Article Text

Abstract
BACKGROUND Murine T cell mediated acute semiallogeneic graft versus host disease (GVHD) is characterised by lymphocytic infiltrates, crypt hyperplasia, and villous atrophy. It has been shown that programmed cell death (apoptosis) of the crypt epithelium takes place during the intestinal manifestation of acute GVHD.
AIMS To investigate which of the two most investigated inductors of apoptosis (Fas ligand (FasL) and tumour necrosis factor α (TNF-α)) is responsible for the induction of apoptosis in this animal model.
METHODS Animals undergoing acute semiallogeneic GvH reaction were treated with neutralising anti-TNF-α, two different anti-FasL antibodies, or pentoxifylline.
RESULTS Anti-TNF-α application inhibited the appearance of apoptotic cells in the intestinal mucosa, whereas anti-FasL antibodies had no influence on mucosal apoptosis. In addition, the transfer of FasL deficient (gld) donor lymphocytes still induced crypt cell apoptosis, villous atrophy, and crypt hyperplasia. Furthermore, when the animals were treated with pentoxifylline, a known inhibitor of TNF-α secretion in vitro and in vivo, there was significant normalisation of the intestinal morphology accompanied by inhibition of epithelial apoptosis.
CONCLUSIONS The FasL-Fas interaction is not involved in the induction of apoptosis during acute GVHD. However, neutralisation of TNF-α by an antibody or by pentoxifylline inhibits the occurrence of apoptosis and of mucosal atrophy in this animal model. These results have implications for the treatment of immunologically mediated human atrophic gut diseases—for example, diet refractory cases of coeliac disease.
- graft versus host disease
- tumour necrosis factor
- Fas ligand
- pentoxifylline
- apoptosis
- crypt cell
Abbreviations used in this paper
- FasL
- Fas ligand
- GVHD
- graft versus host disease
- TNF
- tumour necrosis factor
- IFN
- interferon
- IL
- interleukin
- GvH
- graft versus host
Statistics from Altmetric.com
Apoptosis has been shown to be critically important for developmental processes,1 for control of neoplastic growth,2 and for homoeostasis of the immune system.3 The interaction of T cells with epithelial cells is of specific interest in this respect. In a previous study we showed that crypt epithelial cells die due to apoptosis during the manifestation of mucosal atrophy in the murine model of T cell mediated acute graft versus host disease (GVHD).4
Apoptosis has been shown to be induced by multiple soluble and membrane bound factors in numerous experimental systems.5 However, tumour necrosis factor (TNF) α and the Fas ligand (FasL)-Fas interaction are the two most investigated mediators of apoptosis. Furthermore, they have also been suggested to be effector mechanisms in the injury of the target organs (liver, skin, gastrointestinal tract) during an acute GvH reaction. Mowat and colleagues6 showed that application of recombinant TNF-α alone can induce typical gut injury—consisting of crypt hyperplasia and villous atrophy—and Piguetet al showed that neutralisation of TNF-α in murine acute GVHD leads to reduced target organ damage.7 Whether this damage is induced by direct TNF-α mediated apoptosis and which cell type is mainly affected, is not known. On the other hand, there are few data concerning the relevance of the Fas-FasL interaction in intestinal apoptosis during acute GvH. Recently, Sakai et al published a study in which they showed the capacity of freshly prepared intraepithelial lymphocytes of GvH animals to cause cytotoxicity in vitro via a FasL mediated mechanism.8 However, the in vivo relevance of the FasL-Fas interaction was not investigated in these studies.
Therefore, the different impacts of the blockade of the FasL-Fas interaction on the one hand versus neutralisation of TNF-α on the other, on the occurrence of intestinal crypt cell apoptosis in murine acute GVHD were investigated. As a second step, we studied the effect of pentoxifylline—a known inhibitor of TNF-α liberation in vitro and in vivo9—on the occurrence of apoptosis in the gut and on the development of crypt hyperplasia and villous atrophy.
Materials and methods
ANIMALS, CELL PREPARATIONS, AND INDUCTION OF ACUTE SEMIALLOGENEIC GvH DISEASE
Mouse strains C57/BL6, DBA2, and the F1 generation of these two strains (B6D2F1) were raised and kept under standard conditions in the animal facility of the University Hospital of Kiel, Germany.
To induce semiallogeneic GVHD a slightly modified procedure as described by Guy-Grand and Vassalli10 was performed. To prepare donor lymphocytes, spleen and mesenteric lymph nodes of C57/BL6 mice were removed and pressed through a cell filter (40 μm pore size). Red blood cells were subsequently lysed by a hypotonic lysing buffer (ACK buffer, Boehringer Ingelheim, Germany). The resulting lymphocytes (80 × 106 cells/animal) were transferred to 8–14 week old irradiated (7.5 Gy) B6D2F1 mice of the same sex by intraperitoneal injection. Three or six days after the induction of semiallogeneic GVHD recipient animals were sacrificed. Spleens, and inguinal and mesenteric lymph nodes, as well as the small bowel were removed and frozen in liquid nitrogen or fixed in 10% phosphate buffered formalin (pH 7.4) pending further analysis. Control animals consisted of irradiated B6D2F1 mice which received the same amount of syngeneic (B6D2F1) cells.
In a further subset of experiments lymphocytes of gld/gld (FasL deficient) mice on a C57BL/6 genetic background (Charles River, Sulzield, Germany) were prepared and transferred into B6D2F1 recipient mice as stated above. Mice were sacrificed on day 3 to study apoptosis and on days 3 and 6 to investigate mucosal atrophy.
TREATMENT REGIMENS OF RECIPIENT ANIMALS
After induction of GVHD one group of recipient B6D2F1 mice was treated with a neutralising anti-TNF-α antibody (Pharmingen, San Diego, California, USA; 200 μg/day intraperitoneally), a rabbit polyclonal anti-FasL antibody (200 μg/day intraperitoneally), a known neutralising monoclonal hamster anti-FasL antibody (MFL-3, Pharmingen), or pentoxifylline (Albert-Roussel, Wiesbaden, Germany; 100 mg/kg bodyweight twice a day, intraperitoneally). The animals were sacrificed at day 3 or day 6, respectively, and organs were removed as described above.
The polyclonal anti-FasL antibody was raised in rabbits after immunisation with a KLH linked peptide of the extracellular domain of the murine Fas-L molecule11 (aa 200–214: CNNQPLNHKVYMRNS). The antibody was affinity purified using peptide coated Sepharose (Pharmacia, Uppsala, Sweden) as described previously.12
The efficacy of the monoclonal hamster anti-FasL (MFL-3) antibody in blocking apoptosis in vitro has been shown by Kayagakiet al.13 To test the efficacy of rabbit polyclonal anti-FasL antibody, the following in vitro assay was performed: murine splenic T cells were prepared and stimulated on anti-CD3 coated six well plates for 48 hours. The supernatant was then removed and the cells transferred to a novel, anti-CD3 coated six well plate. Apoptosis under these conditions (interleukin 2 (IL-2) withdrawal during restimulation) is known to be caused by the FasL-Fas interaction.14 Some of the cells were incubated in the presence of polyclonal anti-FasL antibody (10 μg/ml). Six hours later cytospins of these cells were made. Apoptosis was detected using the ApopTag in situ apoptosis detection kit (Oncor, Gaithersburg, Pennsylvania, USA).
DETECTION OF MUCOSAL ATROPHY
Formalin fixed jejunum specimens were embedded in paraffin wax and cut into 6–8 μm sections. To detect mucosal atrophy and crypt hyperplasia slides were stained with haematoxylin and eosin and the villous height and crypt depth were measured using a graded ocular.
DETECTION OF APOPTOTIC CELLS
In order to determine the rate of apoptosis during acute GVHD, treated and untreated recipient animals were sacrificed at various time points (one to six days) after induction of GVHD; the jejunum was removed and snap frozen in liquid nitrogen. To detect apoptotic cells with the help of the TUNEL technique the ApopTag in situ apoptosis detection kit (Oncor) was used according to the manufacturer’s instructions.
MIXED LYMPHOCYTE REACTION
To investigate whether treatment with pentoxifylline leads to a generalised downregulation of the immune response of the C57BL/6 donor lymphocyte against the allogeneic stimulus or whether it only affects TNF-α mediated mechanisms, splenic lymphocytes of syngeneic control animals, GvH animals, and pentoxifylline treated GvH animals were prepared and cocultured with irradiated (3 Gy) DBA2 splenocytes. After 72 hours proliferation was assessed using 3H-thymidine as described elsewhere.15 Interferon γ (IFN-γ) was measured in the supernatants using a sandwich ELISA technique provided by Pharmingen.
Results
HISTOLOGICAL CHANGES IN THE JEJUNUM AFTER INDUCTION OF ACUTE GVHD
In accordance with earlier reports,10 ,16 two to three days after induction of GVHD the lamina propria and the mucosal epithelium began to be infiltrated by increased numbers of CD4+ and CD8+ T cells.4 At that time the villi began to appear atrophic and the crypt depths increased. The animals appeared grossly healthy until about one week after induction of acute GVHD. Later the animals developed diarrhoea, weight loss, and skin changes. Most of the animals died between days 14 and 21 after lymphocyte transfer. Histological changes in the jejunum were assessed on day 6, when the animals still appeared healthy.
Irradiated (7.5 Gy) control animals that received syngeneic lymphocytes did not show a significant change in mucosal architecture at day 6 compared with totally untreated animals (data not shown). This result is in accordance with earlier studies by Potten et al,17 who proved that irradiation at the dosage chosen does not lead to lasting damage of the intestinal mucosa.
RATE OF EPITHELIAL CELL APOPTOSIS
Previously, we have shown that villous atrophy in the GvH animals is preceded by programmed epithelial cell death (apoptosis),4 as has been speculated before.18 However, it is not known which of the two most investigated mediators of apoptosis (Fas and TNF-α) is mainly responsible for induction of apoptosis in this animal model. Therefore, we treated the animals undergoing acute GvH reaction with neutralising anti-TNF-α or two different anti-FasL antibodies, respectively, for three days and counted the number of apoptotic cells in the epithelial cell layer using the TUNEL technique at day 3 after induction of acute GVHD. As shown in fig 1, apoptosis is significantly inhibited by administration of neutralising anti-TNF-α antibody but not by the neutralising anti-FasL antibodies, indicating that the FasL-Fas system is probably not as important for induction of intestinal apoptosis in murine acute GvH as is TNF-α. Table 1 shows the exact numbers of apoptotic cells. Furthermore, treatment with anti-FasL antibodies did not inhibit mucosal transformation in the acute GvH reaction (data not shown).

Irradiated B6D2F1 mice received C57/BL6 lymphocytes (semiallogeneic). One group of GvH animals was not treated further (A; n=6), one group received different anti-FasL antibodies (B; n=5), and one group was treated with anti-TNF-α antibodies (C; n=5). Animals were sacrificed at day 3 and 6–8 μm cryosections of the jejunum were made; slides were stained with the ApopTag direct apoptosis in situ detection kit and counterstained with methyl green. Apoptosis is frequent in A and B, but totally abolished in C.
Number of apoptotic cells in each group
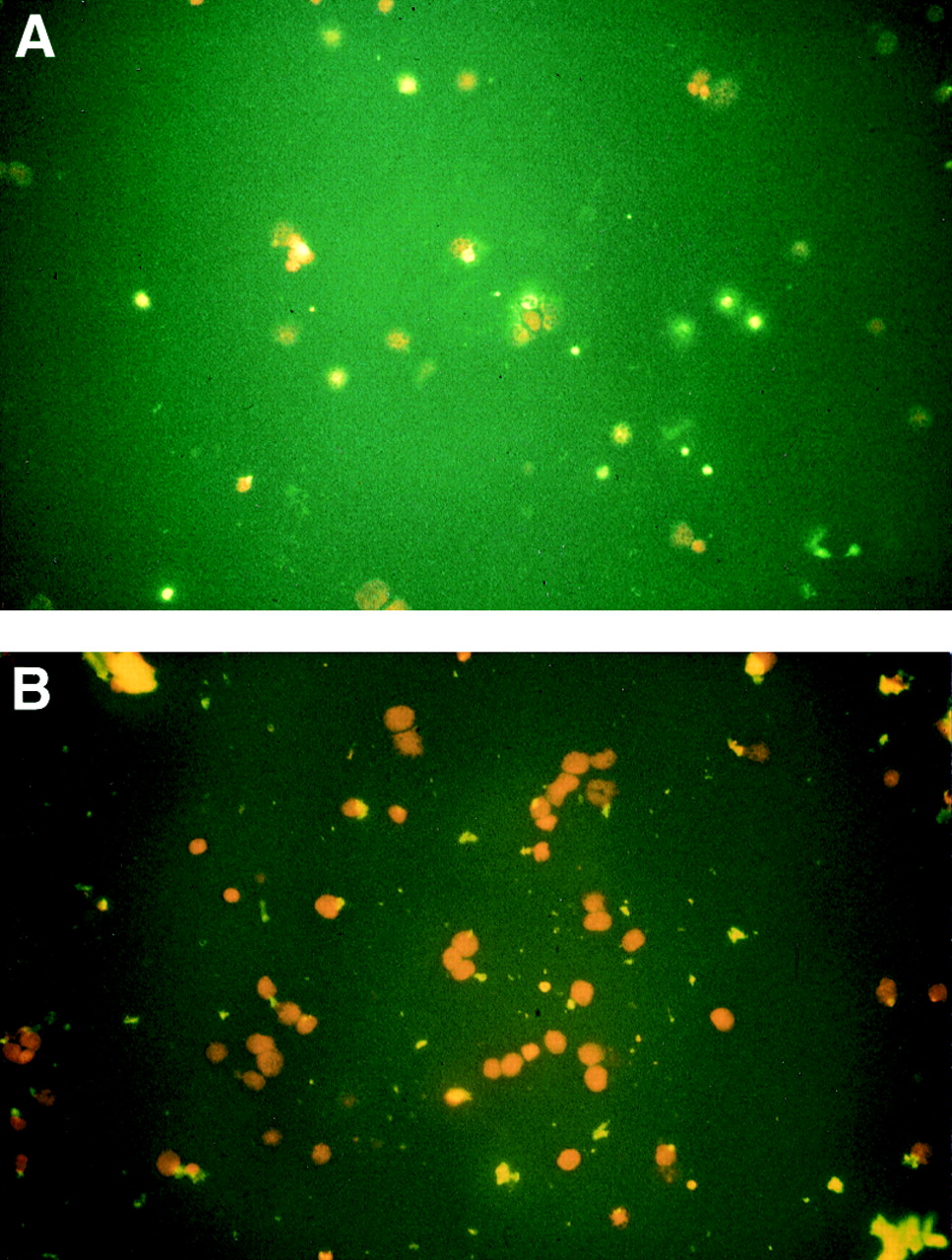
Figure 2 shows the efficacy of the polyclonal anti-FasL antibody in preventing apoptosis in vitro. In the presence of the polyclonal anti-FasL antibody approximately 5–10% of the visible cells underwent apoptosis; without this antibody 30–50% of the cells were apoptotic (fig2A).

(A) Cytospins of T cells without FasL antibody; (B) cytospins of T cells in the presence of polyclonal anti-FasL antibody. Slides were stained with the fluorescence ApopTag direct apoptosis in situ detection kit and counterstained with propidium iodide. Data represent results of two different experiments.
INDUCTION OF APOPTOSIS, VILLOUS ATROPHY, AND CRYPT HYPERPLASIA BY FasL DEFICIENT DONOR CELLS
As described by Takahashi et al,11 the lymphoproliferative disease observed in gld mice is caused by a point mutation in the FasL gene on chromosome 1, resulting in expression of a non-functional FasL protein. The expressed FasL does not induce apoptosis. Therefore, we tested our hypothesis that the FasL-Fas interaction might not be important for induction of crypt cell apoptosis in murine acute semiallogeneic GVHD by using donor lymphocytes of gld mice on a BL/6 genetic background. As shown in fig 3, the FasL deficient donor cells still induce apoptosis (24 (10) apoptotic cells per high power field) and profound villous atrophy and crypt hyperplasia (villous length 174 (12.5) μm, crypt depth 159 (14) μm, crypt/villous ratio 0.91, n=4). This finding thus corroborates results of studies using the two different anti-FasL antibodies as shown above.

Irradiated B6D2F1 mice received gld/gld lymphocytes and were sacrificed on day 3 to detect apoptotic cells using the TUNEL technique (A; n=4); paraffin wax sections of the jejunum were prepared and stained with haematoxylin and eosin (B; n=4). These photomicrographs clearly show ongoing apoptosis in the jejunal crypt epithelium and the manifestation of crypt hyperplasia and villous atrophy.
EFFECT OF PENTOXIFYLLINE ON APOPTOSIS, INTESTINAL MORPHOLOGY, AND ALLOGENEIC LYMPHOCYTE ACTIVATION
Other investigators7 have shown that treatment of mice undergoing acute GvH with anti-TNF-α leads to an inhibition of the development of mucosal atrophy, emphasising the important role of apoptosis in the intestinal manifestation of GVHD. Thus, we wondered whether pentoxifylline, a known inhibitor of TNF-α release in vivo and in vitro,9 would bring about normalisation of intestinal morphology. Therefore, we treated some animals after the induction of acute GvH with pentoxifylline. This treatment led to a profound inhibition of apoptosis (fig 4) and also of the degree of the lymphocytic infiltrate, crypt hyperplasia, and villous atrophy, as shown in fig 5. The ratio of crypt depth to villous length was almost normalised (from 0.88 in GvH animals to 0.37 in pentoxifylline treated animals). Syngeneic control animals had a crypt: villous ratio of 0.29 (table 2).

Irradiated B6D2F1 mice received C57/BL6 lymphocytes (semiallogeneic) or B6D2F1 lymphocytes (syngeneic control) (A; n=5). One group of GvH animals was not treated further (B; n=5) and the other group was treated with pentoxifylline (C; n=5). Animals were sacrificed at day 3, and 6–8 μm cryosections of the jejunum were made; slides were stained with the ApopTag direct apoptosis in situ detection kit and counterstained with methyl green. Apoptosis was significantly inhibited by pentoxifylline treatment. Results were similar for animals sacrificed on days 2, 4, and 5 (data not shown).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Irradiated B6D2F1 mice received C57/BL6 lymphocytes (semiallogeneic) or B6D2F1 lymphocytes (syngeneic control) (A; n=5). One group of GvH animals was not treated further (B; n=5) and the other group was treated with pentoxifylline (C; n=5). Animals were sacrificed at day 6, the jejunum was removed and fixed in 10% phosphate buffered formalin. After being embedded in paraffin wax, 6–8 μm sections were cut and the slides were stained with haematoxylin and eosin. The mucosal architecture was almost normalised after the application of pentoxifylline.
Effect of pentoxifylline (PF) treatment
When a mixed lymphocyte reaction was performed using in vivo activated lymphocytes of treated (pentoxifylline) and untreated GvH animals, no difference in the rate of proliferation on restimulation and in the secretion of IFN-γ could be found (table 3), indicating that complete allogeneic stimulation was not abolished by application of pentoxifylline.
Proliferation and interferon (IFN) γ secretion
Discussion
Murine acute GVHD has long been investigated for two reasons: as an animal model for the immune pathomechanisms of human GVHD after allogeneic bone marrow transplantation. Furthermore, due to its characteristic changes in the intestinal histology consisting of lymphocytic infiltrates, crypt hyperplasia, and villous atrophy, it serves as a model for immunologically mediated atrophic-hyperregenerative diseases of the intestine (for example, coeliac disease, Whipple’s disease, bacterial overgrowth).19 The systemic cytokine profile associated with the above mentioned changes in the intestinal architecture has been determined to be dominated by a Th1 pattern (predominance of IFN-γ secretion).20 Neutralising antibodies to IFN-γ and to IL-12, which governs Th1 differentiation,21 can abolish the systemic manifestation of acute murine GvH.22-24 Inflammatory cyokines such as TNF-α were also shown to be involved in development of these intestinal alterations in this animal model.7 These cytokine studies are paralleled by investigations of human biopsy material of patients with coeliac disease, which show similar cytokine expression patterns to those seen in murine acute GVHD.25-27 How the investigated cytokines regulate epithelial growth, damage, and regeneration is still a matter of debate. However, it has been shown that TNF-α and IFN-γ can induce upregulation of MHC class II molecules in epithelial cells and thereby indirectly render them a better target for cytotoxic T cells.19 Furthermore, it has also been speculated that these cytokines exert directly damaging effects on the gastrointestinal mucosa.6 ,7 ,28 In a previous study we showed that apoptosis of crypt epithelial cells occurs during the process of mucosal transformation in acute murine GvH.4 Again, this finding is paralleled in a study by Moss et al,29 who found increased epithelial cell apoptosis in duodenal biopsy specimens of patients with coeliac disease. In contrast to our studies, they showed enhanced apoptosis in the villous tip area and not in the crypt epithelium. Furthermore, a recent study by Guy-Grand et al showed increased villous cell apoptosis in TNF-α treated normal animals,30 thus corroborating the importance of TNF-α in intestinal apoptosis, as was found in the present study. However, the systemic application of high concentrations of this cytokine does not mimic any known pathophysiological condition (neither GvH reaction nor coeliac disease). Therefore, the difference in the location in which apoptosis occurs might result from the chosen experimental setting.
In the present study we investigated the mechanism of the induction of epithelial cell apoptosis in acute murine GVHD. We focused on the two most investigated mediators of apoptosis: TNF-α and the FasL-Fas interaction. Using neutralising antibodies, we found that TNF-α—either directly or indirectly—is most likely responsible for the induction of apoptosis in the acute GvH reaction. The latter finding is corroborated by studies using gld (FasL deficient) mice, showing that even in the absence of a functional FasL-Fas interaction apoptosis, crypt hyperplasia, and villous atrophy do occur in this animal model. However, these results are in contrast to the conclusion of the study by Sakai et al, in which they showed the capacity of freshly prepared intraepithelial lymphocytes of GvH animals to cause cytotoxicity in vitro via a FasL mediated mechanism.8 However, in their experimental setting the recipient mice were not irradiated. Unirradiated mice develop only insignificant mucosal atrophy (personal observation). Therefore, the in vivo relevance of the FasL-Fas interaction could not be investigated. With reference to our data using the neutralising anti-FasL antibodies we would speculate that the FasL-Fas interaction might have only a minor effect on the manifestation of epithelial apoptosis and villous atrophy in vivo. It is possible that other newly discovered factors—such as TRAIL31 and TRAMP32—may also influence this process. This should be investigated in future studies.
The source of TNF-α in acute GvH is not known. It has been shown that T cells and macrophages produce increased amounts of this inflammatory cytokine, especially in later phases of established GVHD.20 ,33 Surprisingly, it has been shown by in situ hybridisation techniques that in normal human and murine intestine Paneth cells are the major source of TNF-α.34 ,35 Thus, it is possible that in the initial phase of murine acute GvH the Paneth cells are either destroyed and thereby release TNF-α or are somehow stimulated to secrete TNF-α. Later, T cells and macrophages may be the source of this cytokine. This hypothesis may also explain the above stated difference in localisation of the apoptotic cells in the mouse model and in human coeliac disease. To test this theory, we are currently investigating the effects of the manifestation of acute GvH on Paneth cells.
Having found that TNF-α is crucial for crypt cell apoptosis in murine GVHD, we wanted to test the efficacy of the phosphodiesterase inhibitor pentoxifylline, a clinically well known substance that inhibits TNF-α secretion by many cell types both in vitro and in vivo.9We found that pentoxifylline—although it does not completely downregulate allogeneic T cell stimulation—abolished TNF mediated apoptosis of the crypt cells and reduced the occurrence of crypt hyperplasia and villous atrophy after induction of the acute GvH reaction. Thus, it is possible that patients with immune mediated villous atrophy and crypt hyperplasia, that do not respond to standard treatments, could benefit from treatment with pentoxifylline, a substance with few side effects.9 This treatment would be more cost efficient compared with treatment with an antibody against human TNF-α, which should theoretically also be effective in reducing mucosal atrophy. In addition, pentoxifylline has been shown to prevent experimental encephalomyelitis in rats by selectively suppressing Th1 cytokines,36 ,37 thus making this substance even more interesting for the treatment of immune mediated diseases.
The molecular mechanism of pentoxifylline has been shown by a recent study which identified its selective inhibitory effect on the transcription factor c-Rel in activated T cells.38 In contrast to cyclosporin A and FK506 other transcription factors such as NF-AT are not influenced, making pentoxifylline much more selective than these two immunosuppressants. In in vitro experiments39 and also in animal and human studies, the striking inhibitory effect on TNF-α, but not IL-6 production in various models of endotoxaemia induced TNF release was often shown.40 However, when pentoxifylline was tested outside a laboratory setting—for example, in the treatment of GvH after bone marrow transplantation or of Gram negative sepsis, two diseases in which TNF-α is thought to play a crucial, fatal role, the outcome was rather disappointing.41 ,42 Possibly, the effect of pentoxifylline is too selective and does not influence other unknown pathomechanisms besides TNF-α. Therefore, its clinical impact in treatment of diseases other than claudicatio intermittens is limited.
In conclusion, we have shown that TNF-α—and not the FasL-Fas interaction—is (directly or indirectly) mainly responsible for induction of epithelial apoptosis in the acute murine GvH reaction. Treatment with pentoxifylline, a known inhibitor of TNF-α production, results in a decreased rate of crypt cell apoptosis and inhibition of the occurrence of crypt hyperplasia and villous atrophy in this animal model, suggesting a possible clinical application of this substance in patients with immune mediated villous atrophy and crypt hyperplasia.
Acknowledgments
We acknowledge the superb technical assistance of Maren Dirks. The study was supported by a grant of the Deutsche Forschungsgemeinschaft (STU 157/3–1).
Abbreviations used in this paper
- FasL
- Fas ligand
- GVHD
- graft versus host disease
- TNF
- tumour necrosis factor
- IFN
- interferon
- IL
- interleukin
- GvH
- graft versus host