Article Text

Abstract
BACKGROUND Strains ofBacteroides fragilis producing a 20 kDa protein toxin (B fragilis toxin (BFT) or fragilysin) are associated with diarrhoea in animals and humans. Although in vitro results indicate that BFT damages intestinal epithelial cells in culture, the effects of BFT on native human colon are not known.
AIMS To examine the electrophysiological and morphological effects of purified BFT-2 on human colonic mucosa in vitro.
METHODS For resistance (R) measurements, colonic mucosa mounted in Ussing chambers was exposed to luminal or serosal BFT-2 (1.25–10 nM) and after four hours morphological damage was measured on haematoxylin and eosin stained sections using morphometry. F actin distribution was assessed using confocal microscopy.
RESULTS Serosal BFT-2 for four hours was four-, two-, seven-, and threefold more potent than luminal BFT-2 in decreasing resistance, increasing epithelial3H-mannitol permeability, and damaging crypt and surface colonocytes, respectively (p<0.05). Confocal microscopy showed reduced colonocyte F actin staining intensity after exposure to BFT-2.
CONCLUSIONS BFT-2 increases human colonic permeability and damages human colonic epithelial cells in vitro. These effects may be important in the development of diarrhoea and intestinal inflammation caused by B fragilis in vivo.
- B fragilis toxin
- toxin mediated colonocyte damage
- actin filaments
- transepithelial resistance
- morphometry
Abbreviations
- BFT
- Bacteroides fragilis toxin
- ETBF
- enterotoxigenicBacteroides fragilis
- Isc
- short circuit current
- PD
- potential difference
- R
- resistance
Statistics from Altmetric.com
- B fragilis toxin
- toxin mediated colonocyte damage
- actin filaments
- transepithelial resistance
- morphometry
The Gram negative anaerobic rod Bacteroides fragilis is associated with diarrhoea in animals1-9 and humans.10-13 Culture supernatants of enterotoxigenic B fragilis(ETBF) induce fluid secretion in calf, rabbit, and lamb ligated ileal loops,4 ,5 ,9 and ETBF causes fluid secretion and intestinal inflammation when administered orogastrically to newborn rabbits.8 ,9 Histological examination of ileal loops revealed that ETBF culture supernatants or microorganisms cause rounding and exfoliation of surface epithelial cells as well as mucosal inflammation.8 ,9 ,14 Similar secretory and histological changes have been observed in rat, rabbit, and lamb intestinal loops exposed to purified B fragilis toxin 1.15
Several studies suggest that the intestinal effects ofB fragilis are attributable to a 20 kDa protein toxin.14-17 Two isoforms of B fragilis toxin (BFT) have been isolated, one secreted by the lamb (ETBF strain VPI 1378414 ,17), and the second by the piglet (ETBF strain 86–5443–2–216; see also Wu and Sears, unpublished observations). Nucleotide sequence analysis studies revealed an 92% identity and 95% similarity in the amino acid sequences of these two isoforms of BFT17 (see also Wu and Sears, unpublished observations). Based on their biochemical properties, these isoforms represent two distinct proteins and were therefore termed BFT-1 (strain VPI 13784) and BFT-2 (strain 86–5443–2–2). Although detailed cross comparison studies are not yet complete, the biological activities of these two proteins are similar16-23 (see also Wu and Sears, unpublished observations). In vitro studies using human colonic adenocarcinoma cell lines indicate that ETBF and their toxins alter the structure and function of colonic epithelial cells. For example, exposure of subconfluent Caco-2, HT 29/C1, or HT 29 cells to either ETBF culture supernatants18 ,23 or purified toxins 1 or 219-21 ,23 ,24 causes cell rounding, swelling, and loss of cell to cell contacts. Furthermore, BFT-1 or BFT-2 also decrease transepithelial resistance when incubated with HT 29/C1, HT 29, T84, MDCK, or rat alveolar epithelial (type II) cell monolayers in vitro.18 ,24-26 Although a number of in vitro studies indicate that BFT-1 and BFT-2 affect the cell cytoskeleton of intestinal epithelial cells,17-21 ,26 the net F actin content of HT 29/C1 cells is unchanged after BFT treatment.20
Based on the above considerations, the present study was designed to examine the effect of purified BFT-2 on electrophysiology and morphology of human colonic mucosal strips mounted in Ussing chambers.
Methods
TOXIN PREPARATION
Bacteroides fragilis toxin 2 (BFT-2) was purified from culture supernatant broth of enterotoxigenicB fragilis 86–5443–2–2 as previously described.14 ,21 Purity of the toxin was assessed by analytical reverse phase fast protein liquid chromatography as previously described.21 Briefly, approximately 80 μg BFT-2 was injected onto a RepRPC reverse phase column (Pharmacia Inc., Piscataway, New Jersey, USA) equilibrated with water and 0.05% trifluoroacetic acid. Protein was eluted with a 0–100% gradient of acetonitrile and 0.05% trifluoroacetic acid over two hours. Eluted protein was detected by ultraviolet absorption at 226 nm. BFT-2 eluted as a single sharp peak at approximately 41% acetonitrile. Purified BFT-2 was maintained at −70°C in a 0.05 M Tris-0.18 M NaCl buffer (pH 7.5) until use. Biological activity of BFT-2 was tested using subconfluent HT 29/C1 monolayers (obtained from Daniel Louvard, Institute Pasteur, Paris, France) as previously described.21
EXPERIMENTAL DESIGN
Human colonic mucosal sheets were first equilibrated in Ussing chambers with buffer alone for 60 minutes as previously described.27 Serosal solution was then replaced with buffer containing 1.25–10 nM BFT-2 for four hours. In some experiments colonic tissues were incubated with luminal buffer containing 10 nM BFT-2 for four hours. Control tissues were incubated with buffer alone for five hours.
USSING CHAMBER MEASUREMENTS
In this study 51 individual specimens of histological tumour-free rectosigmoid colon were used. After removal of the seromuscular layer by blunt dissection one to five mucosal sheets from each specimen measuring 4–10 cm2 were mounted in Ussing chambers (DCTSYS, Precision Instrument Design, California, USA; 1.03 cm2 surface area) as previously described.27-30 The luminal and serosal sides were bathed at 37°C with a nutrient buffer containing: NaCl 122.0 mM/l, CaCl2 2.0 mM/l, MgSO4 1.3 mM/l, KCl 5.0 mM/l, glucose 20.0 mM/l, and NaHCO3 25.0 mM/l (pH 7.4 when gassed with 95% O2/5% CO2; temperature 37°C). The upper level of fluids in both luminal and serosal reservoirs was identical. Potential difference (PD) and short circuit current (Isc) were continuously measured and registered every 10 minutes as previously described.27Luminal and serosal solutions were connected via Ag-AgCl electrodes to a voltmeter (Model VCC600, Revision G, Voltage-Current Clamp, Physiologic Instruments, San Diego, California, USA). Resistance (R) was calculated using Ohm’s law from the open circuit PD andIsc. PD values were given in mV (lumen negative), Isc in μA/cm2, and R in Ohm.cm2. PD values were corrected for the junction potentials (less than 0.1 mV) between the luminal and serosal solutions.2 ,30
Epithelial permeability to 3H-mannitol was determined as previously described.27 Three human colonic explants from a single individual were mounted in Ussing chambers in parallel and incubated with either buffer alone or serosal or luminal buffer containing 10 nM BFT-2 for four hours (n=5, paired tissues). After an equilibration period of 30 minutes, 3H-mannitol (26.4 Ci/mmol; Dupont-New England Nuclear, Boston, Massachusetts, USA) was added to 8 ml serosal buffer at a final concentration of 0.32 nM. Luminal aliquots of 200 μl were taken for liquid scintillation counting using 5 ml Quicksave A scintillation fluid (Zinser, Maidenhead, UK) and replaced with 200 μl buffer to eliminate a transepithelial solute gradient. The radioactivity of3H-mannitol in the luminal fluid was measured in counts per minute (cpm), determined every 30 minutes for up to four hours in the luminal or serosal presence or absence (control) of 10 nM BFT-2 (n=5).
MORPHOMETRY
After the Ussing chamber experiments, tissues were processed for light microscopy. Mucosal damage of surface and crypt epithelium was assessed by a histopathologist (RS), who performed morphometry on coded, paraffin wax embedded, haematoxylin and eosin stained slides as previously described.27 ,28 ,30 The histopathologist was not informed about the experimental conditions to which the tissues had been exposed. The surface area of each tissue sample was 1×1 cm. Morphometry was performed on nine vertical sections from different locations, each representing the total height and length (1 cm) of the mucosal preparation. Thus a total length of 8–9 cm of mucosal surface was examined for each tissue. A Leitz Diaplan research microscope (Wild Leitz Ltd, Heerbrugg, Switzerland; objective magnification, ×4, ×6.3, ×16, ×25, ×40; ocular magnification, ×12.5), a Panasonic colour CCD videocamera (model WV-CD 130/G, Matshushita Com. Ind. Co. Ltd, Japan) an analog/digital monitor screen (model PVM-1271Q, superfine pitch, Sony, Japan), a personal computer (IBM-PS/2, model PS2; IBM, Armonk, New York, USA), extended by insertion of a frame grabber (ITI PCVision plus board; Imaging Technology Corp., Woburn, Massachusetts, USA), and a graphic tablet (Summasketch plus 12 in × 12 in MM 1201; Summagraphics Corp., Fairfield, Connecticut, USA) were used. Downloading MIPSY (the micro based image processing system), real time morphometry was performed. The extent of damaged crypt or surface epithelium was separately measured in μm and expressed as percentage of total surface or crypt epithelium.
Histological criteria for epithelial damage were: reduced staining intensity of epithelial cells, condensation, fragmentation and dissolution of the nucleus, cell disruption, formation of subepithelial blebs, and lifting of cells from the basal lamina.27 ,28 ,30 No tissue strip used in these experiments showed histological criteria of malignancy or inflammation.
FLUORESCENT MICROSCOPY
At the end of the Ussing chamber experiments tissues were processed for fluorescent microscopy. Fluorescent staining of F actin was performed using fluorescein isothiocyanate (FITC) labelled phalloidin (Molecular Probes, Inc.) on fresh frozen tissues. Sections (4 μm) were fixed in 2% paraformaldehyde, 0.01 M sodium periodate, 0.075 M lysine, and 0.37 M sodium phosphate (pH 7.4), washed in Tris buffered saline (TBS, pH 7.5) and incubated for one hour at room temperature in TBS containing 1% normal donkey serum and 3% bovine serum albumin (TBS block). Sections were then incubated for one hour at room temperature in FITC-phalloidin (Sigma; 10 μg/ml in TBS block). Subsequently, slides were mounted with glycerol:PBS (1:9) andn-propyl-gallate (1 mg/ml) and examined and photographed using a laser confocal microscope (Carl Zeiss Inc., Thornwood, New York, USA). For each experimental condition five tissues derived from five individual specimens were investigated.
STATISTICS
All data were expressed as mean (SEM) and probabilities were regarded as significant if they reached a 95% level of confidence (p<0.05) using the Student’s t test for paired and unpaired observations. The statistical test compared morphometric, electrophysiological (PD, Isc, R), and permeability values of controls versus experimental tissues at defined time points and baseline values of PD,Isc, and R versus values obtained after one and four hours of toxin exposure.
Results
EFFECT OF TOXIN ON ELECTROPHYSIOLOGY
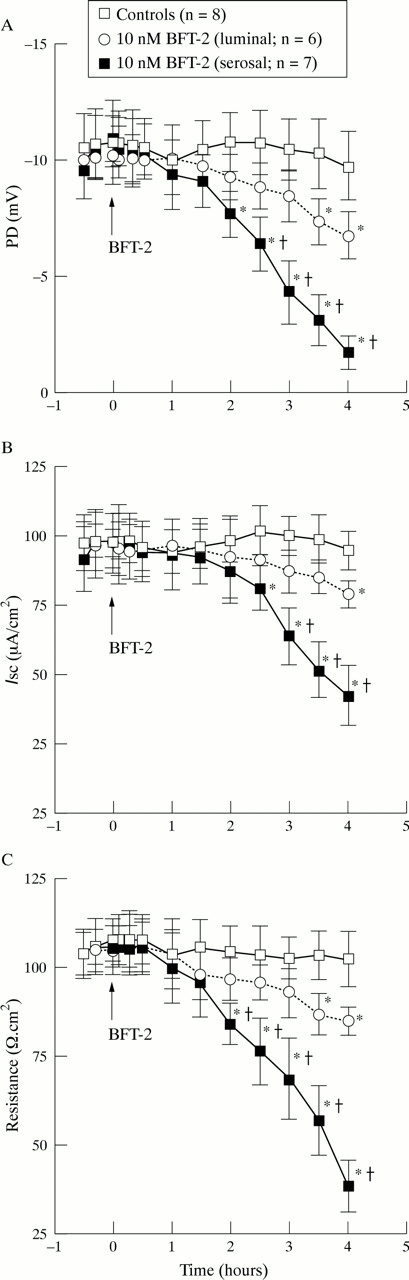
The effects of BFT-2 on colonic electrophysiology were compared using PD, Isc, and R. BFT-2 caused time dependent changes in electrophysiological parameters which were more notable when the toxin (10 nM) was applied on the serosal aspect of tissues versus the luminal side (fig 1). Serosal application of BFT-2 caused a PD increase towards less negative values and R decrease which became statistically significant two hours after toxin exposure (p<0.05 versus controls). PD and R values continued to increase and decrease, respectively, until the end of the four hour observation period. Serosal BFT-2 also caused a decline inIsc which became statistically significant 2.5 hours after toxin exposure (p<0.05 versus controls). ThereafterIsc continued to decrease until the end of the incubation period (fig 1). In contrast, luminal exposure of colonic mucosa to BFT-2 caused a less steep change in electrophysiological parameters which became significantly different from controls only after 3.5 hours of continuous toxin exposure for PD and R and after four hours for Isc (p<0.05; fig 1).

Time course of the effect of BFT-2 on (A) colonic potential difference (PD), (B) short circuit current (Isc), and (C) resistance. Values represent mean (SEM); n = number of experiments. *p<0.05 versus controls; †p<0.05 versus luminal BFT.
The effect of different doses of BFT-2 was tested on electrophysiological parameters. As shown in table 1 and fig 2, PD,Isc, and R remained stable for up to four hours in control strips incubated with buffer alone. In contrast, serosal exposure of mucosal strips to 10, 5, and 2.5 nM BFT-2 for four hours caused a significant, dose dependent PD increase andIsc and R decrease as compared with buffer exposed tissues as well as to the baseline value of the same colonic strip prior to toxin exposure (p<0.05). Incubation of tissues with 1.25 nM serosal BFT-2 did not affect colonic electrophysiology. On a molar basis, serosal BFT-2 was approximately three to four times more potent than luminal BFT-2 in producing a similar drop in resistance (fig 2).
Effect of BFT-2 on electrophysiology of human colonic mucosa

Dose dependent effect of BFT-2 on colonic resistance. Values represent mean (SEM); n = number of experiments. *p<0.05 versus controls.
Permeability to 3H-mannitol
The effect of luminal or serosal administration of 32 nM BFT-2 on serosal to mucosal permeability to 3H-mannitol was studied. As shown in fig 3, continuous serosal or luminal administration of 10 nM BFT-2 for four hours caused a 4.7-fold and 2.2-fold increase in permeability, respectively, when compared with controls (p<0.01). Permeability changes became statistically significant after two and 3.5 hours after serosal and luminal exposure to BFT-2, respectively, when compared with controls (p<0.05).

Effect of BFT-2 on colonic 3H-mannitol permeability. Values are mean (SEM) of five experiments performed in triplicate. *p<0.05, **p<0.01 versus controls.
EFFECT OF TOXIN ON HISTOLOGY
Histological examination of colonic strips incubated with buffer alone for five hours showed normal colonic architecture (fig 4A). Serosal and luminal administration of BFT-2 caused similar morphological changes in crypt and surface epithelium which were time and dose dependent. Following 2.5 hours of serosal incubation with 10 nM BFT-2, histology revealed damage of cells at the crypt base whereas the surface epithelium remained intact (fig 4B). However, after four hours severe damage of both crypt and surface colonocytes was observed (fig 4C). BFT-2 caused cell rounding and detachment of crypt enterocytes into the crypt lumen (long arrows in fig 4D). In surface epithelial cells BFT induced apical localisation of the pyknotic nucleus, and cytoplasmic elongation and luminal protrusion of the enterocyte (arrowheads in fig 4D). Injured enterocytes were separated from neighbouring cells and detached into the lumen. Furthermore, serosal BFT-2 caused loosening and disruption of the basal lamina (broad arrows in fig 4D). Luminal addition of 10 nM BFT-2 for four hours was less effective in damaging surface and crypt epithelium and did not cause alterations of the basal lamina (not shown).

Morphological effect of BFT-2 on human colonic mucosa. Human colonic mucosal sheets were incubated with either buffer alone for four hours (A) or serosal buffer containing 10 nM BFT-2 for 2.5 hours (B) or four hours (C, D). At the end of the incubation period sheets were fixed in formalin and processed for light microscopy. (A) Control tissue appears normal and epithelial cells form a continuous sheet over the lamina propria. (B) The mucosal sheet shows disruption of crypt epithelium whereas the surface epithelium remains intact. (C) The mucosal sheet shows disruption of surface and crypt epithelium. The arrowhead marks an area shown at higher magnification in (D) which shows rounding and detachment of crypt enterocytes into the lumen (long arrows). Surface colonocytes show cytoplasmic elongation and apical localisation of the pyknotic nucleus (arrowheads). Note loosening of the basal lamina (broad arrows). Original magnification × 140 (A, B); × 80 (C); × 420 (D).
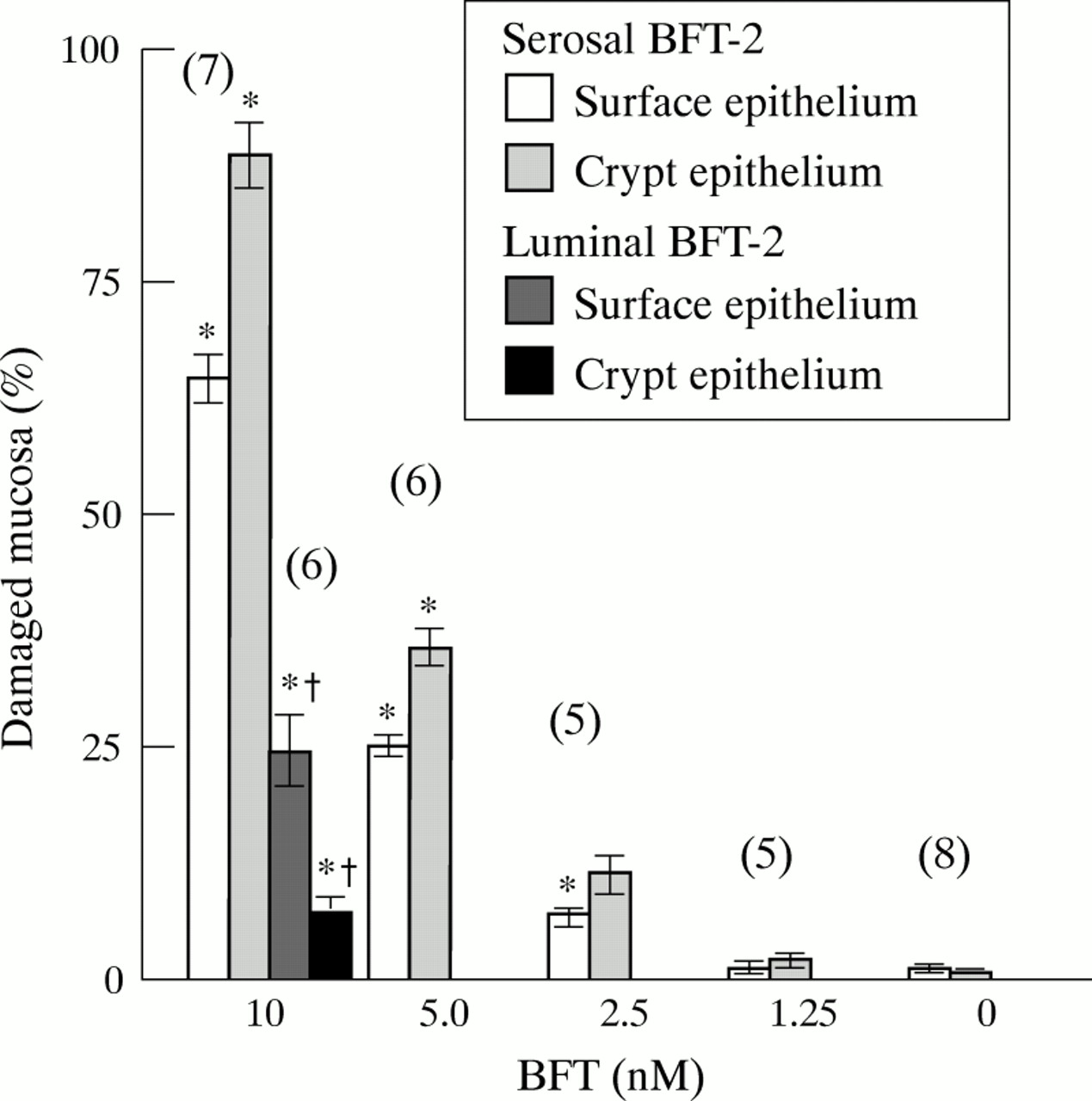
BFT-2 induced morphological damage of crypt and surface epithelium was dose dependent (fig 5). Serosal administration of 10 nM BFT-2 was approximately seven and three times more potent at damaging crypt and surface epithelium, respectively, when compared with luminal BFT-2.
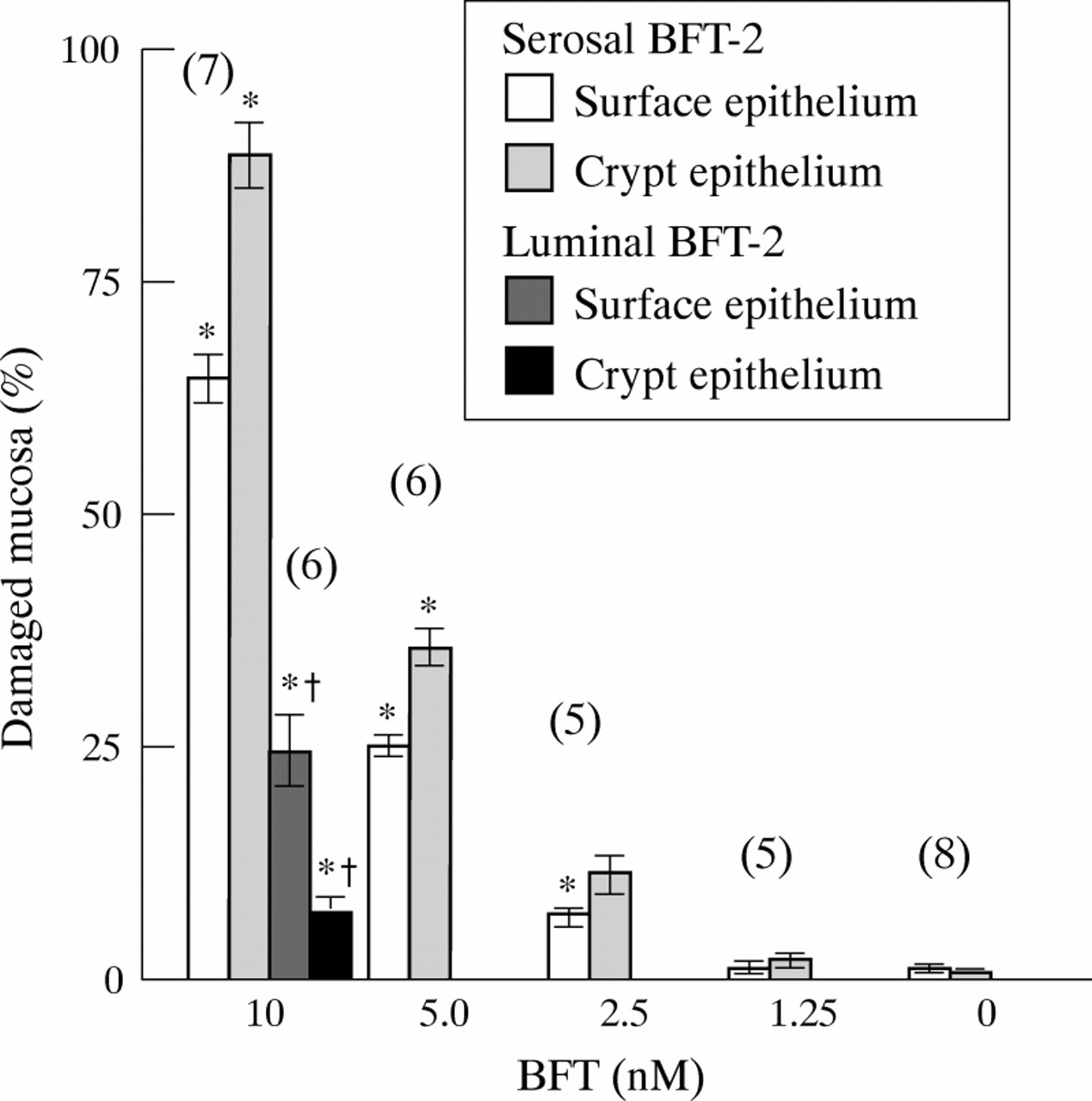
Morphometric analysis of colonic epithelial cell damage caused by BFT-2. Values are mean (SEM). The number of experiments is given in brackets. *p<0.05 versus controls; †p<0.05 versus serosal 10 nM BFT-2.
DISTRIBUTION OF F ACTIN
Previous studies showed that BFT-2 causes alterations in enterocyte F actin.20 Thus, the effect of BFT-2 on F actin distribution in human colonic mucosa was examined, using fluorescent phalloidin to detect F actin. As shown in fig 6A, control cells had a polygonal shape with F actin distributed in the peripheral actin ring. Furthermore, control enterocytes showed intense F actin staining of the apical brush border membrane (arrowheads in fig 6A). As shown in fig6B, serosal exposure to 10 nM BFT-2 for two hours resulted in an overall reduction in F actin staining intensity and disruptions of the F actin ring (arrowheads in fig 6B). After four hours of serosal exposure to 10 nM BFT-2, staining of the enterocyte F actin ring was nearly abolished (fig 6C,D). However, some F actin staining remained in the brush border of damaged enterocytes (EC in fig 6D) or in exfoliated enterocytes (arrowheads in fig 6D). Effects on enterocyte F actin were similar, but less profound after luminal administration of BFT-2 (10 nM) (not shown). In tissue exposed to toxin and used for staining experiments (fig 6B–D) PD increased and R andIsc decreased during Ussing chamber experiments, while control tissues (fig 6A) exhibited stable electrophysiological values throughout the experiment.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Effect of BFT-2 on cellular F actin. Fluorescent photomicrograph of human colonic mucosal sheet incubated with buffer alone (A) or serosal buffer containing 10 nM BFT-2 for two hours (B) or four hours (C, D). Samples were fixed and fresh frozen sections cut perpendicularly (B) or vertically (A, C, D) to the long axis of surface cells. (A) Control cells had a polygonal shape with F actin distributed in the peripheral actin ring. Arrowheads mark intense F actin staining of the apical brush border. L, lumen; LP, lamina propria. (B) There was an overall reduction in F actin staining intensity and disruption of the F actin ring (arrowheads). (C) There was an overall reduction in F actin staining in epithelial cells (EC). The rectangle marks a crypt shown at higher magnification in (D), and the asterisk marks the basal lumen of the crypt. (D) Staining of the enterocyte F actin ring was nearly abolished. Some F actin staining remained in the brush border of damaged enterocytes (EC) or in exfoliated enterocytes (arrowheads). BL, basal lamina. Original magnification × 400 (A, B); × 200 (C); ×480 (D).
Discussion
Purified BFT-2 caused electrophysiological alterations, increased epithelial permeability to mannitol, and morphological damage to human colonic mucosa.
The BFT-2 induced morphological damage of colonocytes was characterised by rounding and exfoliation, which was paralleled by an increase in transepithelial 3H-mannitol flux, a sensitive measure of tight junction permeability. Myers and colleagues8 ,9showed that luminal administration of B fragilis causes rounding and exfoliation of surface and crypt epithelial cells of rabbit colon in vivo. Weikel and colleagues23 showed that culture medium of ETBF induces cell rounding and loss of cell to cell contact in subconfluent T84, Caco2, and HT 29/C1 cells. Similar morphological changes in HT 29 or HT 29/C1 cells in vitro were evident after treatment with purified BFT-1 or BFT-2.14 ,18 ,20 ,24 We have recently observed that BFT-1 or BFT-2 causes dose dependent disruptions of tight junctions and zonula adherens in T84 cell monolayers.25 Furthermore, Obiso and colleagues26 recently showed that BFT-1 causes a decrease in the immunofluorescent staining intensity of the tight junction protein ZO-1 in MDCK cells in vitro. In addition, Moncrief and colleagues17 showed that BFT-1 is a metalloprotease which hydrolyses actin, fibrinogen, and collagen in vitro. Therefore, in addition to loosening cell to cell contacts, digestion of the basal lamina might contribute to increased damage after serosal administration of BFT-2. Taken together, these results indicate that the interaction of BFTs with epithelial cells leads to disruption of intercellular junctions and/or cell−basal laminal binding, resulting in increased paracellular permeability.
Serosal BFT-2 administration was more potent than luminal administration at inducing electrophysiological, permeability, and morphological changes. In agreement with this finding, in vitro studies using polarised intestinal22 ,24-26 and non-intestinal epithelial cell lines26 have shown that BFT-1 and BFT-2 reduce monolayer resistance and that this decline in resistance is polar, occurring more quickly with application of the toxin to the basolateral, rather than the apical surface of the monolayers. Taken together, the results of the present and recent studies22 ,24-26 indicate that BFT-2 may act via a receptor(s) or a target molecule localised on both apical and basolateral membranes of surface and crypt enterocytes. We speculate that the polarity dependency of BFT-2 induced biological effects may be due to either easier access of this toxin to target molecule(s) or receptors localised on the basolateral side of the epithelial cells, and/or that BFT-2 may also interact with subepithelial inflammatory cells, nerve cells or fibroblasts. Release of proinflammatory cytokines from these cells may contribute to the biological responses following BFT-2 administration.
Our results did not show electrophysiological changes indicative of BFT-2 induced secretion. In contrast, Chambers and colleagues25 found that BFT-2 induces an early increase inIsc in T84 cell monolayers that was attributable to chloride secretion. The discrepancy between the results presented here and those of Chambers et almay reflect differences between a malignant cell line used by Chamberset al and the normal colonic mucosal strips used in this study.
As shown by confocal microscopy of FITC-phalloidin stained sections, BFT-2 almost completely abolished F actin staining in damaged enterocytes, except for some intracellular and apical condensations of F actin. Interestingly, this result differs from the effects of BFT on F actin in intestinal epithelial cells in vitro. In intact HT 29 or HT 29/C1 cells BFT seems to stimulate actin rearrangement without cleavage of F actin in vitro.18 ,20 ,24 Furthermore Saidi and colleagues20 showed that incubation with BFT-1 for up to 24 hours did not alter net actin content in HT 29/C1 cells in vitro. However, an increased F action staining intensity could be observed on the periphery of HT 29/C119 ,20 and HT 2918cells after two to three hours of with incubation with BFT. Modulation of F actin distribution in intestinal epithelial cells is not unique toB fragilis. For example,Clostridium difficile toxins A and B cause disruption and condensation of F actin without completely abolishing F actin staining in intestinal epithelial cells27 and fibroblasts.31-33 The zonula occludens toxin ofVibrio cholerae 34 and heat stable enterotoxin of Escherichia coli 35 were also shown to cause rearrangement of F actin in rabbit ileum and T84 cell monolayers, respectively.
In summary, the present study shows that BFT-2 increases human colonic permeability and damages colonic epithelial cells in vitro. These results may be relevant to the pathophysiological mechanism by whichB fragilis causes diarrhoea and intestinal inflammation in vivo.
Acknowledgments
This study was supported by grants from the Jubiläumsfonds der Österreichischen Nationalbank, the Anton Dreher-Gedächtnisschenkung des medizinischen Dekanates der Universität Wien, and, in part, by NIH grant DK 45496 to CS. MR was supported by a grant from the Max Kade Foundation, New York, and M L and IC were supported by a Research Carrier Development Award from the Crohn’s and Colitis Foundation of America, Inc. We thank Dr Shaoquang Wu for purification of the BFT-2 used in these studies and Ingrid Hammer for her expert technical support.
Abbreviations
- BFT
- Bacteroides fragilis toxin
- ETBF
- enterotoxigenicBacteroides fragilis
- Isc
- short circuit current
- PD
- potential difference
- R
- resistance