Article Text

Abstract
BACKGROUND Measles virus is implicated in the aetiology of Crohn's disease. This measles hypothesis is mainly supported by immunohistochemical findings that the measles related antigen is present in the intestine of patients with Crohn's disease. Recently we isolated this antigen from the intestine of a patient with Crohn's disease using a molecular cloning technique and produced the monoclonal antibody against it (designated 4F12).
AIM To discover whether the measles related antigen is uniquely present in Crohn's disease.
SUBJECTS/METHODS Colonic mucosa samples from 20 patients with Crohn's disease, 20 with ulcerative colitis, 11 with non-inflammatory bowel disease (IBD) colitis, and nine controls were immunohistochemically stained with the anti-measles monoclonal antibody 4F12. The numbers of positive cells, the ratio of positive cells to nucleated cells, and the staining intensity of the positive cells were compared. Furthermore, the distribution of the measles antigen in other human organs was examined.
RESULTS Both the number of positive cells and the ratio of positive cells to nucleated cells were significantly increased in Crohn's disease, ulcerative colitis, and non-IBD colitis compared with controls (p<0.05) but were similar among the three disease groups. The staining intensity of the positive cells was also similar among the three disease groups. Small numbers of positive cells were observed in the oesophagus, stomach, duodenum, jejunum, and lung.
CONCLUSIONS The presence of the measles related antigen in the colonic mucosa was not unique to Crohn's disease. These results, together with the observation that such a measles related antigen was derived from host protein, do not support the hypothesis that measles virus causes Crohn's disease.
- Crohn's disease
- measles virus
- immunohistochemistry
- ulcerative colitis
- inflammatory bowel disease
- molecular mimicry
Statistics from Altmetric.com
- Crohn's disease
- measles virus
- immunohistochemistry
- ulcerative colitis
- inflammatory bowel disease
- molecular mimicry
Persistent measles virus infection has been implicated as a cause of Crohn's disease.1-7 It was first claimed by Wakefield et al,1 which even had social implications in the rejection of measles vaccination in the United Kingdom.8 This measles hypothesis is based mainly on immunohistochemical1-4 and epidemiological findings.5-7 Recent epidemiological investigations, however, did not support the association between measles virus and Crohn's disease.9-15 The immunohistochemical evidence derives essentially from the observation that both the monoclonal antibody and the polyclonal hyperimmune serum specific for measles virus recognised the measles related antigen in the intestine of patients with Crohn's disease.1-3 We confirmed this observation with the monoclonal antibody (MAS 182r: Harlan Sera-lab, Crawley Down, Sussex, UK) used by Wakefield et al.16 However, the highly sensitive reverse transcription polymerase chain reaction (PCR) has failed to identify any measles virus genome in the intestinal tissues of patients with Crohn's disease.17-20 Thus the measles hypothesis is very controversial.21
Our recent study,16 in which cDNA library from intestinal tissue was screened with the monoclonal antibody MAS 182r, disclosed the unexpected result that the measles related antigen found in the intestine of patients with Crohn's disease was derived not from measles virus but from an as yet unidentified human protein. Combined with the negative PCR results, this observation is clearly incompatible with the hypothesis that measles virus persistently infects the intestine of patients with Crohn's disease. Therefore, to investigate the measles hypothesis fairly, it was imperative to clarify whether the measles related antigen was uniquely present in the intestine of patients with Crohn's disease. In this study, we investigated its distribution in the colonic mucosa of patients with several types of colitis and controls by immunohistochemical methods. Furthermore, its distribution in other human organs was also examined.
Materials and methods
TISSUE SAMPLES
Colonic mucosa was taken with informed consent from 20 patients with Crohn's disease, 20 with ulcerative colitis, 11 with non-inflammatory bowel disease (IBD) colitis, and nine without colitis (controls) by endoscopic biopsy or surgical resection. As a rule, samples were taken from inflamed mucosa (margin of ulcer, erosion, aphthoid lesion, oedematous mucosa) in the patients with colitis or from macroscopically normal mucosa in the patients without (controls). In each case, the diagnosis was established by standard clinical, radiological, endoscopic, and histological criteria. Table 1 gives the clinical profiles of the patients.
Clinical profiles of patients from whom colonic mucosa was taken
PREPARATION OF THE MONOCLONAL ANTIBODY AGAINST MEASLES VIRUS
As previously reported,16 we screened 1.5 million clones from a λgt11 expression library, constructed from the intestine of a patient with typical Crohn's disease, with measles specific monoclonal antibody (MAS 182r), which recognises the measles virus nucleocapsid protein,1 and identified one positive clone. The double stranded DNA of the positive clone was isolated from the λgt11 vector gene and amplified by PCR. Then the PCR amplified DNA was inserted into the TA cloning vector (Invitrogen, San Diego, California, USA), and the nucleotide sequence of the positive clone determined by the DyeDeoxy Terminator Cycle Sequencing method (Applied Biosystems, a division of Perkin-Elmer Corporation, Cleveland, Ohio, USA). On the basis of the nucleotide sequence determined, genetic analysis of the positive clone was performed. Furthermore, a monoclonal antibody against the positive clone was produced.22Firstly, a Balb/c mouse was immunised with λgt11 expressed positive clone mixed with Freund's adjuvant, and then cell lines were created in vitro by fusing spleen lymphocytes with a myeloma cell line. Spleen and myeloma cells were fused and plated in a special medium containing hypoxanthine, aminopterin, and thymidine. Hybridoma that could produce a specific antibody to the positive clone was selected by enzyme linked immunosorbent assay (ELISA). A cell suspension of this hybridoma was injected into the peritoneum of the mouse. Thereafter, ascitic fluid was aseptically aspirated. After purification, the specificity of the monoclonal antibody, named 4F12, was examined by western blot23 and immunofluorescence.24 In addition, a double immunohistochemical study24 with two monoclonal antibodies, MAS 182r and 4F12, was performed using two staining substances, diaminobenzidine and 4-chloro-1-naphthol, to confirm that both monoclonal antibodies reacted with the same cells in the intestinal tissue.
We expressed the positive clone as a fusion protein with glutathione S-transferase (GST) using the GST gene fusion system (Amersham Pharmacia Biotech, Amersham, Buckinghamshire, UK).25Briefly, the double stranded DNA of the positive clone was ligated into the pGEX-4T-2 vector which can maintain the GST reading frame. Then competent Escherichia coli BL21 cells (Strategene, La Jolla, California, USA) were transformed by the ligation reaction and a fusion protein was expressed by adding 0.1 mM isopropyl β-d-thiogalactoside. A fusion protein was purified from the bacterial lysates by affinity chromatography using glutathione-Sepharose 4B, and eluted in glutathione elution buffer containing 10 mM reduced glutathione in 50 mM Tris/HCl (pH 8.0). Expression of the fusion protein was screened by both sodium dodecyl sulphate (SDS)/polyacrylamide gel electrophoresis (PAGE) and western blot.
IMMUNOHISTOCHEMISTRY
An immunohistochemical study was performed by the indirect immunoperoxidase staining method.26 27 Briefly, tissue pieces were fixed with periodate/lysine/4% paraformaldehyde. Then serial frozen sections were made and endogenous peroxidase activity was blocked with 5 mM periodic acid/3 mM sodium borohydride. The monoclonal antibody 4F12 was applied as the first antibody, and peroxidase labelled rabbit anti-mouse immunoglobulin (Dako, Copenhagen, Denmark) as the second antibody. For nuclear staining after the diaminobenzidine reaction, 0.1% methyl green was used. In addition, in the case of Crohn's disease, double immmunohistochemical staining using 4F12 and anti-CD68 monoclonal antibody (clone PG-M1, Dako) was performed.
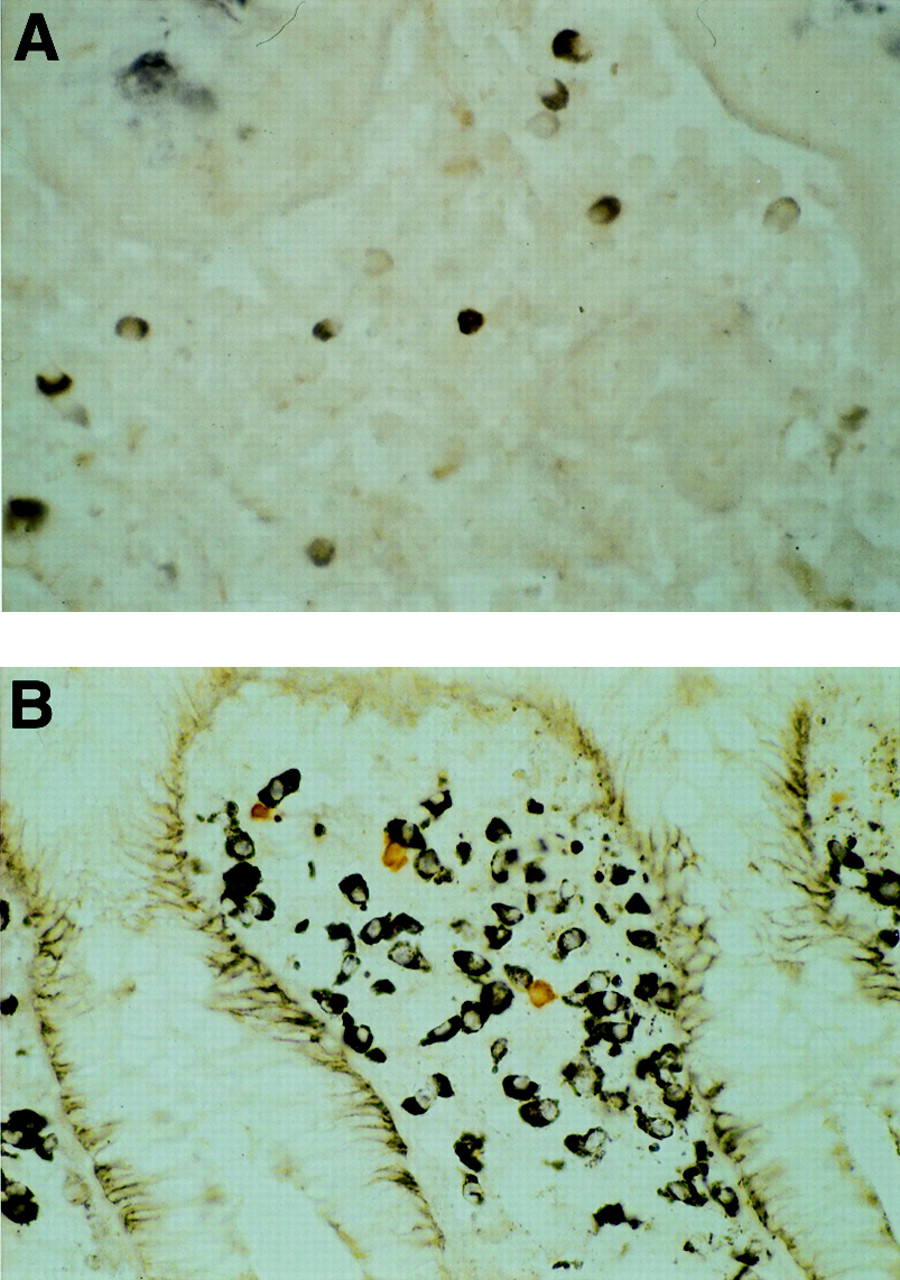
After nuclear staining, all samples were coded and three authors counted positive cells independently. The authors counted all positive cells in the lamina propria within the ½ view (400 × magnification) where positive cells were most densely observed. Positive cells in lymph follicles were excluded from the count. The numbers of positive cells were expressed as cells/mm2. Nucleated cells in the same area were also counted and the ratio of positive cells to nucleated cells was evaluated. In addition, the staining intensity of the positive cells was classified into three grades: strong (fig 1A), moderate (fig 1B), and weak (fig 1C). No positive cells were observed in the negative control sample (fig 1D), for which the incubation with 4F12 was omitted. We judged the staining intensity as strong when more than a half of the positive cells in the sample were stained as strongly as those shown in fig 1A.

Staining intensity of positive cells: (A) strong; (B) moderate; (C) weak. No positive cells are observed in the negative control sample (D).
The data on numbers of positive cells and the ratio of positive cells to nucleated cells were analysed for statistical significance by the Mann Whitney U test to compare mean values between groups; proportions of the staining intensities of each group were compared by Fisher's exact test.
We also examined the distribution of 4F12 positive cells in several human organs (oesophagus, stomach, duodenum, jejunum, liver, lung, spleen, kidney, heart) taken at autopsy from a 43 year old female patient (cause of death mitral stenosis).
Results
PREPARATION OF MONOCLONAL ANTIBODY
Genetic analysis of the positive clone
A homology search showed 99% of the nucleotide sequence of the positive clone was identical with that of the human gene (AA449055) (fig 2), but no appreciable homology was found between the positive clone and the measles virus. Figure 3 shows the deduced amino acid sequence of the positive clone. We could not find any definite homologous amino acid sequence in the protein data bases.

Multiple alignment of the nucleotide sequences between the positive clone and AA449055. Nucleotide sequences of non-homologous regions are shown in bold and underlined.

Deduced amino acids sequence of the positive clone. Each amino acid is shown using the single letter code.
Specificity of 4F12
SDS/PAGE showed a fusion protein containing the positive clone and β-galactosidase at 130 kDa (fig 4). The western blot showed that both MAS 182r and 4F12 formed strong bands with the fusion protein and that no positive band was formed between 4F12 and untransformedE coli extracts (fig 4). We also confirmed by western blot that 4F12 did not react with β-galactosidase(data not shown).

SDS/PAGE of the positive clone expressed by Escherichia coli as a fusion protein with β-galactosidase, and western blot analysis of the fusion protein (the positive clone and β-galactosidase) and anti-measles monoclonal antibodies.
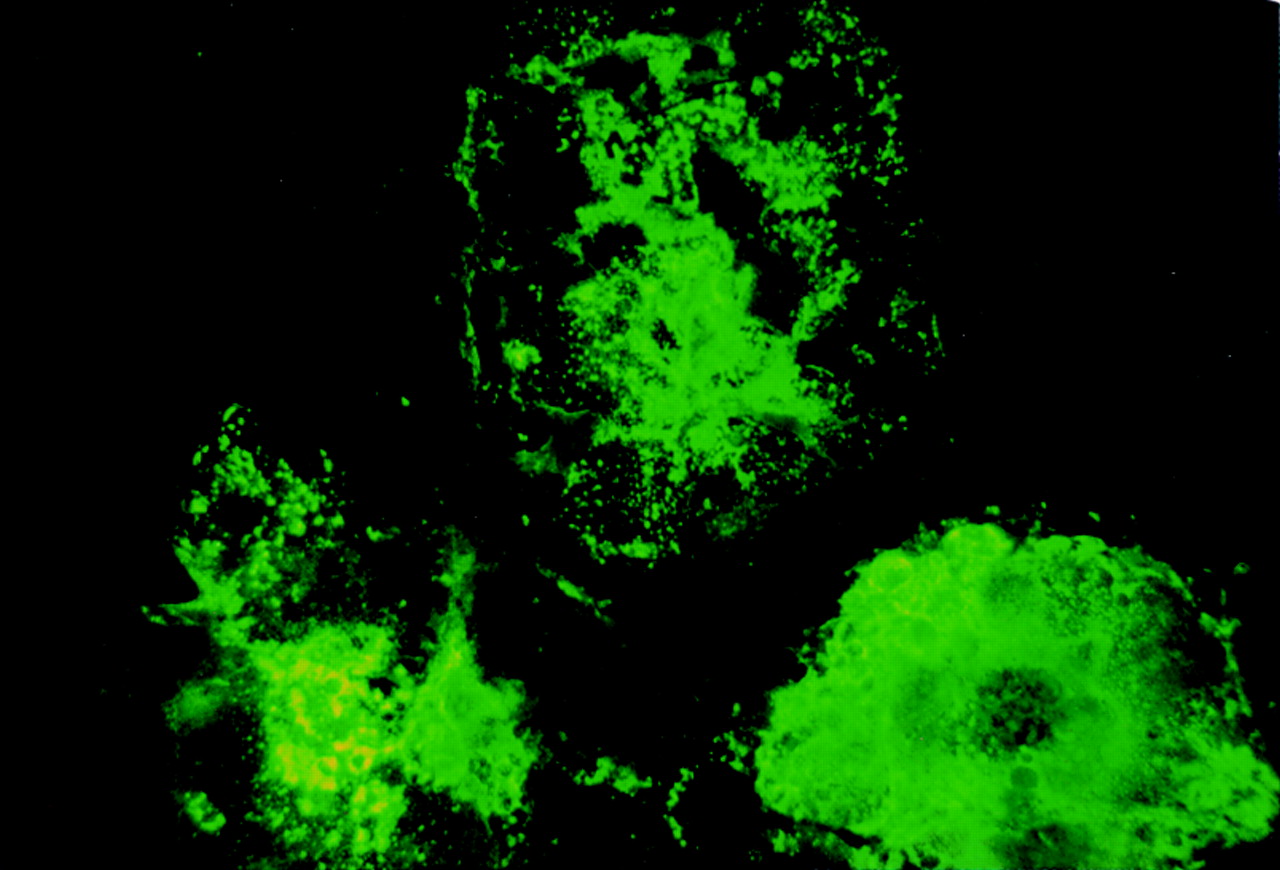
The immunofluorescence study clearly shows that measles virus infected cells reacted with 4F12 (fig 5) as well as with MAS 182r (data not shown). Cells not infected with measles virus were not stained with either 4F12 or MAS 182r (data not shown). The double immunohistochemical study showed that virtually all MAS 182r positive cells were stained with 4F12 (fig 6A) and vice versa.
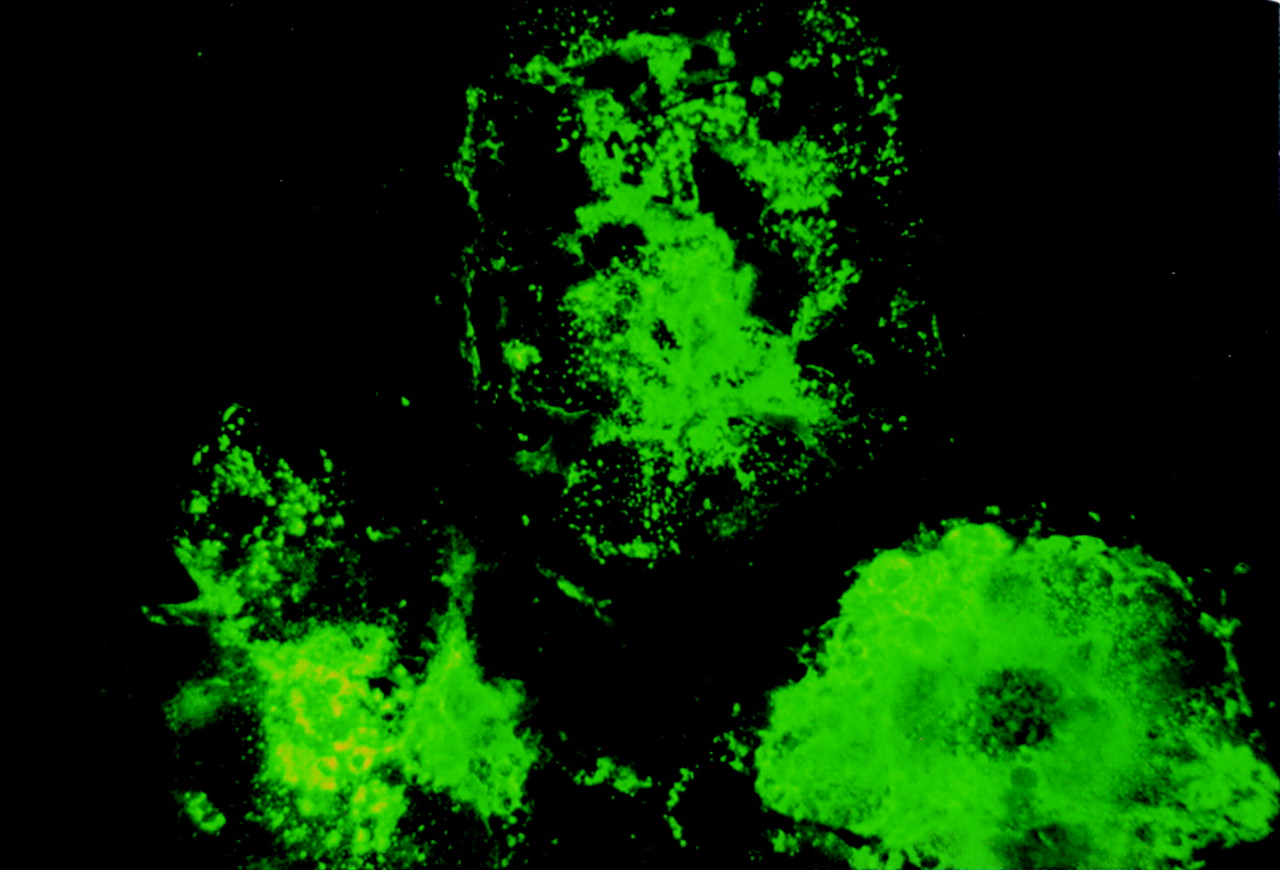
Double immunohistochemical study with MAS 182r and4F12. Virtually all positive cells are doubly stained with MAS 182r and4F12 (A). Doubly stained cells appear as a mixture of dark blue and brown. Cells stained singly with either 4-chloro-1-naphthol (dark blue) or diaminobenzidine (brown) are shown in (B).
Expression of a fusion protein of the positive clone using the GST gene fusion system
SDS/PAGE showed the positive clone as a fusion protein of GST at 45 kDa (fig 7). Ladder bands of less than 45 kDa are assumed to be partially decomposed fusion proteins. The thin band at about 70 kDa is suggested to be an E coli dnaK protein that was sometimes co-purified with GST recombinant protein.28The western blot showed that both MAS 182r and 4F12 formed positive bands with the fusion protein and partially decomposed fusion proteins (fig 7). No positive band was formed between 4F12 and the purified GST (fig 7).

SDS/PAGE of the positive clone expressed as a fusion protein with GST, and western blot analysis of the fusion protein and anti-measles monoclonal antibodies.
IMMUNOHISTOCHEMISTRY
Observation of 4F12 positive cells
Positive cells were observed in all tissue samples. All positive cells were stained in the cytoplasm, and some were stained in both the cytoplasm and the nucleus. As Wakefield and colleagues1reported, positive cells comprised macrophages, lymphocytes, vascular endothelial cells, fibroblasts, and neutrophils. The double immunohistochemical study showed that some positive cells were doubly stained with anti-CD68 monoclonal antibody.
Numbers of positive cells
Figure 8 shows the numbers of positive cells. The mean (SE) number of positive cells was 22.2 (4.7) in Crohn's disease, 21.6 (3.5) in ulcerative colitis, 25.2 (6.7) in non-IBD colitis, and 8.9 (1.5) in controls; the increase in Crohn's disease, ulcerative colitis, and non-IBD colitis compared with controls was significant (p<0.05). No significant difference was found between the three disease groups.

Numbers of positive cells in the colonic mucosa. The number of positive cells for each sample is shown as a closed circle. The mean and SE of the numbers of positive cells in each group is shown as an open circle and vertical bar. IBD, inflammatory bowel disease.
Ratio of positive cells to nucleated cells
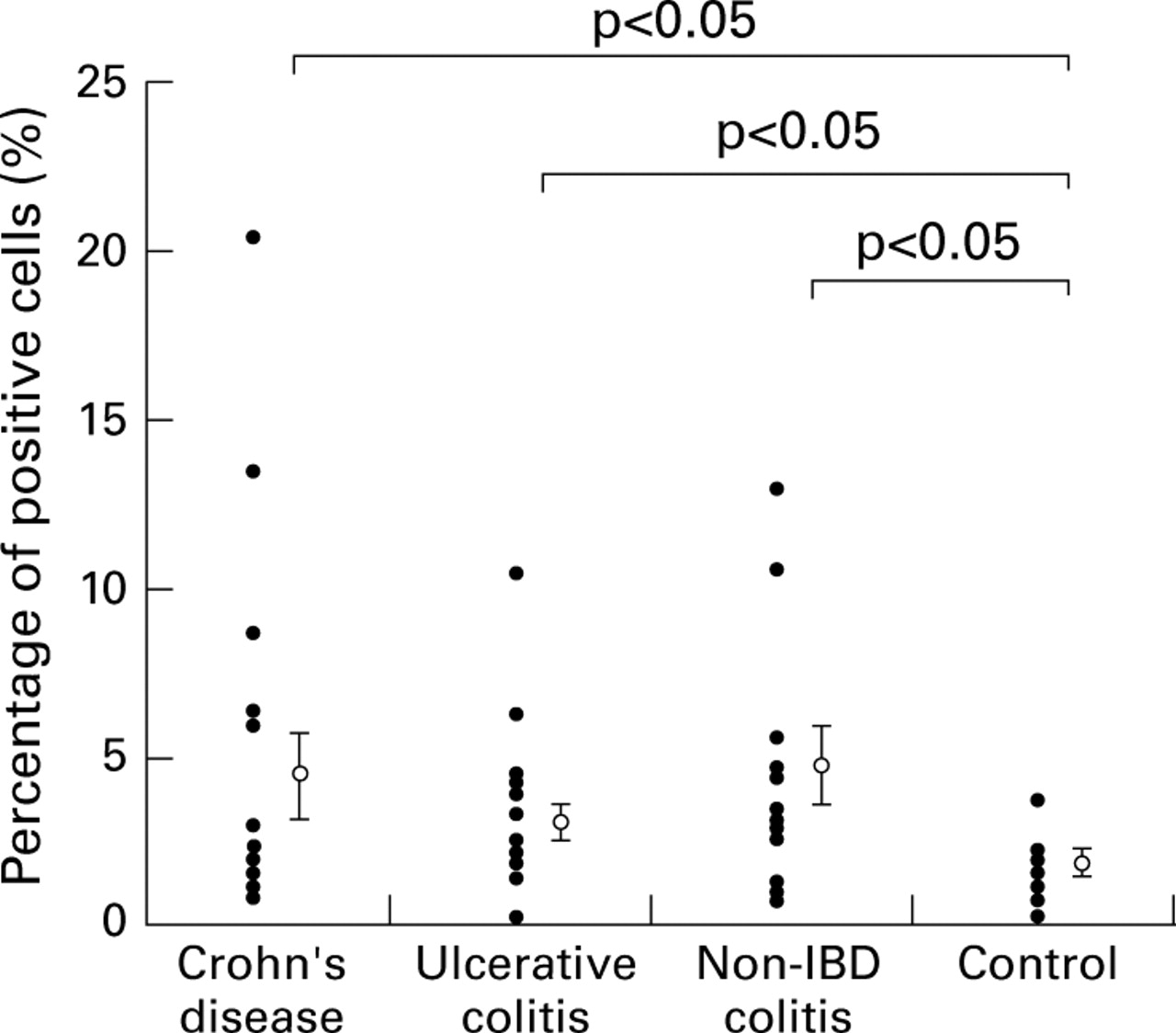
The ratio of positive cells to all nucleated cells is expressed as a percentage and shown in fig 9. The mean (SE) was 4.3 (1.1) in Crohn's disease, 2.9 (0.5) in ulcerative colitis, 4.6 (1.2) in non-IBD colitis, and 1.6 (0.3) in controls, representing a significant increase in Crohn's disease, ulcerative colitis, and non-IBD colitis compared with controls (p<0.05). No significant difference was found between the three disease groups.

Ratio of positive cells to nucleated cells expressed as a percentage. The percentage for each sample is shown as a closed circle, and the mean and SE for each group is shown as an open circle and vertical bar.
Staining intensity of positive cells
Figure 10 shows the proportions of the staining intensities of the positive cells in each group. Positive cells were stained strongly in 25% in Crohn's disease, 20% in ulcerative colitis, 27% in non-IBD colitis, but 0% in controls, and were stained moderately in 45% in Crohn's disease, 35% in ulcerative colitis, 18% in non-IBD colitis, and 22% in controls. A significant difference (p<0.05) was found between Crohn's disease and controls, but no significant difference was found between the three disease groups.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Proportions of the staining intensities in each group. The proportion staining strongly is shown as black, that staining moderately as dark grey, and that staining weakly as grey.
Distribution of positive cells in human organs
Small numbers of positive cells were observed in tissue samples from the oesophagus, stomach, duodenum, jejunum, and lung, but no positive cells were identified in tissue samples from the liver, spleen, kidney, and heart (data not shown).
Discussion
This study does not show that the measles related antigen is exclusively present in the intestine in Crohn's disease; it was also present in the intestine in ulcerative colitis and non-IBD colitis. These results are not in accordance with the previous immunohistochemical study by Wakefield et al,1 in which positive cells were observed in the intestine of patients with Crohn's disease but not in the intestine of two patients with intestinal tuberculosis. In this study we used the monoclonal antibody 4F12 instead of MAS 182r which was used by Wakefield et al, as we found that 4F12 and MAS 182r recognised the same epitope and that both monoclonal antibodies reacted with the same cells in the intestine on immunohistochemical examination. In our study positive cells were weakly stained in more than half of the colonic samples from patients with non-IBD colitis. Therefore it is possible that the weakly stained cells were regarded as negative by Wakefield et al.
We recently reported that the origin of the measles related antigen found in the intestine in Crohn's disease was not measles virus but an as yet unidentified human protein.16 These results, together with the negative reverse transcription PCR reports,17-20 are incompatible with the hypothesis that measles virus persistently infected the intestine in Crohn's disease. The immunohistochemical evidence supporting the measles hypothesis is based on the theory that measles antibody recognises measles virus itself and the measles virus antigen is uniquely present in Crohn's disease. Our results directly contradict this theory, thereby making the measles hypothesis less attractive. We further analysed the antigenicity of the positive material called “measles related antigen”. Western blot analysis showed that both monoclonal antibodies, MAS 182r and 4F12, reacted with each fusion protein of the “measles related antigen”—that is, the one constructed from the “measles related antigen” and GST and that constructed from the “measles related antigen” and β-galactosidase. Conversely, both monoclonal antibodies similarly reacted with measles virus infected cells. As the “measles virus related antigen” was not detected in human kidney, we consider that both monoclonal antibodies reacted with the measles virus itself in the measles virus infected Vero cells which are derived from monkey kidney cell lines. From these observations, we think that MAS 182r and 4F12 recognise the identical epitope and that such an epitope can be expected to be shared between the “measles related antigen” and the measles virus. Therefore we hypothesise that anti-measles antibody cross reacted with the host derived “measles related antigen” in the intestine in Crohn's disease which was therefore regarded as measles virus in the previous study. Although the possibility still remains that, in the present study, the antigen recognised in colonic mucosa by 4F12 is measles virus antigen (the measles specific antigen), we assume such a possibility is extremely slight for the following reasons. If 4F12recognised “the measles specific antigen” in the colonic mucosa of Crohn's disease, some measles virus genes ought to be identified by screening with anti-measles monoclonal antibody the 1.5 million clone cDNA library constructed from the intestine of a patient with Crohn's disease. However, no measles virus genes were identified by the screening. Moreover, it is contradictory that the “measles specific antigen” increases temporally in the colonic mucosa in non-IBD colitis, which includes campylobacter colitis, intestinal tuberculosis, etc.
As described above, the “measles related antigen” has not been identified, as no homologous proteins have been found in the protein data bases. However, the nucleotide sequence of the positive clone, which is thought to be a gene fragment, was 99% homologous with the human gene (AA449055) deposited in DNA data bases. A Southern blot analysis further confirmed the human origin of the “measles related antigen”.16 It is not known what role the “measles related antigen” plays in the intestine. In view of our immunohistochemical results showing that it was increased in inflamed colonic mucosa and observed in the immunocytes, it may be a byproduct of inflammation or belong to the group of cytokines or other chemical mediators. The double immunohistochemical study showed that many 4F12positive cells also expressed the CD68 antigen, suggesting that they are macrophages. Previous studies29 30 have reported that some macrophage subpopulations are increased in the inflamed intestinal tissues in IBD. Therefore it could be suggested that the “measles related antigen” is associated with a subpopulation of macrophages.
On the other hand, the possibility still exists that the host derived protein of the “measles related antigen” is the target of autoimmune response through molecular mimicry because it is assumed to share a common epitope with measles virus. The idea that antibodies to infectious agents cross react with normal human tissues has been previously put forward.31-33 Srinivasappa and colleagues31 examined the cross reactivities of 600 anti-viral monoclonal antibodies and showed that about 3.5% of them reacted with normal mouse tissues. With reference to anti-measles monoclonal antibodies, cross reactivities with human intermediate filaments,32 mouse anterior pituitary, gastric mucosa, salivary gland, and neurones in the brain31 have been reported. Cunningham and colleagues33 reported that monoclonal antibodies that recognise epitopes on both enteroviruses andStreptococcus pyogenes were cross reactive with human cardiac myosin and such antibodies may be associated with autoimmune heart disease. Thus the “measles related antigen” may be the target of the autoantibody and may evoke intestinal damage in hosts that can produce antibody against it. However, further studies are needed to validate this autoimmune hypothesis.
In conclusion, our results clearly show the invalidity of the immunohistochemical evidence for the measles hypothesis and therefore they do not support the hypothesis that persistent measles virus infection is implicated in Crohn's disease.
Acknowledgments
We thank Dr Sugita, Yokohama City University School of Medicine, for providing the colonic samples, Dr Kawamura, Akita University School of Medicine, for providing tissues from human organs, and Miss Pamyla Yates, Minnesota State University in Akita, for her critical review of the manuscript. This work was supported in part by a grant from the Kanae Foundation of Research for New Medicine and Grant-In-Aid for Scientific Research from the Ministry of Education, Science and Culture, and a grant from the Ministry of Health and Welfare, Japan.
References
Footnotes
- Abbreviations used in this paper:
- IBD
- inflammatory bowel disease
- PCR
- polymerase chain reaction
- ELISA
- enzyme linked immunosorbent assay
- GST
- glutathione S-transferase
- SDS
- sodium dodecyl sulphate
- PAGE
- polyacrylamide gel electrophoresis