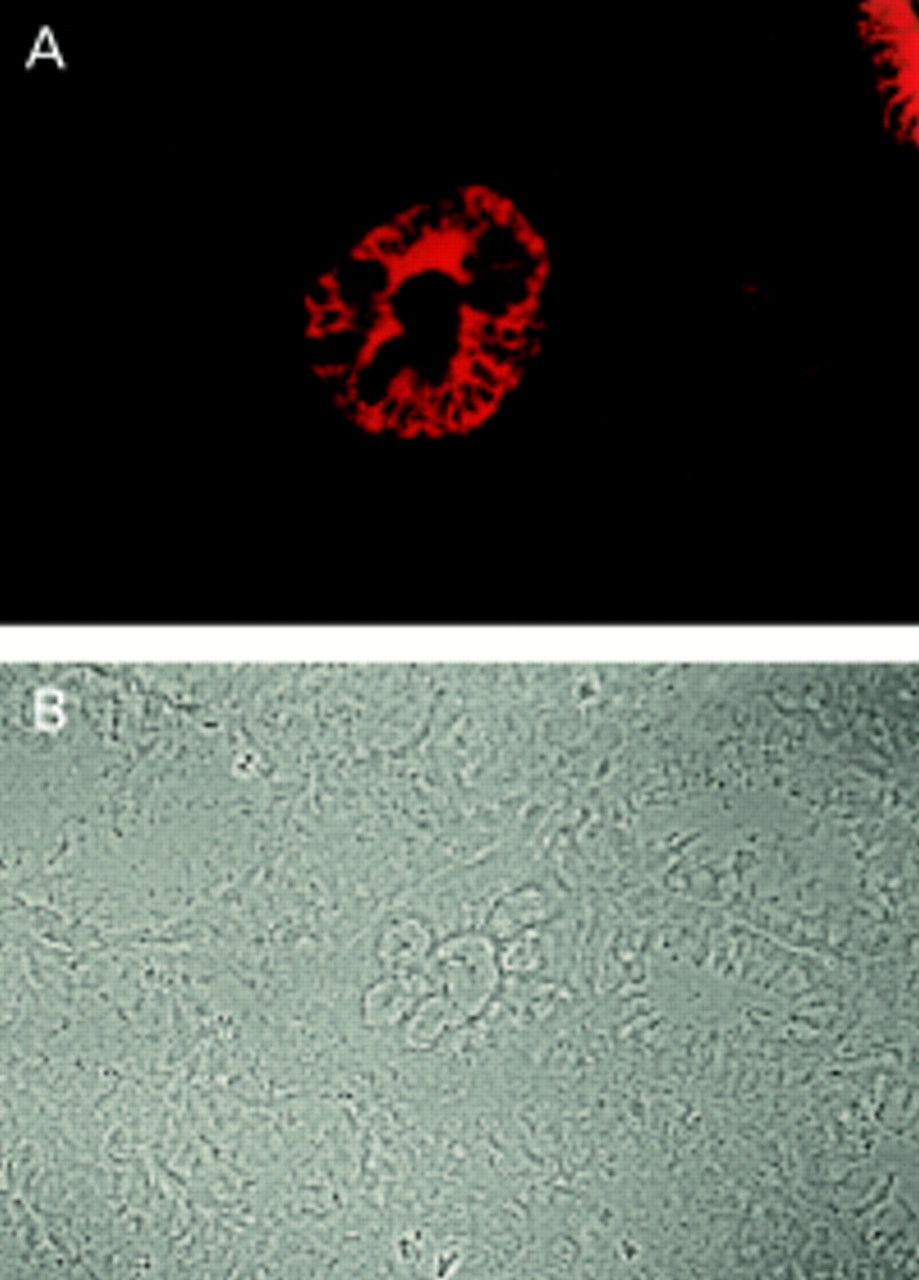
Staining of LI-cadherin was always limited to single glands or groups of glands and was never found in the whole mucosa (fig 2). Evaluation of LI-cadherin expression was facilitated by the low background of negative regions and the very strong signal of positive cells. Whereas in most patients only a few gastric glands stained positive for LI-cadherin, some patients showed more frequent staining and one patient had more than 70% of all glands expressing LI-cadherin. With respect to subcellular distribution, LI-cadherin was localised preferentially in the lateral domains of the plasma membrane. Both the apical plasma membrane and cytoplasm were negative.

Immunofluorescence microscopy of a gastric biopsy stained with polyclonal antibody against LI-cadherin: (A) fluorescence signal of LI-cadherin, (B) phase contrast. Note the distinct LI-cadherin staining of goblet cells in one gland whereas surrounding glands are negative.
COLOCALISATION OF LI-CADHERIN WITH SUCRASE-ISOMALTASE AND VILLIN IN GASTRIC BIOPSIES
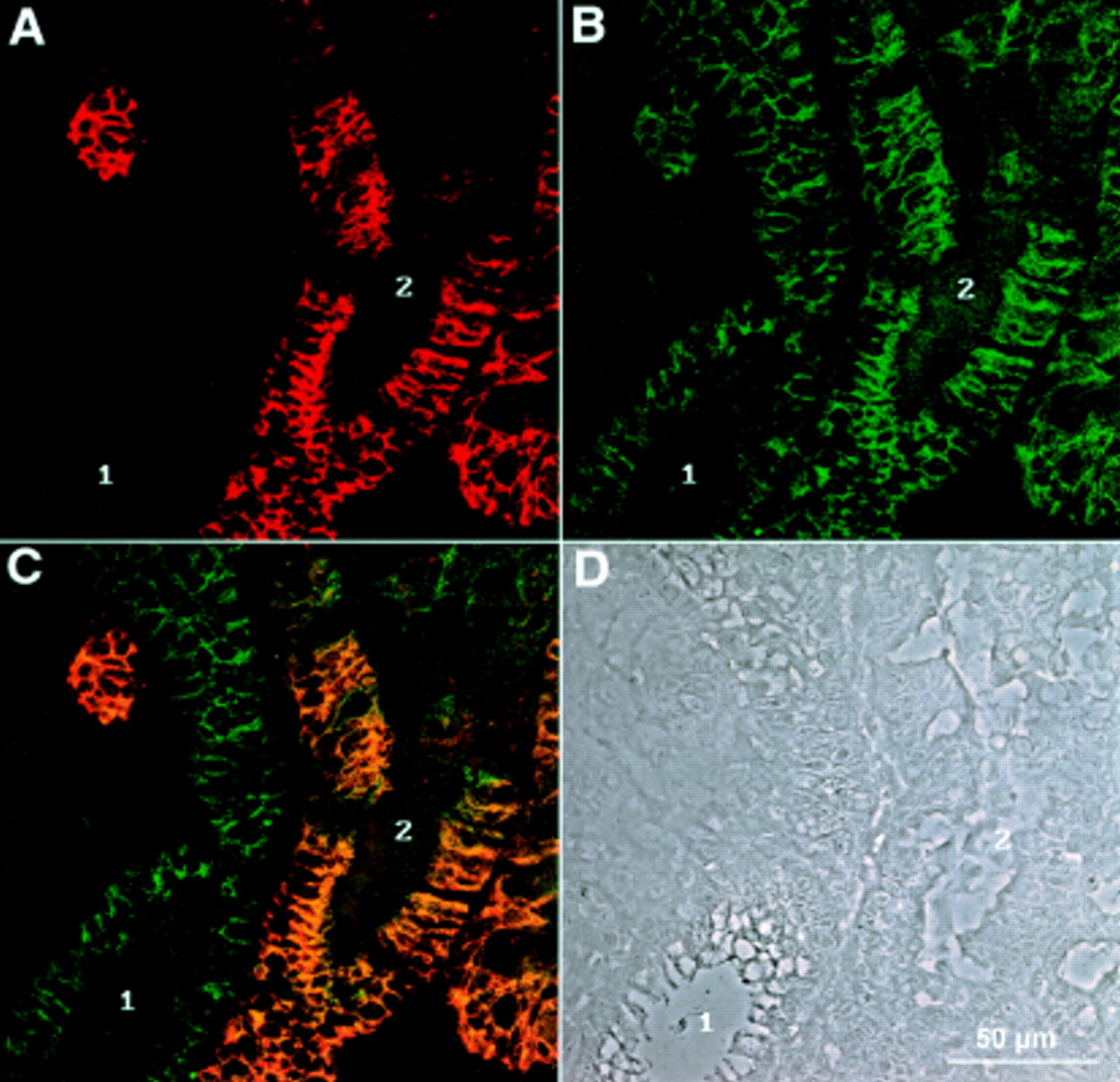
Immunoreactivity for LI-cadherin was compared with E-cadherin and β-catenin. Whereas E-cadherin and β-catenin (not shown) were observed—as expected—in all epithelial cells (fig 3B), LI-cadherin was only detected in a subpopulation of gastric glands (fig 3A). In the case of colocalisation of LI-cadherin with E-cadherin (fig 3C, yellow colour) and with β-catenin (not shown), almost complete overlap in the basolateral plasma membrane was observed. Non-epithelial cells (for example, vascular endothelium, nerve fibres, and connective tissue) showed no immunoreactivity, indicating the high specificity of the applied antibodies for epithelial cells.

Double immunofluorescence recorded by confocal laser microscopy of a gastric biopsy stained with polyclonal antiserum against LI-cadherin (A) and monoclonal antibody against E-cadherin (B). (C) Combined images of both proteins. The phase contrast image of the same area (D) reveals regions of normal mucosa (1) next to those showing characteristics of intestinal metaplasia (2). Note the high degree of colocalisation in cells expressing both LI-cadherin and E-cadherin.
Sucrase-isomaltase and villin represent established specific markers for the normal intestinal but not the normal gastric epithelium. Therefore, immunoreactivity of LI-cadherin was compared with immunostaining for sucrase-isomaltase and villin in both the mucosa of the small intestine (positive control, not shown) and the gastric mucosa (figs 4, 5). In biopsies of patients without intestinal metaplasia, LI-cadherin as well as villin and sucrase-isomaltase were not detected (not shown). However, in cases of gastric intestinal metaplasia diagnosed by conventional histology, immunoreactivity for all three intestinal epithelial marker molecules (that is, LI-cadherin, sucrase-isomaltase and villin) were observed (table 1; figs 4, 5).
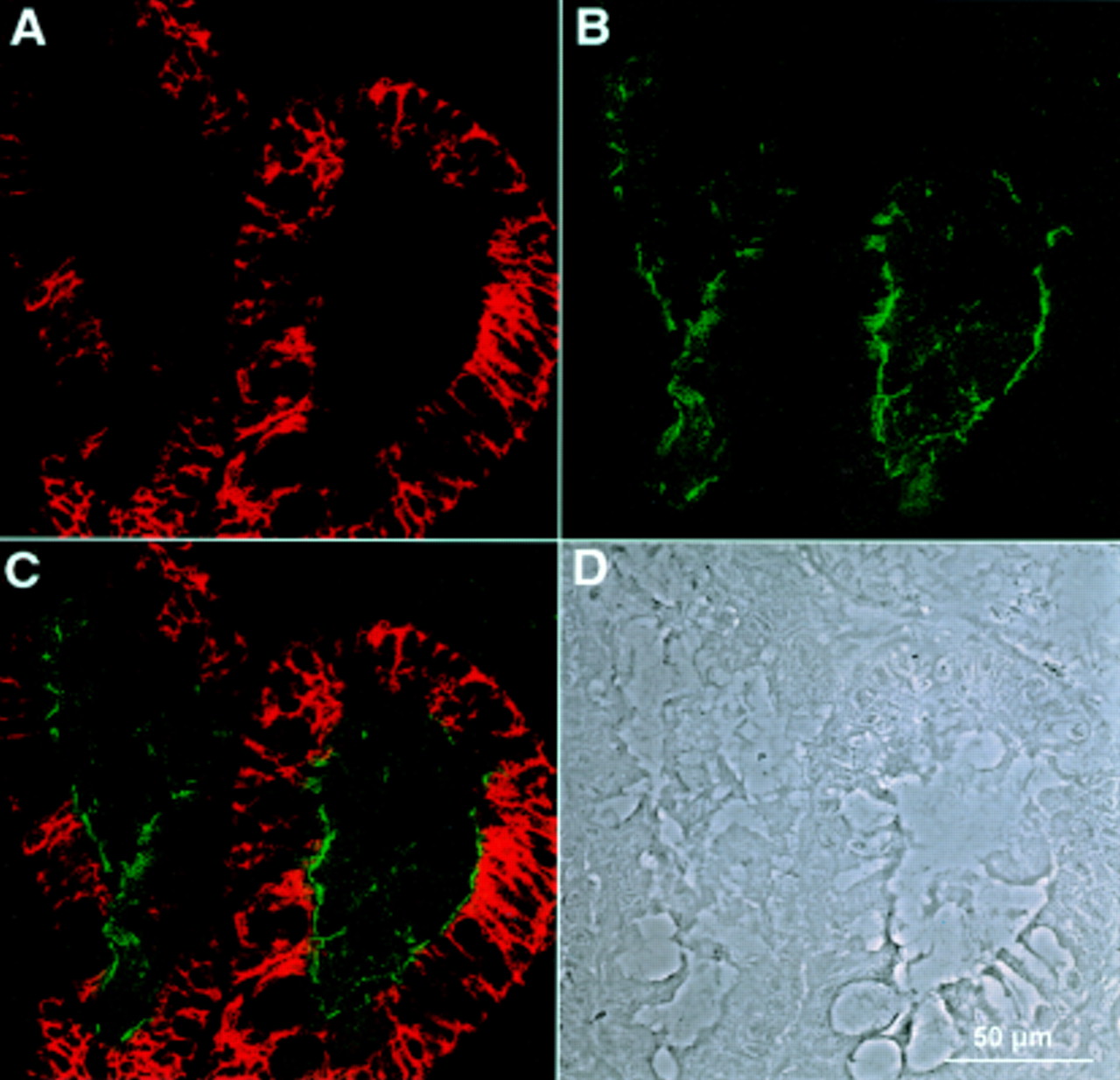
Double immunofluorescence recorded by confocal laser microscopy of a gastric biopsy with polyclonal antiserum against LI-cadherin (A) and monoclonal antibody against sucrase-isomaltase (B). (C) Combined images of both proteins. (D) Phase contrast image of the same area. Whereas LI-cadherin expression is restricted to the basolateral plasma membrane regions, sucrase-isomaltase is exclusively found in the apical regions and does not colocalise with LI-cadherin.

Immunofluorescence recorded by confocal laser microscopy of a gastric biopsy with polyclonal antiserum against LI-cadherin (A) and monoclonal antibody against villin (B). (C) Combined images of both proteins. (D) Phase contrast image of the same area. Whereas LI-cadherin background is very low in non-metaplastic regions (1, A), villin staining results typically in higher background staining (1, B). Connective tissue in between the glands is totally negative for both proteins (centre).
Interestingly, conventional histological diagnosis failed to detect gastric intestinal metaplasia in three of 10 patients. In contrast, only LI-cadherin and villin were positive in all those cases, whereas sucrase-isomaltase even failed to detect intestinal metaplasia in four of 10 patients (table 1). At the subcellular level, LI-cadherin did not colocalise with sucrase-isomaltase. Whereas sucrase-isomaltase was restricted to the apical plasma membrane domain, LI-cadherin was solely detectable on the basolateral surface (fig 4C). Villin is detectable in the apical membrane but also shows some overlap with LI-cadherin in the lateral but not basal membrane regions (fig 5).
LI-CADHERIN EXPRESSION IN HUMAN GASTRIC CARCINOMAS
To assess expression of LI-cadherin in gastric carcinomas, sections of 25 gastric carcinoma specimens were analysed by immunohistochemistry (fig 6). All but one of the gastric carcinomas exhibited intense immune staining for LI-cadherin. However, the staining intensity varied considerably between different areas within the same tumour. Nevertheless, LI-cadherin immunostaining facilitated discrimination between tumour cells and mesenchymal tissue and inflammatory infiltrates.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Immunoreactivity of LI-cadherin in gastric carcinomas. Immunohistochemical staining of LI-cadherin in mucinous (A), tubular (B), and signet ring (C) adenocarcinoma, and in undifferentiated carcinoma of the stomach (D). Note the intense staining of the cell-cell contact areas in well differentiated regions (1) of adenocarcinomas contrasted by weaker staining in less differentiated regions (2). The undifferentiated carcinoma (D) exhibits only faint cytoplasmic staining for LI-cadherin.
In gastric mucinous carcinoma, intense staining of LI-cadherin was found all along the lateral contact areas between well aligned polarised tumour cells (fig 6A). The apical face of the cells remained unstained. Similar localisation of LI-cadherin was revealed in tubular adenocarcinomas (fig 6B). Whereas in well ordered regions the aligned tumour cells exhibited strong expression of LI-cadherin along the cellular contact regions (1), single and less coherent tumour cells appeared to express LI-cadherin to a smaller extent (2). On comparing many different sections it becomes apparent that LI-cadherin staining is most intense in well differentiated gastric carcinomas of the intestinal type. In signet cell carcinomas, LI-cadherin is also predominantly found at the plasma membrane but staining was somewhat weaker than in adenocarcinomas of the intestinal type (fig 6C). The same holds true for diffusely infiltrating and undifferentiated gastric carcinomas. A particularly weak and diffuse staining, as is sometimes seen in those tumour sections, is displayed in fig 6D. Reduced or missing LI-cadherin staining is also observed in necrotic tumour areas.
Discussion
This study shows for the first time that LI-cadherin, an intestine specific cell adhesion molecule, is ectopically expressed in the metaplastic mucosa of the stomach as well as in gastric adenocarcinomas of the intestinal type.
All biopsies from patients diagnosed positive by the histopathologist as intestinal metaplasia also showed strong immunoreactivity for LI-cadherin. In addition, biopsies from patients missed by conventional histology were still identified as intestinal metaplasia by LI-cadherin immunofluorescence. These samples also stained positive for the established intestinal marker villin.28 This finding is best explained by the higher sensitivity of immunofluorescence detection compared with conventional light microscopy.
As LI-cadherin and villin antibodies always stained the same gastric glands and the same cells within, both proteins appear to be most sensitive in detecting the early stages of metaplasia—that is, when only a few glands per biopsy or just a few cells are affected. In contrast, routine histopathological examination of standard haematoxylin-eosin sections seems to miss subtle forms of intestinal metaplasia, as does immunofluorescence detection using sucrase-isomaltase antibodies, at least in some cases.
LI-cadherin proved to be superior to villin for two reasons. Firstly, it showed more intense cell-type specific immunoreaction than villin. This is best explained by the fact that LI-cadherin resides in the larger basolateral membrane compartments easier accessible for immunohistochemical analysis than apically located epitopes. Secondly, villin appears to be expressed at low levels in normal gastric epithelial cells30-32 leading to background staining. In contrast, LI-cadherin is not expressed in normal gastric epithelial cells resulting in a higher signal to noise ratio for LI-cadherin compared with villin.
The majority of biopsies positive for LI-cadherin showed only a focal staining pattern in certain glands. This finding is explained by the fact that the appearance of intestinal metaplasia is a process that starts with independent foci within the mucosa that merge at later stages.11 Within metaplastic glands, LI-cadherin is concentrated in the lateral contact areas between adjacent cells as has been observed for its expression in the intestinal mucosa.26 Indeed, the epithelial lining of metaplastic glands very much resembles that of normal intestinal crypts with respect to the elongated cell shape and presence of goblet cells.22 E-cadherin which is known to be expressed in most epithelia and to induce the formation of adherens junctions and thereby cell polarisation, almost perfectly colocalises with LI-cadherin in metaplastic glands. The observed lack of differences in expression between E-cadherin and β-catenin in normal and metaplastic gastric mucosa is in line with the findings of Shino and colleagues23 and Jawhari and colleagues33 who reported that the expression patterns of E-cadherin, β-catenin, plakoglobin, and α-catenin were not altered in gastritis, intestinal metaplasia, or gastric atrophy.
An antibody against villin, a 95 kDa protein present in the microvillus core, was used as a marker of the brush border.34 In agreement with Moll and colleagues30 we found only a weak luminal villin signal in the normal antral mucosa and even less in the corpus and fundus. Using antibodies against villin, Moll and colleagues30 and Osborn and colleagues28reported pronounced apical microvilli associated staining in intestinal metaplasia and gastric adenocarcinoma. Strong expression of villin in the gastric mucosa is therefore considered to be a marker of metaplastic or neoplastic changes of gastric epithelial cells. In the present study, exactly those epithelial foci were positive for LI-cadherin that also showed increased staining for villin.
Sucrase-isomaltase—a hydrolyase limited to the brush border membrane of absorbing intestinal cells35 ,36—served as the second marker protein for intestinally differentiated epithelium and revealed positive staining in double label immunofluorescence with LI-cadherin. In a subgroup of biopsies positive for LI-cadherin, sucrase-isomaltase could not be detected. This finding indicates potentially late or even missing induction of sucrase-isomaltase expression in relation to LI-cadherin and villin in certain metaplastic cells.
It is intriguing to speculate that LI-cadherin and villin, two typical intestinal proteins, appear to be expressed at an early stage of the metaplastic transition that is accompanied by subtle changes in cell morphology towards the intestinal cell type. As cadherins are known to be morphoregulatory proteins,14 it is conceivable that LI-cadherin may even be actively involved in those cellular transitions. Induction of LI-cadherin expression during intestinal metaplasia is paralleled by the observation that P-cadherin, which shows almost no immunoreactivity in normal gastric mucosa, becomes induced in well differentiated gastric adenocarcinomas.20 ,37 The authors speculate that induced expression of P-cadherin may participate in the morphogenesis of well differentiated adenocarcinomas.37
Similar to metaplastic cells, gastric carcinomas also showed strong expression of LI-cadherin in the cell-cell contact areas. If LI-cadherin expression should indeed be linked to the morphogenesis of tumour cells as proposed for P-cadherin, its induction should be one of the first steps of cellular reprogramming as metaplasia is regarded as a lesion that has the potential to slowly develop over many years into gastric carcinoma.
LI-cadherin expression seems to be reduced in less differentiated areas of adenocarcinomas and is weaker still in undifferentiated gastric carcinoma. This observation is in line with the reported reduced expression of P- and E-cadherin in poorly differentiated carcinomas exhibiting invasive growth.21 ,37 Moreover, Jawhari and colleagues33 observed that the progress of gastric epithelial metaplasia to dysplasia and finally to gastric carcinoma is associated with partial or total loss of the E-cadherin-catenin complex.
In conclusion, LI-cadherin represents a new marker molecule for both gastric metaplastic and gastric neoplasia. LI-cadherin staining yielded a lower background and thus a higher discriminatory power than villin, a well established marker of intestinal metaplasia. Finally, LI-cadherin clearly showed higher sensitivity than sucrase-isomaltase and may therefore become an important additional marker for gastric metaplastic and neoplasia.
Abbreviations used in this paper
- BSA
- bovine serum albumin
- IgG
- immunoglobulin G
- LI-cadherin
- liver-intestinal cadherin
- PBS
- phosphate buffered saline
- TBS
- Tris buffered saline
References
Footnotes
↵† Both authors contributed equally to this work.
Linked Articles
- Commentary