Article Text

Abstract
BACKGROUND The pathogenesis of pancreatic fibrosis is unknown. In the liver, stellate cells play a major role in fibrogenesis by synthesising increased amounts of collagen and other extracellular matrix (ECM) proteins when activated by profibrogenic mediators such as cytokines and oxidant stress.
AIMS To determine whether cultured rat pancreatic stellate cells produce collagen and other ECM proteins, and exhibit signs of activation when exposed to the cytokines platelet derived growth factor (PDGF) or transforming growth factor β (TGF-β).
METHODS Cultured pancreatic stellate cells were immunostained for the ECM proteins procollagen III, collagen I, laminin, and fibronectin using specific polyclonal antibodies. For cytokine studies, triplicate wells of cells were incubated with increasing concentrations of PDGF or TGF-β.
RESULTS Cultured pancreatic stellate cells stained strongly positive for all ECM proteins tested. Incubation of cells with 1, 5, and 10 ng/ml PDGF led to a significant dose related increase in cell counts as well as in the incorporation of3H-thymidine into DNA. Stellate cells exposed to 0.25, 0.5, and 1 ng/ml TGF-β showed a dose dependent increase in α smooth muscle actin expression and increased collagen synthesis. In addition, TGF-β increased the expression of PDGF receptors on stellate cells.
CONCLUSIONS Pancreatic stellate cells produce collagen and other extracellular matrix proteins, and respond to the cytokines PDGF and TGF-β by increased proliferation and increased collagen synthesis. These results suggest an important role for stellate cells in pancreatic fibrogenesis.
- pancreatic fibrosis
- stellate cell activation
- cytokines
Abbreviations
- ECM
- extracellular matrix
- GFAP
- glial fibrillary acidic protein
- PGDF
- platelet derived growth factor
- PGDF-R
- platelet derived growth factor receptor
- SMA
- smooth muscle actin
- TGF
- transforming growth factor
- PLSD
- Fisher’s protected least significant difference test
Statistics from Altmetric.com
Pancreatic fibrosis is a characteristic pathological feature of chronic pancreatitis.1 However, the mechanisms responsible for the development of fibrosis in the pancreas are not known and, to date, have received little attention.
In contrast to the pancreas, fibrogenesis in the liver has been well studied. It is now established that the hepatic stellate cell plays a central role in the production of hepatic fibrosis.2-4 In the normal liver, stellate cells are the primary storage site for vitamin A.2 In liver injury, these cells are activated by various profibrogenic mediators including cytokines5(released from Kupffer cells and activated macrophages) and oxidant stress.6 ,7 The process of stellate cell activation involves: (a) enlargement and proliferation; (b) transformation into a myofibroblast-like phenotype exhibiting positive staining for the cytoskeletal marker protein α smooth muscle actin (αSMA); (c) increased responsiveness to proliferative and fibrogenic cytokines (such as platelet derived growth factor (PDGF) and transforming growth factor β (TGF-β) respectively) via increased expression of relevant receptors on the cell surface; and (d) increased synthesis and secretion of individual extracellular matrix (ECM) components including collagens, fibronectin, laminin, proteoglycans, and hyaluronan. It has been shown that activated hepatic stellate cells produce the major fraction of ECM proteins during hepatic fibrogenesis, with only minor contributions (if any) from hepatocytes, Kupffer cells, and bile duct epithelial cells.8
It is possible that pancreatic fibrosis is produced by a pathway similar to that in the liver, namely via activation of pancreatic stellate cells. This concept is strengthened by recent studies from our group showing that: (a) the normal rat pancreas contains stellate shaped cells (identified by positive staining for the cytoskeletal markers desmin and glial fibrillary acidic protein (GFAP)), which are morphologically similar to hepatic stellate cells9; and (b) in a rat model of pancreatic fibrosis and in human chronic pancreatitis, positive staining for αSMA is observed (suggesting the presence of activated stellate cells) in fibrotic areas.10 A method has been developed to isolate and culture pancreatic stellate cells,9 providing a novel in vitro method to study their biology. In culture, pancreatic stellate cells exhibit features identical to those described for cultured hepatic stellate cells including: abundant lipid droplets in the cytoplasm, positive autofluorescence for vitamin A, and positive staining for desmin and GFAP.
As described earlier, the morphology of pancreatic stellate cells has now been well characterised both in vivo and in vitro. In normal rat pancreas these cells exist in a quiescent, non-activated state as shown by negative staining for αSMA.9 By analogy with hepatic stellate cells, activation of pancreatic stellate cells might be expected to occur under conditions of pancreatic injury when there is increased production of profibrogenic factors such as cytokines11 ,12 and oxidant stress.13 ,14 If pancreatic stellate cells play a role in pancreatic fibrogenesis in a manner analogous to stellate cells in the liver, it is essential to show that these cells have the ability to produce ECM proteins in response to the profibrogenic mediators. However, these functional aspects of pancreatic stellate cells have not yet been defined. Therefore, the aims of this study were to determine whether cultured pancreatic stellate cells: (a) have the capacity to produce ECM proteins; and (b) are activated by cytokines such as PDGF and TGF-β1.
Methods
ISOLATION AND CULTURE OF PANCREATIC STELLATE SHAPED CELLS
Rat pancreatic stellate cells were isolated as detailed previously,9 by a modification of the method described by Schafer et al in the liver.15Briefly, the pancreas was digested with a mixture of collagenase P (0.05%), pronase (0.02%), and DNAse (0.1%) in Gey’s balanced salt solution. The resultant suspension of cells was centrifuged in a 13.2% Nycodenz gradient at 1400 g for 20 minutes. Stellate cells separated into a fuzzy band just above the interface of the Nycodenz solution and the aqueous buffer. This band was harvested, and the cells were washed and then resuspended in Iscove’s modified Dulbecco’s medium containing 10% fetal bovine serum, 4 mM glutamine, and antibiotics (penicillin 100 units/ml; streptomycin 100 μg/ml). The above technique yields a preparation of stellate cells devoid of contamination by endothelial cells or macrophages as evidenced by negative staining for the markers factor VIII and ED1 respectively.
Freshly isolated stellate cells were cultured in 24 well culture plates (for cytokine experiments) or Nunc chamber slides (for immunocytochemistry).
IMMUNOCYTOCHEMISTRY FOR ECM PROTEINS (PROCOLLAGEN III, COLLAGEN I, FIBRONECTIN, AND LAMININ)
Cells cultured in Nunc chamber slides were washed in Hank’s balanced salt solution (HBSS), fixed in acetone at 4°C for 10 minutes, and air dried. After washing for five minutes with three changes of 1× Tris buffered saline (TBS) at room temperature, cells were incubated with 1% H2O2−methanol for 30 minutes to block endogenous peroxidase activity. After further washes, cells were exposed to a blocking solution to prevent non-specific binding of antibody as follows: cells to be examined for procollagen III, laminin, and fibronectin were incubated with TBS containing 10% goat serum and 1% bovine serum albumin (BSA) whereas those to be examined for collagen I were incubated with TBS containing 10% swine serum and 1% BSA. Cells were then incubated with the primary antibody (polyclonal rabbit antiprocollagen III antibody diluted 1 in 20, polyclonal rabbit anticollagen I antibody diluted 1 in 50, polyclonal rabbit antilaminin antibody diluted 1 in 30, or polyclonal rabbit antifibronectin antibody diluted 1 in 50) in the appropriate blocking solution overnight at 4°C. After three washes in TBS for five minutes each, cells to be tested for procollagen III, laminin, and fibronectin were incubated for 60 minutes at room temperature with peroxidase conjugated goat antirabbit IgG diluted 1 in 100 in TBS containing 10% goat serum and 1% BSA. Cells tested for collagen I were incubated with alkaline phosphatase conjugated swine antirabbit IgG diluted 1 in 100 in TBS containing 10% swine serum and 1% BSA. Colour was developed using the Sigma liquid diaminobenzidine (DAB) substrate-chromogen system (for procollagen III, laminin, and fibronectin) and the Sigma liquid fast red-naphthol substrate-chromogen system for collagen I. Cells were then counterstained with Mayer’s haematoxylin for five minutes. Stellate cells incubated with the appropriate preimmune host serum served as negative controls.
DETERMINATION OF CELL PROLIFERATION IN RESPONSE TO PDGF
Cultured pancreatic stellate cells were passaged twice and replated at equal seeding densities into 24 well culture plates. Triplicate wells of cells were then exposed to the PDGF isoform PDGF-BB in increasing concentrations (1, 5, and 10 ng/ml culture medium) for 48 hours. Cells incubated in medium without PDGF-BB served as controls. The rate of cell proliferation after exposure to PDGF-BB was assessed using two parameters—cell counts per well and rate of DNA synthesis as measured by the incorporation of 3H-thymidine into trichloroacetic acid (TCA) precipitable material.
Cell counts
Cells were washed twice in HBSS, harvested by trypsinisation using 0.5% trypsin−0.2% EDTA, resuspended in 200 μl culture medium, and counted using a haemocytometer.
DNA synthesis (incorporation of 3H-thymidine)
For the final 16 hours of the 48 hour incubation with PDGF-BB, cells were pulsed with 3H-thymidine at a concentration of 5 μCi per well. The reaction was stopped by aspirating the medium and cells were washed twice with HBSS. Ice cold 10% TCA was added to the wells and cells incubated for 10 minutes at 4°C. This was followed by a further two incubations with 5% TCA for 10 minutes each at 4°C. A 500 μl aliquot of 1 N NaOH was added to the wells which were shaken on a rotary shaker at room temperature for 30 minutes. The reaction was stopped by adding 500 μl 1 N HC1. The mixture was transferred to scintillation vials and radioactivity of the samples was measured using a liquid scintillation counter (Tricarb 4000 Series, Canberra Packard, Five Dock, NSW, Australia).
ASSESSMENT OF STELLATE CELL ACTIVATION IN RESPONSE TO TGF-β1
Cells at passage 3, plated at equal seeding density in 24 well culture plates, were used for TGF-β1 studies. Triplicate wells of cells were incubated with TGF-β1 at concentrations of 0.25, 0.5, and 1 ng/ml culture medium for 48 hours at 37°C in a 5% CO2−air humidified atmosphere. Cells incubated without TGF-β1 were used as controls. Stellate cell activation was assessed by determination of expression of αSMA and by measurement of collagen synthesis. The effect of TGF-β1 on the expression of PDGF receptors, specifically the β subunit of the PDGF receptor (PDGF-Rβ), on the surface of pancreatic stellate cells was also examined.
αSMA expression
Concentrations of αSMA in cells incubated with and without TGF-β1 were estimated by western blotting of cell lysate proteins, using a monoclonal mouse antibody to αSMA. Briefly, cells were harvested by trypsinisation, and cell lysates were obtained by incubating the cells overnight in lysis buffer (50 mM Tris HCl (pH 7.5), 150 mM NaCl, 1% Triton X-100, 0.5% deoxycholate, 1% sodium dodecyl sulphate (SDS), 1 mM Na orthovanadate, 2 mM EDTA, 10 mM NaF, 10 μg/ml aprotinin, 10 μg/ml leupeptin, 1 mM phenylmethylsulphonyl fluoride).16 Samples were then centrifuged at 2200g for 10 minutes and the supernatant was harvested for measurement of αSMA concentrations.
Immunoblotting for αSMA—Protein concentrations in cell lysates were measured by the method of Lowryet al 17 using BSA as the standard. Proteins (15 μg) from each sample were separated by gel electrophoresis using a 15% SDS-polyacrylamide gel. Known molecular weight protein standards were run alongside the samples. Separated proteins were transferred onto a nitrocellulose membrane using a commercial semidry blotting apparatus (Biorad, Richmond, California, USA). The nitrocellulose membrane was then incubated at room temperature with 5% skimmed milk powder in TBS (pH 7.6) for one hour to prevent non-specific binding of antibody. This was followed by an overnight incubation at 4°C with the primary antibody (monoclonal mouse anti-αSMA antibody diluted 1 in 200) in fresh blocking solution. The filter was then washed and incubated with the secondary antibody (goat antimouse IgG diluted 1 in 500; Amersham International, Buckinghamshire, UK) for 60 minutes at room temperature. αSMA bands were detected by the enhanced chemiluminescence technique using the Amersham ECL kit and quantitated by densitometry of scanned autoradiographs (Scanalytics One-D Scan, Boston, Massachusetts, USA). Densitometry readings were expressed as integrated optical densities (arbitrary densitometer units calculated from the density as well as the size of each band) per μg protein loaded onto the membranes.
Collagen synthesis
Collagen synthesis was assessed in cells incubated with and without TGF-β1 by measuring the incorporation of radiolabelled proline into collagenase sensitive protein, using a modification of the method of Agelli and Wahl.18 Briefly, cells were incubated for one hour in proline-free medium and then labelled for 16 hours with 2 μCi/ml 14C-proline in Dulbecco’s medium containing 1% fetal bovine serum, 50 μg/ml ascorbic acid (freshly made), and 75 μg/ml β-aminoproprionitrile. At the end of the incubation period, culture medium in the wells was aspirated and kept aside. Cells were harvested by trypsinisation, washed, and added to the reserved medium. The samples were sonicated for 90 seconds and proteins were precipitated with 10% TCA (final concentration). TCA treated samples were centrifuged at 1000 g for 10 minutes and the pellet (comprising precipitated protein) was solubilised in 200 μl 0.2N NaOH. After neutralisation with 0.2 N HCl, samples were divided into two equal aliquots. One aliquot was incubated with collagenase solution (containing 6.25 mM CaCl2, 70 mM Tris (pH 7.4), 30 mM N-ethylmaleimide (NEM) and 360 units/ml collagenase type III) whereas the other aliquot was incubated with the above solution in the absence of collagenase for 120 minutes at 37°C. Incorporation of 14C-proline into collagen and non-collagen protein was determined following precipitation with TCA. Collagen incorporated radioactivity was recovered in the TCA soluble fraction whereas non-collagen radioactivity was found in the TCA precipitate. Collagen production was determined using formulae derived by Peterkofsky et al 19 and was expressed as counts per minute (cpm) per μg DNA.
Expression of PDGF-Rβ
Cells cultured on Nunc chamber slides (four separate cell preparations) were incubated in duplicate with or without 1 ng/ml TGF-β1 for 48 hours. The cells were then immunostained. for PDGF-Rβ according to the protocol described earlier. The primary antibody used was polyclonal rabbit anti-PDGF-Rβ diluted 1 in 25. Horseradish peroxidase conjugated goat antirabbit IgG (diluted 1 in 100) was used as the secondary antibody and colour was developed using the Sigma liquid DAB substrate-chromogen system. Cells incubated with non-immune rabbit serum served as negative controls.
PROTEIN AND DNA DETERMINATIONS
Protein content of cell lysates was measured by the method of Lowry et al 17 using BSA as the standard. Pancreatic DNA was assayed by a modified fluorimetric microassay as described by Kapuscinski and Skoczylas20using calf thymus DNA as the standard.
STATISTICAL ANALYSIS
Results are expressed as mean (SEM) for five separate cell preparations per experimental protocol. Data were analysed using repeated measures analysis of variance (ANOVA).21Fisher’s protected least significant difference (PLSD) was used for comparison of individual groups provided that the F test was significant.21 ,22 The analyses were run on a Macintosh LC 630 personal computer using the Statview II statistical program.
MATERIALS
All general chemicals were of analytical reagent grade and were purchased from Sigma Chemical Company (St Louis, Missouri, USA). Collagenase P was purchased from Boehringer Mannheim (Mannheim, Germany), collagenase type III (from Clostridium histolyticum) and protease type XIV (fromStreptomyces griseus) were obtained from Sigma Chemical Company, and DNAse was purchased from Pharmacia Biotech (Uppsala, Sweden). Cell culture reagents were purchased from Sigma Chemical Company. Nycodenz was obtained from Nycomed Pharma AS (Oslo, Norway). Iscove’s modified Dulbecco’s medium was purchased from Gibco BRL. Antibodies were obtained from the following sources: polyclonal rabbit antihuman type III procollagen (Chemicon International, Temecula, California, USA), polyclonal rabbit antirat collagen type I (Amrad Pharmacia Biotech, Victoria, Australia), polyclonal rabbit antimouse laminin (Sigma Chemical Company), polyclonal rabbit antirat fibronectin (Calbiochem-Novabiochem Corp., San Diego, California, USA), and monoclonal antibody to αSMA (Sigma Chemical Company).3H-thymidine (specific activity 6.7 Ci/mmol) was purchased from ICN Pharmaceuticals (Costa Mesa, California, USA), and14C-proline (specific activity 250 mCi/mmol) was obtained from NEN (Boston, Massachusetts, USA). Human recombinant PDGF-BB and TGF-β1 were purchased from Sigma Chemical Company.
ETHICS APPROVAL
This study was approved by the Animal Care and Ethics Committee of the University of New South Wales, Australia.
Results
IMMUNOSTAINING FOR ECM PROTEINS
Pancreatic stellate cells in late primary culture and early passage exhibited strong positive staining for all ECM proteins tested—procollagen III, collagen I, fibronectin, and laminin (fig 1). Staining for procollagen III and collagen I and laminin was predominantly perinuclear, whereas that for fibronectin was spread more diffusely in the cytoplasm.

Pancreatic stellate cells in culture stained strongly positive for procollagen III, collagen I, laminin, and fibronectin (panels B, D, F, and H respectively) compared with control cells incubated with the appropriate non-immune sera (panels A, C, E, and G respectively). Staining for the collagens and for laminin was perinuclear in distribution; that for fibronectin was diffuse.
EFFECT OF PDGF-BB ON CELL PROLIFERATION
Cell counts
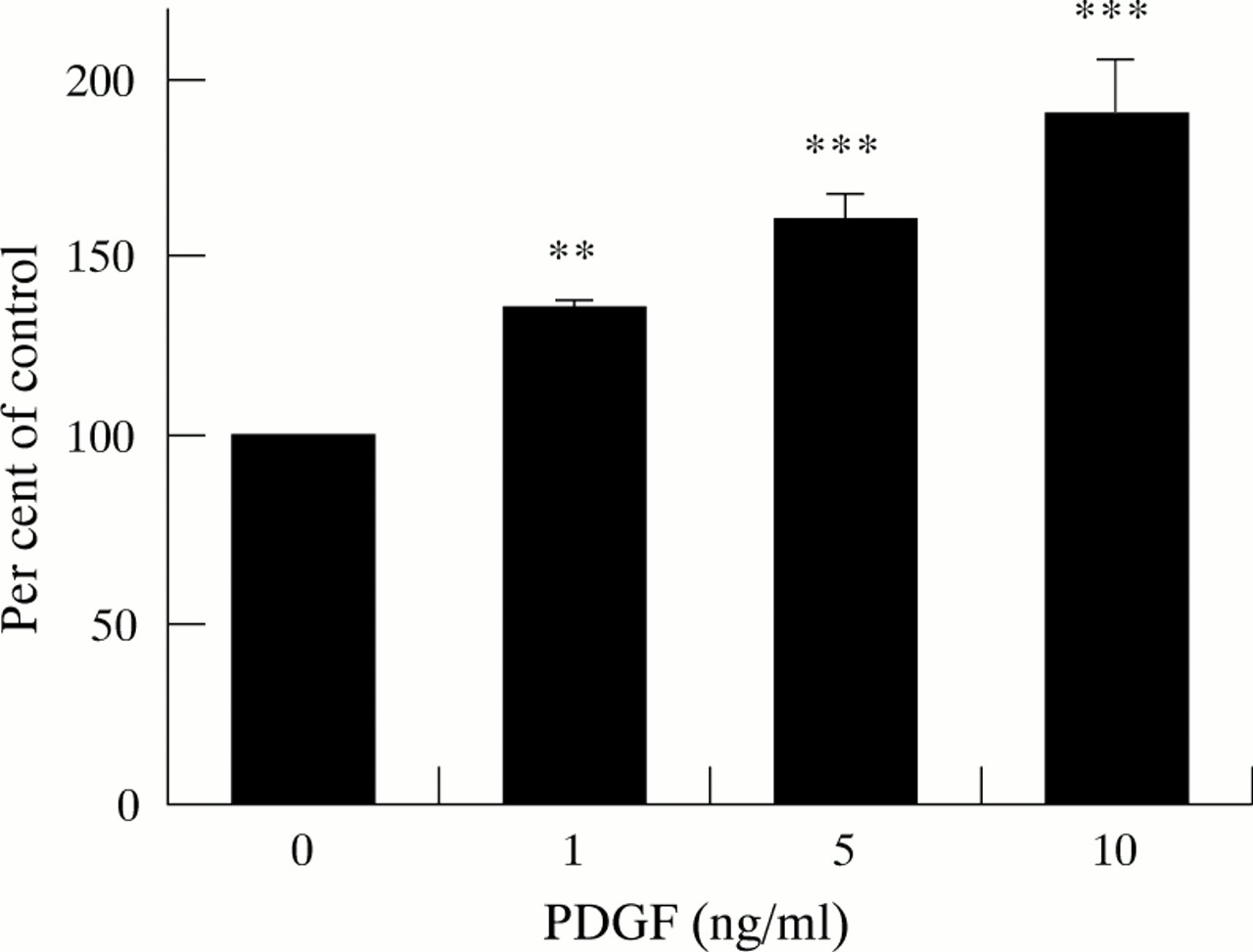
Increasing concentrations of PDGF-BB resulted in a dose dependent, statistically significant increase in cell numbers compared with control cells not incubated with PDGF. As depicted in fig 2, at a PDGF concentration of 10 ng/ml, cell numbers were increased to 237 (13.1)% of control values.

Effect of PDGF on cell counts (five separate cell preparations). Results are expressed as a percentage of control values (cells not incubated with PDGF). Increasing concentrations of PDGF resulted in a dose related increase in cell numbers (*p<0.025, †p<0.001).
3H-thymidine incorporation
Similarly to the results obtained with cell counts, incorporation of radiolabelled thymidine was significantly increased in a dose related manner in cells exposed to increasing concentrations of PDGF-BB (fig 3), indicating an increased rate of DNA synthesis in response to PDGF.

Effect of PDGF on DNA synthesis (five separate cell preparations). DNA synthesis was estimated by measuring the incorporation of 3H-thymidine into TCA precipitable material. Results are expressed as a percentage of control values observed in cells not incubated with PDGF. **p<0.01, ***p<0.001.
EFFECT OF TGF-β1 ON CELL ACTIVATION
αSMA expression
Immunoblotting for αSMA revealed a single band in each lane corresponding to the known molecular weight of αSMA (42 kilodaltons). A representative immunoblot in fig 4 shows the notable, dose dependent increase in αSMA protein expression observed with increasing concentrations of TGF-β1. Densitometry of autoradiographs of immunoblots revealed a statistically significant increase in αSMA concentrations in cells incubated with TGF-β1 compared with control cells (table 1).

Western blot analysis of cell lysates for αSMA expression. The figure shows a representative immunoblot for expression in cells incubated with and without TGF-β1. A single band was detected in each lane corresponding to the known molecular weight (42 kDa) of αSMA.
Effect of TGF-β1 on α SMA concentrations in pancreatic stellate cells
Collagen synthesis
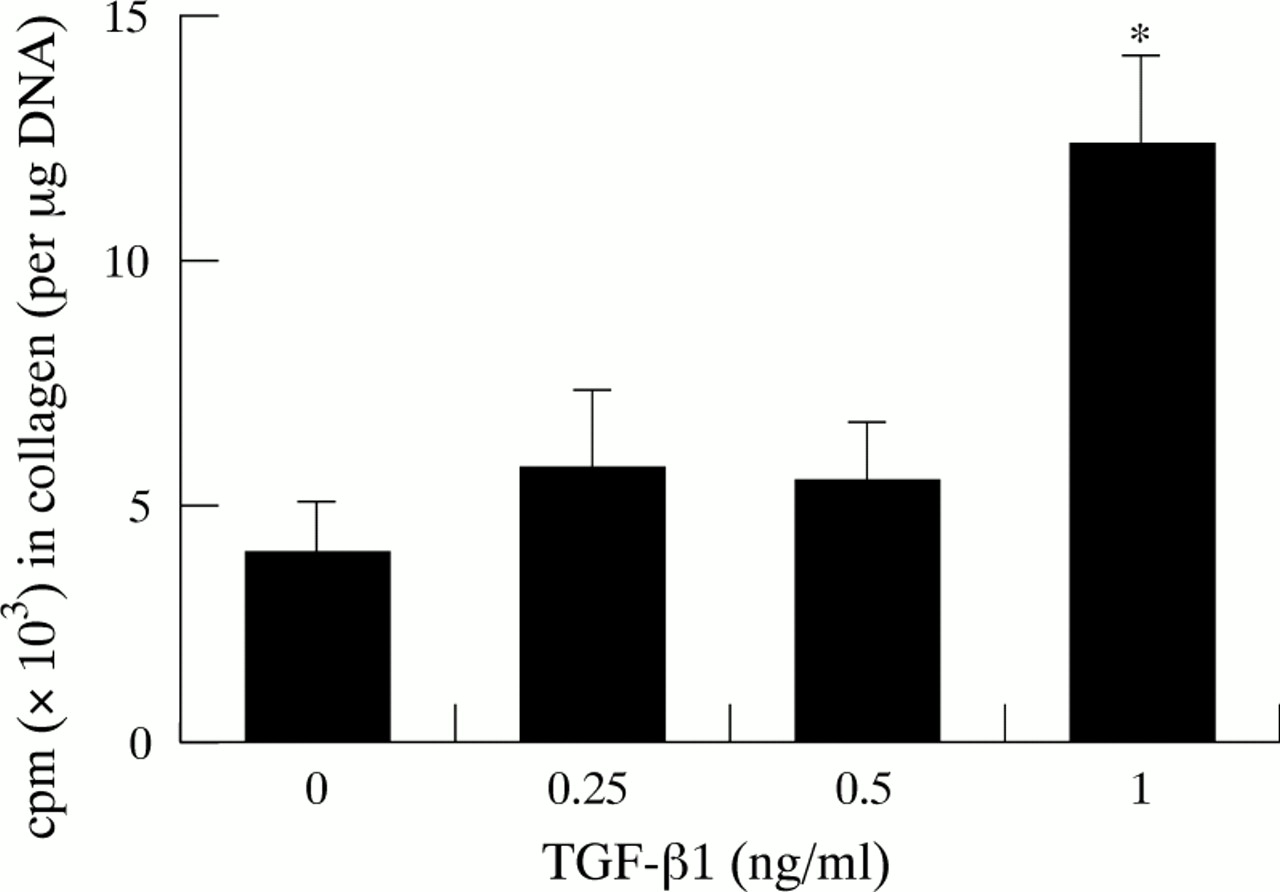
Collagen synthesis was expressed as cpm (counts per minute) in collagen per μg DNA (fig 5). Stellate cells incubated with TGF-β1 showed increased rates of collagen production compared with controls. Although the increases in collagen synthesis with the lower concentrations of TGF-β1 (0.25 and 0.5 ng/ml) were modest, a significant increase in collagen synthesis was observed in cells incubated with 1 ng/ml TGF-β1.

Effect of TGF-β1 on collagen synthesis (five separate cell preparations). *p<0.005.
Expression of PDGF-Rβ
Stellate cells stained positively for PDGF-Rβ regardless of whether or not they were exposed to TGF-β1. However the intensity of staining was notably greater in cells incubated with 1 ng/ml TGF-β1 (fig 6).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Effect of TGF-β1 on expression of the β subunit of the PDGF receptor (PDGF-Rβ). Panels A and B show stellate cells (not exposed to TGF-β1) incubated with non-immune serum (negative control) and an antibody to PDGF-Rβ respectively. Panels C and D show TGF-β1 treated stellate cells incubated with non-immune serum (negative control) and an antibody to PDGF-Rβ respectively.
Discussion
This study showed that cultured rat pancreatic stellate cells have the capacity to produce collagen and other ECM proteins and are activated (exhibit increased proliferation, αSMA expression, and collagen synthesis) when exposed to the cytokines PDGF and TGF-β. In these respects, pancreatic stellate cells are similar to liver stellate cells (which are known to be central to hepatic fibrogenesis). This supports the notion that pancreatic stellate cells may play a role in the development of fibrosis in the pancreas.
Pancreatic fibrosis is a common response to chronic pancreatic injury from various causes including recurrent and/or persistent inflammation, alcohol abuse, malnutrition, and trauma.1 However, the pathogenesis of this constant feature of chronic pancreatitis has not yet been elucidated. In general, fibrosis represents a pathological change in the composition and amount of ECM in tissues.2 ,4 Fibrous/scar tissue consists of fibril forming collagens (types I and III) and other glycosylated proteins such as proteoglycans, fibronectin, and laminin. The present study has shown that pancreatic stellate cells in late primary culture and early passage stain strongly positive for collagen types I and III, fibronectin, and laminin—that is, they have the capacity to produce the proteins that constitute fibrous tissue. Procollagen III staining was also observed in early primary culture, at a time when collagen I staining was absent (data not shown). This observation is similar to that reported in cultured hepatic stellate cells where quiescent cells (in early culture) were found to produce predominantly collagen III whereas collagen I production was only observed in activated cells in late culture.23 Positive staining for laminin in pancreatic stellate cells also indicates transformation of these cells to a myofibroblast-like phenotype in culture, as distinct from fibroblasts which do not synthesise laminin.24
To support the hypothesis that pancreatic stellate cells play an active role in pancreatic fibrogenesis, it is essential to establish not only the capacity of these cells to produce ECM proteins (described earlier), but also their ability to respond to profibrogenic mediators. The latter has been clearly shown in this study by testing the effects of the cytokines PDGF and TGF-β1.
PDGF, a ubiquitous growth regulatory peptide, is the most potent proliferative mediator of hepatic stellate cells.25 ,26During tissue repair and inflammatory processes, PDGF is secreted at high concentrations by various cells including platelets, mononuclear cells, and activated macrophages.27 PDGF is a dimerised protein consisting of two polypeptide chains (A and/or B) linked via disulphide bonds, resulting in three isoforms, PDGF-AA, PDGF-BB, and PDGF-AB. With respect to hepatic stellate cells, the isoforms PDGF-BB and PDGF-AB have been shown to display greater mitogenicity than PDGF-AA.28 In the present study, the isoform PDGF-BB was selected for use as preliminary experiments had shown that PDGF-AA had little effect on pancreatic stellate cell proliferation (data not shown). The isoform PDGF-BB was found to have a significant proliferative effect on pancreatic stellate cells as assessed by both cell counts and DNA synthesis. The mechanism(s) by which PDGF exerts this mitogenic effect on pancreatic stellate cells was not examined in this study. It is possible that, as has been shown in hepatic stellate cells and in other cell systems, PDGF has a stimulatory effect on cell signalling pathways such as phosphoinositide turnover and intracellular Ca2+ release.28-30 Exposure of pancreatic stellate cells to PDGF in vivo is likely to occur in conditions of pancreatic inflammation characterised by the presence of cell types such as platelets and macrophages that can synthesise and secrete peptide growth factors and cytokines. PDGF induced stellate cell proliferation in vivo may be further aided by the upregulation of PDGF receptors on the surface of stellate cells. In this regard, it is of interest to note that in a rat model of pancreatic fibrosis, immunostaining for PDGF receptor (known to bind the isoform PDGF-BB) was found to be notably increased in association with areas of fibrosis.10 This observation is in accord with the reports of increased PDGF receptor mRNA levels in cirrhotic human livers31 and in rat liver following acute CCl4induced injury.32
The second cytokine examined in the present study was TGF-β, which is a homodimer composed of two identical subunits linked via disulphide bonds.33 Five homologous isoforms of this protein have been described; the most abundant isoform in the liver is TGF-β1.34 This isoform of TGF-β plays a regulatory role in the biosynthesis and turnover of the ECM and is the most potent fibrogenic mediator described for hepatic stellate cells.35 Transformation to a myofibroblast-like phenotype and stimulation of collagen synthesis have been shown in cultured hepatic stellate cells exposed to TGF-β1.36 The role of TGF-β1 in pancreatic disease has been studied in experimental models of acute pancreatitis as well as in human chronic pancreatitis. Repeated injections of TGF-β1 after recurrent episodes of acute pancreatitis in mice have been reported to promote the development of pancreatic fibrosis.37 In a rat model of caerulein induced pancreatitis, increased protein and mRNA levels of TGF-β1 were observed (during the regeneration phase) associated with collagen deposition and increased mRNA levels for collagen types I and III.38 Furthermore, inhibition of TGF-β1 by administration of a neutralising antibody in the above rat model was found to reduce collagen synthesis and ECM deposition.39In human chronic pancreatitis, TGF-β1 expression has been shown to be increased in association with areas of fibrosis.40
In the present study, we have shown that TGF-β1 induces the transformation of pancreatic stellate cells to myofibroblasts (as evidenced by increased αSMA expression) and also stimulates collagen synthesis in these cells. These results suggest that the observed effect of TGF-β on pancreatic collagen synthesis during pancreatic injury in vivo may be mediated via activation of stellate cells in the gland. An additional observation made in this study was the induction of PDGF receptor expression in cells exposed to TGF-β1. This observation is similar to that described in stellate cells cultured from human liver where expression of PDGF-Rβ was increased at both the mRNA and protein level in cells exposed to 1 ng/ml TGF-β1.16 TGF-β1 induced upregulation of PDGF receptors could have implications for the perpetuation of fibrogenesis in vivo. Increased expression of cell surface receptors would result in increased responsiveness of stellate cells to PDGF and a consequent perpetuation of the activated state of these cells.
In conclusion, the results of this study provide information regarding potential mechanisms leading to the development of pancreatic fibrosis. We have shown that the pancreas contains cells that are similar, not only morphologically but also functionally, to the cells known to be primarily responsible for fibrogenesis in the liver. Thus the findings of this study provide support for the concept that pancreatic stellate cells, when activated by profibrogenic mediators, have the potential to play an important role in pancreatic fibrogenesis.
Abbreviations
- ECM
- extracellular matrix
- GFAP
- glial fibrillary acidic protein
- PGDF
- platelet derived growth factor
- PGDF-R
- platelet derived growth factor receptor
- SMA
- smooth muscle actin
- TGF
- transforming growth factor
- PLSD
- Fisher’s protected least significant difference test