Article Text

Abstract
BACKGROUND Paracellular permeability to solutes across the descending colon is much higher in cattle than sheep. This is a possible route for transmission of infective materials, such as scrapie prion.
AIMS To compare the permeabilities of labelled scrapie prion protein and other macromolecules in bovine and ovine descending colons in vitro.
METHODS Using fresh slaughterhouse material, transepithelial fluxes of macromolecules across colonic mucosae mounted in Ussing chambers were measured by monitoring transport of either enzyme activity or radioactivity.
RESULTS The comparative bovine to ovine permeability ratio of the probes increased with molecular weight: from 3.1 (0.13) for PEG400 to 10.67 (0.20) (p<0.001) for PEG4000; and from 1.64 (0.17) for microperoxidase to 7.03 (0.20) (p<0.001) for horseradish peroxidase (HRP). The permeability of125I-labelled inactivated Syrian hamster scrapie prion protein (ShaPrPsc) was 7.02 (0.33)-fold higher in bovine than ovine colon (p<0.0025). In each species, the probe permeabilities decreased according to the formula: P = Po.exp(−K.ra). The “ideal” permeabilities, Po are similar, however, K(ovine) = 2.46 (0.20) cm/h/nm exceeds K(bovine) = 0.85 (0.15) cm/h/nm (p<0.001) indicating that bovine colon has a higher proportion of wide pores than ovine. Image analysis confirmed that HRP permeated through the bovine mucosal layer via a pericryptal paracellular route much more rapidly than in sheep.
CONCLUSIONS These data may imply that scrapie prion is transmitted in vivo more easily across the low resistance bovine colonic barrier than in other species.
- cattle
- sheep
- colon
- paracellular permeability
- horseradish peroxidase
- hamster scrapie prion protein
Abbreviations used in this paper
- BSE
- bovine spongiform encephalitis
- DAB
- 3,3′-diamino-benzidine tetrahydrochloride
- HRP
- horseradish peroxidase
- MBM
- meat and bone meal
- MS
- mucosal to serosal
- PEG
- polyethylene glycol
- PrPsc
- scrapie prion protein
- ShaPrPsc
- Syrian hamster scrapie prion protein
- SM
- serosal to mucosal
Statistics from Altmetric.com
Transfer of proteins and polymer antigens across the intestinal mucosal barrier occurs to a variable extent. Most proteins crossing via the transcellular route are degraded by intracellular proteases.1-9 This results in the loss of enzymatic and antigenic activity. However, in some cases the absorbed protein emerges intact and immunologically active. It is conventionally assumed that absorption of intact proteins implies absorption via a paracellular route, which avoids contact with intracellular proteases. However, this view has been disputed as it has been shown that peptides or horseradish peroxidase (HRP) with antigenic potential can remain intact after transcytosis across monolayers of HT29–19A, a human colonic epithelial cell line.1 Transcytosis of intact HRP may be enhanced by raising lysosomal pH either withHelicobacter pylori or bafilomycin A1 which increase ammonia in lysosomes.10 Mucosal to serosal flux of HRP is increased across rat jejunum in vitro via paracellular and endocytotic transcellular routes by carbachol.4Horseradish peroxidase (molecular weight 40 kDa) can cross the jejunal mucosa with relative ease via the paracellular route.6
The colon is considered to be relatively impermeable to macromolecules—for example, polyethylene glycols (PEG) of various sizes have been used as indicators of paracellular flow across the intestine.11-15 This permeation greatly increases during stress and inflammatory states in all parts of the intestine, including the colon.6 ,7 ,11 ,15
The consensus view is that macromolecular transport can occur, albeit to a small extent, via both paracellular and transcellular routes in all parts of the intestinal tract, and to a lesser extent towards the distal end. Inflammatory processes and possibly neurotransmitter stimuli may also increase uptake via both transcellular and paracellular routes.
Although sheep and cattle have similar diets and habitats, sheep produce pelleted faeces with a low water content (65–70% wt/vol), whereas cattle faeces are soft and unformed with a water content of more than 95% wt/vol.16-18 Consolidation of faeces requires a very large force, about 4–10 atmospheres.19 ,20 This pressure is generated by the osmotic gradient produced by accumulation of NaCl in the pericryptal spaces of descending colonic crypts. The osmotic pressure draws fluid across the crypt wall out of the crypt lumen. The ensuing low pressure in the crypt lumen draws fluid out of the faeces in the colonic lumen.21-24 As bovine descending colon has a high passive permeability to NaCl, a large osmotic pressure gradient cannot be sustained across the crypt wall.20 Thus cattle are unable to generate hard faeces. This property may have survival value, as liquid faeces can readily be recycled along the entire length of the large intestine, thereby prolonging the fermentation time and raising the efficiency of energy extraction from feed largely composed of cellulose.
Another important consequence of the high paracellular permeability of cattle colon may be that they are more susceptible to antigen uptake than animals that produce consolidated faeces. Scrapie prion protein (PrPsc) is very resistant to all forms of physical and chemical assault. It resists proteolytic digestion from proteinase K and it is insoluble in high concentrations of detergents.25 ,26 It also retains its infectivity when buried in soil for three years or more, and resists all but the most severe methods of disinfection.27-29 Because of its high resistance to digestive enzymes in the small intestine and fermentation processes in the foregut and hindgut, it remains biologically active within the gastrointestinal tract and faeces. As uptake of scrapie prion into brain is contingent on it crossing the gastrointestinal barrier, the resistance of this barrier to prion uptake may be an important means of reducing its eventual uptake into brain.
Scrapie prion entry into the systemic circulation is currently considered to be mainly by transcytosis via lymphoid tissue in the gastrointestinal tract, for example, via tonsillar glands and Peyer's patches.30-32 As the prion infectivity resists phagocytic degradation, this view may be correct in general. However, it does not provide a satisfactory explanation for the apparently higher rate of scrapie prion infectivity in cattle than sheep from meat and bone meal (MBM) feed.28 ,29
The aim of this paper is to test the hypothesis that the scrapie prion protein is more permeable in bovine than ovine colon, because bovine colon has a higher permeability than most other species to macromolecules in general.
To this end the rates and routes of passive movement of a number of macromolecules across isolated bovine and ovine colonic mucosa have been compared. The macromolecules were selected on the basis of having an easily monitored activity, either biological, or by radiological techniques, and being of similar size to scrapie prion protein. We have also compared the rates of macromolecular permeation with that of iodine-125 labelled hamster scrapie prion protein.
We used an inactivated solubilised form of the scrapie prion as this permitted us to investigate its permeability across colon without the need for high security biological containment. Furthermore, use of high concentrations of detergents was not required to keep the prion protein in solution. Although a convenient model to investigate intestinal permeability, it could be argued that it is an unrealistic one, as the scrapie prion naturally occurs in multimeric fibrillar aggregates. Although this view has some force it has been shown that the infective nucleus of the prion is less than 60 kDa.33 As the prion is subjected to processes, both during preparation prior to ingestion and during digestion, which could lead to fibrillar disaggregation, it is likely that at least some of the prion exists in gut lumen in its monomeric form (28–30 kDa). Hence, it likely that the present model is a plausible, albeit imperfect, way of studying the immediate routes of bovine spongiform encephalitis (BSE) transmission in vivo.
Methods
PREPARATION OF TISSUES FOR PERMEABILITY MEASUREMENTS
Short lengths of bovine and ovine descending colons (0.5 m) were obtained from a local slaughterhouse (Bedford Slaughterhouse, Bedfordshire, UK) immediately after slaughter. The colons were washed free of faeces with cold Tyrode's solution (Na+ 150 mM, Cl− 120 mM, K+ 5 mM, Ca2+ 0.6 mM, NaH2PO4 0.5 mM, HCO3 12 mM, Mg2+ 0.83 mM, and glucose 5.6 mM) on site and quickly transported on ice to King's College. The mucosa was stripped from the underlying circular and longitudinal smooth muscle layers as described previously.20 The stripped mucosae were mounted in Ussing type chambers (six ports in parallel per chamber) which were maintained at 35°C or 20°C by an integral temperature controlled water supply. As gassing of the highly radioactive “hot” sides was considered both hazardous and a source of possible cross contamination into the “cold” solutions, it was avoided. Only the cold sides were gassed with 95% O2:5% CO2. Hot side solutions were fully gassed prior to addition of the labelled materials. The bathing solution consisted of Tyrode's solution. The slightly lower temperature (35°C) than normal was used to prevent tissue damage from hypoxia. Although probably an unnecessary precaution, it was taken to avoid gassing the hot side in order to prevent foaming and release of any potentially toxic material into the atmosphere. The tissue permeability to HRP was stable over a two hour incubation period using this protocol (see fig 2).

Effect of temperature on horseradish peroxidase (HRP) flux across bovine and ovine colon. Values are means (SEM); n=6 for bovine, n=8 for ovine from two animals.
PEROXIDASE PERMEABILITY
Horseradish peroxidase (type VI, molecular weight 40 kDa) or microperoxidase (molecular weight 1800 kDa; Sigma Chemical Company, Poole, Dorset, UK) in Tyrode's solution were freshly made up prior to flux experiments. The flux measurements were made using Ussing chambers as described previously.20 The enzyme marker was added to either the serosal or mucosal compartment, depending on whether serosal to mucosal (SM) or mucosal to serosal (MS) flux was being determined. The final concentration of HRP or microperoxidase in the labelled hot compartment was generally 100 μM. Samples from the unlabelled compartment were taken at 30 minute intervals for two hours. When the tissues were prepared for histology 200 μM HRP was used in the serosal solution.
ASSAY FOR HRP ACTIVITY
Samples (0.3 ml) from the cold side were added to 2.7 ml “Start solution” (50 ml 0.2 M NaH2PO4, 1 ml 0.2 M Na2HPO4, 20.4 μl 30% H2O2, 1.7 ml 1% dianisidine, made up to 204 ml with water). After 10 minutes the reaction was terminated by addition of 120 μl 4% sodium azide and the absorbance read at 460 nm. The relation between peroxidase concentration and absorbance is linear in the concentration range 10–100 pM.34
The HRP and microperoxidase fluxes and permeabilities were calculated using the following formulae: 
 , where JHRP = HRP flux (moles/h/cm), KHRP = HRP permeability, HRP2 = HRP concentration in the cold solution; HRP1 = HRP concentration in the hot solution, A = tissue area exposed (cm2), and Δt = the time increment (h) between t1 and t2.
, where JHRP = HRP flux (moles/h/cm), KHRP = HRP permeability, HRP2 = HRP concentration in the cold solution; HRP1 = HRP concentration in the hot solution, A = tissue area exposed (cm2), and Δt = the time increment (h) between t1 and t2.
PEG400 AND PEG4000 PERMEABILITY
Polyethylene glycol 4000 (molecular weight 4000) and PEG400 (molecular weight 400) were labelled with 3H, obtained from New England Nuclear and Amersham International; 0.1 μCi/ml was initially present in each hot side chamber. The radioactivity of these solutes was estimated using a liquid scintillation counter.
LIGHT AND ELECTRON MICROSCOPY
In order to observe exogenous peroxidase in bovine and ovine mucosa, strips of tissue were mounted in flux chambers, as described previously.20 HRP (type VI) was added to either the serosal or mucosal compartment (8 mg/ml, 200 μM) for 90–120 minutes. The mucosal strips were then fixed in situ for two hours by addition of 5 ml of 2.5% glutaraldehyde solution to both the mucosal and serosal compartments of the flux chambers. Following fixation, the tissues were stored in 0.1 M phosphate buffer at 4°C. The mucosal strips were incubated in 3,3′-diamino-benzidine tetrahydrochloride (DAB, Sigma Chemicals) containing medium (50 mg DAB per 100 ml of 0.1 M phosphate buffer plus 0.05% cobalt chloride at pH 6.0) and placed on a shaker rack for 10 minutes. The HRP was then visualised within the tissues by addition of 10 μl of 1% H2O2 per ml of DAB solution. The reaction was stopped after 10 minutes. Tissues were rinsed three times for one minute in ice cold 0.1 M phosphate buffer at pH 6.0 and then rinsed twice more in 0.1 M phosphate buffer at pH 7.4.34
For light microscopy tissues were dehydrated in graded alcohols, and incubated briefly in xylene before being embedded in wax. Serial lateral and transverse sections of 10 μm thickness were cut and stained with haematoxylin and permanently mounted on glass slides for photography. HRP was omitted from controls in order to monitor endogenous peroxidase activity. Peroxidase activity was localised in lateral and transverse sections of 10 μm thickness.
For electron microscopy, tissues were postfixed with osmium, dehydrated in graded alcohols, and embedded in EPON 812. Sections, 1 μm in thickness, were stained with uranyl acetate.34 The tissues were viewed with a Jeol 200CX electron microscope at 80 kV.
GEL PREPARATION AND ELECTROPHORESIS
Samples were run on 5% sodium dodecyl sulphate (SDS) gels, prepared according to the method of Laemmli.35 The tank buffer for electrophoresis contained 0.1% SDS, 27 mM Tris base, and 192 mM glycine, pH 8.3. Proteins bands were visualised using Coomassie stain.
LABELLING HRP WITH IODOGEN
Iodogen was purchased from Sigma Chemicals; 1 mCi 125I (Amersham International) was added to each tube in 1 ml of phosphate buffered saline (PBS) containing the protein to be radioiodinated at a concentration of 1 mg/ml. This solid phase method of radioiodination of proteins gives high yields of product material.36 The reaction was stopped after 30 minutes by separating the HRP from the low molecular weight materials in a G50 (Sephadex) column. The eluted labelled HRP was collected and used within 48 hours for flux measurements.
Preliminary experiments showed a large discrepancy between flux monitored by radioactive HRP and flux monitored by HRP activity. This difference was due to release of 125I in low molecular weight fragments from the HRP. Thus, the true flux of macromolecules could only be determined after separating the low molecular weight material from labelled HRP. This separation was performed with Ultrafree MC filters (UFC3 LGC regenerated cellulose membranes, Millipore), which reflect molecules above 10 kDa.
Breakdown of 125I-HRP into low molecular weight fragments was confirmed by SDS polyacrylamide gel electrophoresis (PAGE) analysis of the 125I-labelled HRP (fig 1). It is evident that there is a large dispersion of proteinaceous material in this unfiltered material (lane B). However, when this material was filtered through a Millipore MC centrifugal filter only a single band was evident (lane C). This filtered material had the same mobility as that of filtered material collected from the cold side (lane A) and it also had the same mobility as pure unlabelled HRP sample (lane E). These findings explain why the permeability ratio between bovine and ovine colon of the crude125I-labelled HRP is less than that of HRP activity.
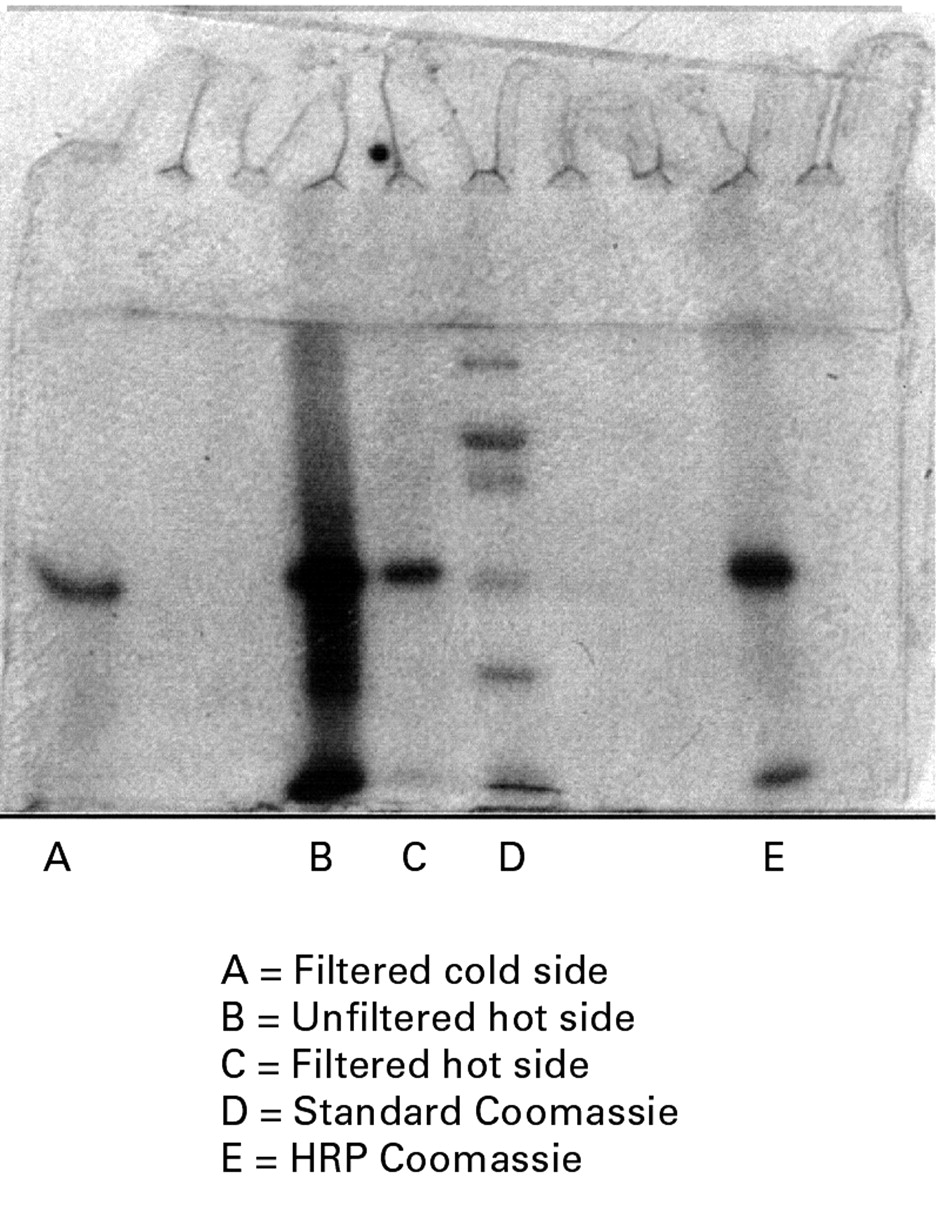
SDS-PAGE showing horseradish peroxidase (HRP) stained with Coomassie stain. Lane A, single band of 125I-HRP obtained from an ultrafiltered residue from the cold side; lane B, the unfiltered hot side of 125I-HRP; lane C, the filtered residue from the hot side of 125I-HRP; lane D, the molecular weight standards; lane E, pure unlabelled HRP.
LABELLING THE HAMSTER PrPSCWITH 125I IN IODOGEN
The purified and biologically inactivated hamster prion protein was initially obtained as guanidinium HCl inactivated fibres (gift from Dr James Hope, BBSRC Animal Health Unit, Edinburgh, UK). This material was added to the fresh iodogen tubes and incubated with 1 mCi125I for three hours. The labelled scrapie prion protein fibres were solubilised in 6 M guanidinium HCl and diluted into 5 ml of Tyrode's solution; 10 μl aliquots were added to the hot sides of the Ussing chambers (final concentration of labelled prion protein in hot side 1 μg/ml) together with HRP (100 μM).
Fluxes of unlabelled HRP activity and 125I-labelled hamster PrPsc protein were monitored simultaneously. The permeability of enzyme activity was determined by the peroxidase activity increase with time in the cold chamber and the permeability of PrPsc from the increase in 125I bound to protein. Each of the samples was filtered through the Millipore Ultrafree MC filters and washed twice in PBS before counting. The flux of radioactive label was monitored at 30 minute intervals as described above.
GEL ELECTROPHORESIS OF HRP IN THE HOT AND COLD SIDES OF THE TISSUE
Samples of 125I-labelled HRP were added to SDS lysis solution which consisted of 4 M urea, 1% SDS. Protein concentrations were estimated using the BCA assay (Pierce Chemical Co.) in which 10 μm of the homogenate was added to 3 ml of the assay solution. One lane contained 10 μl of a 25:75 mixture of high molecular weight Rainbow markers (high range 14.3–200 kDa; Amersham).
IMAGE ANALYSIS OF HRP PARTICLES IN MUCUS LAYERS
The numbers and densities of DAB stained HRP particles in the mucosal mucus layer were counted as follows: microscopic images of lateral sections through the mucus layer were examined using NIH image 1.62 (Wayne Rasband, NIH, Bethesda, Maryland, USA). The particle numbers were counted using the particle analysis macro. The total particle densities in each field were obtained, then the particle densities in the regions above the crypts were obtained by outlining regions of interest around the crypts. The number of particles within these regions was obtained using the particle counter as before. The particle densities in the pericryptal regions were obtained by subtracting the total number of particles subtending crypts from the total number of particles in the field and dividing by the pericryptal area. Four fields from each tissue were examined to obtain the data in table 2.
Analysis of staining particles in mucosal mucus after one hour of exposure to horseradish peroxidase (HRP; 200 μM) in serosal solution
SAFETY PRECAUTIONS
In this paper the permeabilities of ovine and bovine colonic mucosa, obtained from freshly slaughtered animals, to HRP and inactivated hamster scrapie prion protein are described. Precautions were taken against the possible biohazard from infected offal and hamster (although fully inactivated with 6 M guanidinium HCl) prion protein.
STATISTICS
Statistical analysis of the data was performed using Student'st test; only the two tailed probabilities are quoted.
Results
EFFECTS OF TEMPERATURE ON PERMEABILITY OF BOVINE AND OVINE DESCENDING COLONIC MUCOSA TO HRP
Horseradish peroxidase flux was linear after the first 30 minutes of incubation across both bovine and ovine colonic mucosa at both 35°C and 20°C (see fig 2; the linear regression lines are through the data points from 30 to 120 minutes). After an initial lag phase of 20–30 minutes HRP flux was linear. HRP flux was faster across bovine colon than ovine colon both at 35°C and 20°C. The effect of a temperature shift of 15°C was also much greater in bovine colon than ovine colon. HRP flux increased in bovine colon by 3.7 (0.17)-fold; with ovine colon the flux increased only by 1.16 (0.05)-fold (p<0.001). As both paracellular and transcellular fluxes were sensitive to changes in temperature,37 it is not possible to make any inferences about the routes of HRP movements across the colonic mucosa of either species. Both mucosal to serosal and serosal to mucosal fluxes were measured, but no significant differences between the unidirectional fluxes in either direction were found. Only mucosal to serosal fluxes are shown.
PERMEABILITY OF BOVINE AND OVINE DESCENDING COLONIC MUCOSA TO HRP, MICROPEROXIDASE, PEG400, AND PEG4000 AT 35°C
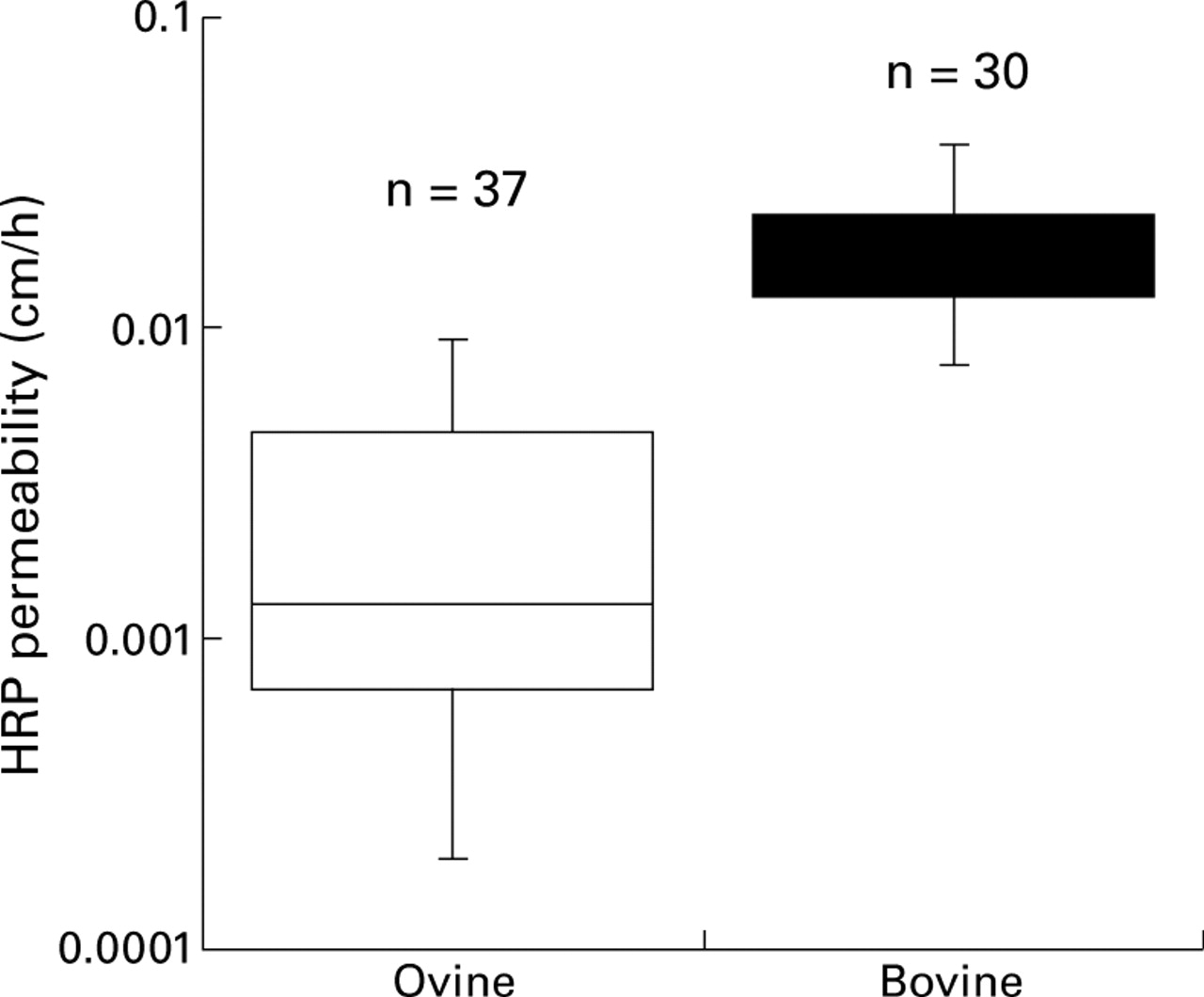
HRP flux across cattle and sheep colon was not normally distributed. The variance was consistent with a log normal distribution. The modal permeability was 0.0013 cm/h, and the mean permeability was 0.0027 (0.0005) cm/h. A box plot of the logarithmically transformed data shows that HRP permeability of bovine colon had a modal value approximately 14.5-fold higher than that of sheep (fig 3). These findings are consistent with the presence of a wider shunt pathway across bovine colon compared with ovine, and with previous findings.20 It would seem from the HRP flux data that small increments in the paracellular shunt size give exponential increases in HRP permeability. Several groups have also found that protein fluxes across small intestine have a log normal distribution.37 ,38 It is evident that bovine colon has a much higher permeability to hydrophilic solutes in general than ovine colon; as the solute size increases the disparity between bovine and ovine permeability increases. We observed that the permeability across bovine colon at 35°C was approximately 10–20-fold higher (figs 2 and3) than has been reported for HRP flux across rat jejunum.6

Box plots of permeabilities of horseradish peroxidase (HRP) activity in ovine and bovine colon in vitro. Each box encloses 50% of the data; the lines below and above show the minimal and maximal values within the range. The horizontal line is the modal value.
Permeability of 125I-labelled HRP (the macromolecular moiety) was monitored after filtering out the low molecular weight contaminants and found to be similar to that of HRP activity (unpublished data). The permeability of bovine colon to isotopically labelled PEG4000 was also approximately 10 times higher (p<0.001) than in ovine colon, which is significantly higher than the ratio of permeabilities of labelled PEG400 (table 1).
Comparison of permeabilities of various probes across isolated bovine and ovine descending colon
PERMEABILITY OF 125I-LABELLED HAMSTER SCRAPIE PRION PROTEIN
It is possible to separate 125I-labelled HRP from low molecular weight residues using Ultrafree Millipore centrifugal filtration. This method was used to monitor 125I-labelled PrPsc protein permeability across bovine and ovine colon. Table 1 shows the results.
The labelled, inactivated, scrapie prion protein has a similar permeability to HRP activity in both bovine and ovine colons. The scrapie prion protein is about sevenfold more permeable across bovine than ovine colon (p<0.001). These results indicate that the labelled prion protein permeability is of similar magnitude to any macromolecule in this molecular weight range (28 kDa) and indicates that it passes across the mucosa mainly via the paracellular route.
COMPARISON OF SOLUTE PERMEABILITIES BETWEEN OVINE AND BOVINE COLON
As has been observed previously in rat intestine, the size range of permeable solutes far exceeds the width of pore that is conventionally considered present in the intestine (0.8–1.2 nm).38-40 Over a very large dispersion of solute sizes, a plot of solute permeabilities (cm/h) versus the radius of the hydrated solute has been shown to fit the relation:  , where ra is the hydrated radius of the probe.41-43 The larger value of K, the greater the size discrimination of pores against smaller probes; hence a higher value of K reflects a higher relative proportion of small width to large width pores. P0 (cm/h) is the parameter which reflects the theoretical solute permeability, unhindered by frictional interactions with the pore wall = flux (moles/cm2/h)/concentration (moles/cm3); c is a constant reflecting leakage due to edge damage and possible cross contamination. This is only of consequence when observing solute fluxes. No significant difference was apparent between P0 bovine and ovine colons (fig 4). This implies that the total pore areas in ovine and bovine colon are similar. As K(ovine) is larger than K(bovine) by a factor of 2.9 (0.57) (p<0.001), this indicates that there is a much more discrimination against large molecules by ovine colon than bovine colon. It may be inferred that bovine colon has approximately 10 times more large pores in the radius range 2–4 nm than ovine colon. This is consistent with our previous finding that the electrical conductance of bovine colon (62.5 (5.4) mS) exceeds that of ovine colon (35.7 (3.06) mS) only by a factor of two (p<0.001).20
, where ra is the hydrated radius of the probe.41-43 The larger value of K, the greater the size discrimination of pores against smaller probes; hence a higher value of K reflects a higher relative proportion of small width to large width pores. P0 (cm/h) is the parameter which reflects the theoretical solute permeability, unhindered by frictional interactions with the pore wall = flux (moles/cm2/h)/concentration (moles/cm3); c is a constant reflecting leakage due to edge damage and possible cross contamination. This is only of consequence when observing solute fluxes. No significant difference was apparent between P0 bovine and ovine colons (fig 4). This implies that the total pore areas in ovine and bovine colon are similar. As K(ovine) is larger than K(bovine) by a factor of 2.9 (0.57) (p<0.001), this indicates that there is a much more discrimination against large molecules by ovine colon than bovine colon. It may be inferred that bovine colon has approximately 10 times more large pores in the radius range 2–4 nm than ovine colon. This is consistent with our previous finding that the electrical conductance of bovine colon (62.5 (5.4) mS) exceeds that of ovine colon (35.7 (3.06) mS) only by a factor of two (p<0.001).20

Semilogarithmic plot of the permeabilities (SEM) of the probes shown in table 1. The line is the best fit monoexponential line Px = Po.exp(−K.ra) + 0.002 fitted to the data weighted to the SEMs of the data. The equations fitting the lines have the following coefficients: bovine: Po = 0.35, K = 0.85 (0.15), regression coefficient (R) = 0.923; ovine: Po = 0.35, K = 2.46 (0.19), R = 0.952. The factor 0.002 represents a non-specific permeability coefficient, which may be due to edge damage; it only has significance with the low permeability ovine colon.
Electrolytes can pass through narrow channels with radii in the range 0.4–6 nm in colon. A small percentage difference in size of large pores can dramatically increase the permeability of macromolecules which are significantly hindered by frictional interactions with the pore wall, but will have an inconsequential effect on conductance of small molecules such as NaCl, which pass through large pores with negligible hindrance. Consequently, at best there can only be a poor correlation between macromolecular permeabilities and ionic conductance.4 ,6 ,20
HRP PERMEATION ACROSS BOVINE AND OVINE COLONIC MUCOSA
In bovine colon, HRP was taken up from the mucosal side into the lamina propria and accumulated on the inner surface of the muscularis mucosae (figs 5, 6, 7, and 8). In contrast, with ovine colon, only very slight HRP staining was detected along the abluminal border and no staining was evident within the lamina propria. This result suggests that the ovine mucosal border is a more effective barrier to HRP than bovine colon and it correlates with the higher transepithelial fluxes of HRP found in bovine colon.

Staining pattern of exogenous horseradish peroxidase (HRP) within ovine (A) and bovine (B) colonic mucosa. In control tissues there is no brown staining as this tissue was not exposed to HRP; mm, muscularis mucosae; M, mucosal surface; S. serosal surface. Stain: DAB plus haematoxylin. Original magnification × 25.

Staining pattern of exogenous horseradish peroxidase (HRP) within ovine (A) and bovine (B) colonic mucosa following exposure of the mucosal surface to HRP only. There is virtually no staining within the mucosal cells of the lamina propria of ovine colon; the lamina propria of bovine colon is heavily stained. M, mucosal surface, S, serosal surface. Stain: DAB and haematoxylin. Original magnification × 25.
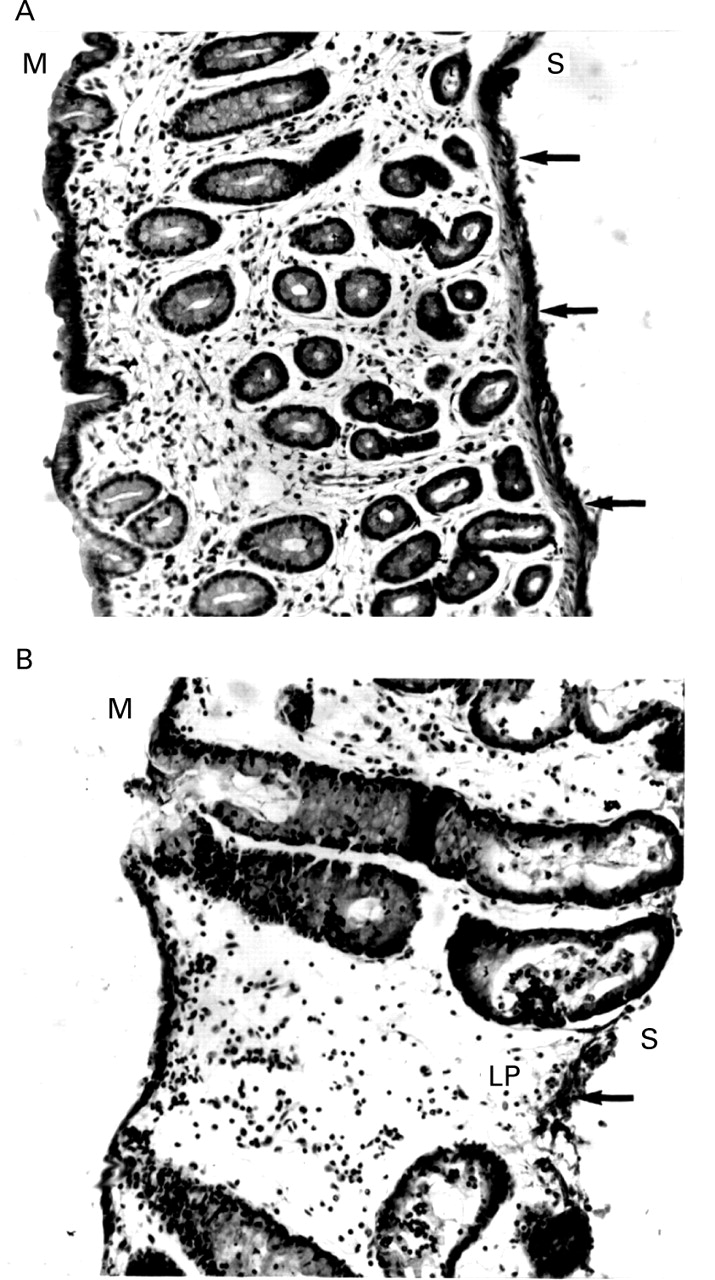
Staining pattern of exogenous horseradish peroxidase (HRP) within ovine (A) and bovine (B) colonic mucosa. The serosal side only is exposed to HRP. Note the accumulation of HRP stain indicated by arrows along the muscularis mucosae of ovine colon. In bovine colon there is more penetration of HRP from the serosa into the lamina propria (LP) compared with ovine. M, mucosal surface; S, serosal surface. Stain: DAB and haematoxylin. Original magnification × 16.

Lateral sections through the surface mucosa of bovine (A) and ovine (B) colon. The serosal side is exposed to horseradish peroxidase (HRP) only. Note that approximately six times more HRP staining is evident in the mucus layer above bovine colon than is present in ovine colon (see table 2). Stain: DAB and haematoxylin. Original magnification × 40.
When HRP was applied to the serosal side alone, the distribution of the HRP staining also differed notably in bovine and ovine colons (fig 7). A larger accumulation of HRP occurred along the serosal border of the ovine colon than in the bovine colon. This implies that the serosal layer in ovine colon provides a second barrier to diffusion of HRP. Additionally a larger amount of HRP stain accumulated below the mucosal surface of bovine colon than ovine. These findings indicate that HRP easily penetrates through the lamina propria of bovine colon. The intrinsic peroxidase activity of the tissues does not interfere with interpretation of the HRP fluxes as no significant DAB staining was observed in control tissue (fig 5).
IMAGE ANALYSIS OF THE HRP STAIN IN MUCOSAL MUCUS
Following exposure to serosal HRP alone for one hour the peroxidase penetrated through the mucosal layer and accumulated in the mucosal mucus in much larger amounts in bovine colon than in ovine colon (fig 8, table 2). The density of HRP particles in bovine mucus was almost sixfold greater than in ovine (p<0.001). This result is consistent with the data in table 1 and figs 2 and 3. Because HRP is fixed in the sticky mucus at its site of emergence on the mucosal surface, its distribution within the mucus layer is a good indicator of the region of tissue from whence it passed. Almost 70% of the particles in bovine mucus are assigned to a pericryptal route, whereas only 3% of the particles in ovine mucus can be assigned to the pericryptal route. However, the particle densities in the mucus above the crypts of both tissues were similar (bovine, 23.5 (1.4) and ovine 21.4 (1.2) particles per mm2 crypt area). This indicates that the rates of HRP movement via the transcellular and paracellular routes across the crypt walls of both species are similar.
The major difference between bovine and ovine colon is evidently the rate of HRP movement via the pericryptal (paracellular) route. The very low rate of HRP permeation via the pericryptal route in ovine colon in comparison to the transcryptal route suggests that the pericryptal route is normally sealed to HRP.
COMPARISON OF ELECTRON MICROGRAPHS OF BOVINE AND OVINE COLON
In bovine tissue only a thin fibroblastic layer was present within a thin basement membrane; thin and sparse collagen fibres were also present in the submucosal support tissue (fig 9). The myofibroblasts and collagen fibres in the basement membranes of ovine colon were both thicker and denser than in bovine colon. These findings corroborate the difference seen in the light microscopic appearance of bovine and ovine colon (figs 5, 6, and 7). Bovine colon has a more open, loose knit structure than ovine colon. These findings indicate that there is likely to be an anatomical basis for the wide difference in macromolecular permeability of the pericryptal paracellular route across the surface mucosa.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Electron micrograph of the serosal surface of bovine crypt luminal cells (A) and ovine crypt luminal cells (B). BM, basement membrane; CF, collagen fibres in cross section. Original magnification × 13 000.
Discussion
The results illustrate that bovine colon in vitro has a much higher permeability to macromolecules of 4 kDa and above than does ovine colon. The data in fig 4 show that this difference is due to a lower discrimination against larger molecules in bovine colon rather than to an overall increase in the number of pores. This suggests that the permeability of some paracellular pathway in bovine colon is higher.
Alternatively, it could be argued that the difference is due to higher rates of endocytosis in bovine than ovine colon. Higher rates of endocytosis have been reported in human colon from patients with Crohn's disease.7 Similarly it has been reported that there is some kind of size discrimination in transcytosis rates.1 ,10 Thus it can be claimed that neither size discrimination nor the absolute difference in rates of transport of macromolecules can be ascribed only to a difference in paracellular permeability. However, the data in table 2 and fig 8 show that transcytosis does not satisfactorily explain the difference in rates of macromolecular transport across bovine and ovine colon. A more likely explanation for the difference is the faster movement of solutes via the unrestricted bovine pericryptal pathway.
Histological comparison of the distributions of exogenous HRP within the bovine and ovine colon reveals that in vitro, ovine colon has two permeability barriers, whereas bovine colon has only the mucosal barrier. HRP is hindered from permeating between the serosal solution and the mucosal solution because access through ovine muscularis mucosa and lamina propria is restricted more than in bovine colon (figs 6 and7). These findings suggest that the submucosal tissues are more tightly held together by extracellular matrix and cellular adhesions in ovine than bovine colon. This view is corroborated by the electron micrographs (fig 9) showing that the myofibroblast layer, which is known to generate the collagen support matrix for the pericryptal sheath,44 ,45 is deficient in bovine colon.
This qualitative impression from histological appearances is supported by quantitative analysis of HRP penetration into the mucosal mucus layers. Approximately sixfold more HRP collects in the mucus layer above the mucosa of bovine colon after exposure to HRP in the serosal bathing solution than in ovine colon (fig 8, table 2). HRP staining patterns show that 70% diffuses via a pericryptal route in bovine colon and that paracryptal HRP flow is absent in ovine colon.
Collection of HRP in bovine mucus generates a honeycomb pattern surrounding bovine crypts. Each crypt is framed by annular deposition of DAB stain. Apart from the pericryptal insertion to the surface mucosa, the rates of HRP diffusion through the crypt wall either by transcytosis, or via the paracellular routes are similar in bovine and ovine colon.
These findings indicate that bovine descending colon is similar to rat caecum45 in that both have deficient myofibroblastic support matrices and neither tissue can generate hard faeces. The lack of a support matrix in bovine colon may explain the very high pericryptal permeability to macromolecules of the pericryptal route across the bovine surface mucosa. Differences in permeability and capacity to generate hard faeces exist between rat caecal and descending colonic mucosa for the same reason as between bovine and ovine descending colon; namely that rat caecal mucosa has a deficient myofibroblast support matrix compared with rat descending colon.44
PARACELLULAR TRANSPORT OF SCRAPIE PRION PROTEIN IN COLON
Bovine spongiform encephalitis (BSE) is generally considered to be caused by feeding cattle with MBM contaminated with scrapie agent from sheep and recycled cattle.28-30 ,46 The tests of transmissibility of BSE have depended mainly on bypassing the gastrointestinal barrier.31 ,47 Recent studies on transmissibility of BSE have concentrated on its genetic basis. These studies have shown that the species barrier to transmission may, in part, be due to the lower efficiency of conversion of the normal proteinase K sensitive form of the prion protein Pr(sens), to the abnormal proteinase K insensitive form, Pr(insen), which causes transmissible spongiform encephalopathy, PrPsc.48 ,49 Susceptibility to scrapie transmission depends on compatibility between the Prc and PrPsc forms of the prion protein; for example, 263K Syrian hamster strain, ShaPrPsc has a 60 day incubation in Syrian hamsters, but has not been successfully transmitted to mice.47 It has now been shown that this correlates with the low efficiency of conversion of Prc from 263K Syrian hamster strain to mouse PrPsc in vitro.
The presence of scrapie prion protein in lymphoid tissue associated with gut before the appearance of neurological lesions suggests that transcytosis via gastrointestinal lymphoid tissue may be an important route of uptake PrPsc from the gut lumen.32However, no such material has been reported in bovine lymphoid tissue.50
A recent paper by Bons and colleagues32 shows that lemurs fed with BSE infected bovine brain (0.5 g/100 g animal) had widespread dissemination of the PrPsc in the gastrointestinal tract (tonsils, oesophagus, stomach, large and small intestine), spleen, brain, and spinal cord six months after infection. The authors suggest that the presence of antibody in the gastrointestinal lymphoid tissue is good evidence that this is the primary route by which the infection enters the body. However, this view is based on data obtained six months after exposure. At this time the PrPsc has already spread into the central nervous system of these animals, so it cannot be assumed with confidence that the infection has not also spread back into the gastrointestinal lymphoid tissue after primary infection via some other route.
A tight gastrointestinal permeability barrier, as in sheep, may mean that the efficiency of transmission of PrPsc via the oral route is lower in these animals, as most of the ingested material is excreted in faeces. On the other hand, a high bovine permeability to PrPsc may permit entry of a much greater amount of the ingested Pr(insen) into the systemic circulation and thereby seed the transformation of Prc to Prscwithin the bovine reticuloendothelial system and brain. This factor, combined with the very high resistance of PrPsc to degradation in, or on the ground, in the intestinal tract, in rendering procedures, or in silage plants suggests that recycling of PrPsc via the oral route may occur through feed contaminated with faeces.27-29
The proposal that the high paracellular leakage of colon to scrapie prion protein may be a cause of the high rate of its transmission from MBM in cattle and the relatively low rate in other species (particularly sheep), depends on a number of important assumptions. The first assumption is that differences in colonic permeability observed here, using in vitro methods, also hold true in vivo. A major permeability difference between sheep and cattle colon in vivo must exist, as cattle are unable to produce hard faeces in vivo.16 This failure is due to the high passive permeability of bovine colon to Na+.20 ,21Whether this difference in Na+ permeability is reflected in a higher permeability of cattle colon in vivo to macromolecules is still to be determined.
Another important assumption is that the inactivated hamster scrapie prion protein has a similar transmission rate to the biologically active BSE PrPsc. Denaturation studies with guanidinium HCl on scrapie prion show that the hydraulic radius of denatured prion protein is similar to that of the undenatured scrapie prion protein52 and that the size of the basic scrapie prion infectivity unit is around 30 kDa.26 ,33 It remains uncertain whether the paracellular route would permit movement of multimeric aggregates of prion fibrils.
It may also be inferred from this study that the general population of both sheep and humans could, fortuitously, be relatively resistant, in comparison with cattle, to horizontal transmission of BSE.53 Diseases such as Crohn's disease or high permeability states as in neonates may compromise this resistance in humans.2 ,7 ,8 ,11 ,15 ,51
Acknowledgments
The authors are very grateful for advice and assistance with radioiodination of HRP and prion protein from Dr Peter Jones, King's College London. Thanks are also extended to Dr James Hope, BBSRC Animal Health Unit, Edinburgh, now at BBRS Animal Health Unit, Compton, Berks. The authors are grateful to the Wellcome Trust for financial support.
Abbreviations used in this paper
- BSE
- bovine spongiform encephalitis
- DAB
- 3,3′-diamino-benzidine tetrahydrochloride
- HRP
- horseradish peroxidase
- MBM
- meat and bone meal
- MS
- mucosal to serosal
- PEG
- polyethylene glycol
- PrPsc
- scrapie prion protein
- ShaPrPsc
- Syrian hamster scrapie prion protein
- SM
- serosal to mucosal