Article Text

Abstract
BACKGROUND Mucins play an important protective role in the colonic mucosa. Luminal factors modulating colonic mucus release have been not fully identified.
AIM To determine the effect of some dietary compounds on mucus discharge in rat colon.
METHODS An isolated vascularly perfused rat colon model was used. Mucus secretion was induced by a variety of luminal factors administered as a bolus of 1 ml for 30 minutes in the colonic loop. Mucin release was evaluated using a sandwich enzyme linked immunosorbent assay supported by histological analysis.
RESULTS The three dietary fibres tested in this study (pectin, gum arabic, and cellulose) did not provoke mucus secretion. Luminal administration of sodium alginate (an algal polysaccharide used as a food additive) or ulvan (a sulphated algal polymer) induced a dose dependent increase in mucin discharge over the concentration range 1–25 mg/l (p<0.05 for 25 mg/l alginate and p<0.05 for 10 and 25 mg/l ulvan). Glucuronic acid and galacturonic acid, which are major constituents of a variety of fibres, produced significant mucin secretion (p<0.05). Hydrogen sulphide and mercaptoacetate, two sulphides produced in the colonic lumen by microbial fermentation of sulphated polysaccharides, did not modify mucin secretion. Among the short chain fatty acids, acetate (5–100 mM) induced a dose dependent release of mucus (p<0.05 for 100 mM acetate). Interestingly, butyrate at a concentration of 5 mM produced colonic mucin secretion (p<0.05), but increasing its concentration to 100 mM provoked a gradual decrease in mucus discharge. Propionate (5–100 mM) did not induce mucin release. Several dietary phenolic compounds (quercetin, epicatechin, resveratrol) did not provoke mucus discharge.
CONCLUSIONS Two algal polysaccharides (alginate and ulvan), two uronic acids (glucuronic acid and galacturonic acid), and the short chain fatty acids acetate and butyrate induce mucin secretion in rat colon. Taken together, these data suggest that some food constituents and their fermentation products may regulate the secretory function of colonic goblet cells.
- mucin secretion
- colon
- dietary fibres
- algal fibres
- short chain fatty acids
- sulphides
Statistics from Altmetric.com
Intestinal mucins are high molecular mass glycoproteins which form a highly hydrated mucus gel coating the epithelial surface of the intestinal tract.1 Secreted by goblet cells, intestinal mucins are involved in a variety of cytoprotective functions against a number of hazards including mechanical insults, colonisation by pathogenic bacteria and their toxins, luminal proteases arising from bacterial and mucosal cells, and potential carcinogens.2
As mucins play a key role in the protection of the underlying epithelium, any quantitative change in mucus secretion may modify this defensive barrier and have important physiological implications.3 4 In particular, the interactions of dietary components with the secretory activity of goblet cells could represent new interesting possibilities for the manipulation of this important protective function. However, our knowledge of dietary factors that control the release of intestinal mucus is limited. Previous studies have simply shown by ultrastructural,5 6autoradiographic,7 and enzyme linked immunosorbent assay (ELISA) techniques8 9 that dietary supplementation with fibres increases the secretory activity of intestinal goblet cells. However, the short term effect of dietary fibres on colonic mucus secretion was not directly examined. Besides dietary fibres, a variety of algal polysaccharides are also found in human food as additives.10 These polysaccharides exhibit several unusual structural and chemical properties, especially the presence of sulphated groups (carrageenans).11 The physiological effects of these algal fibres on mucus secretion have not yet been investigated.
The effects of fibres, as observed in vivo on mucus cells, may be either direct or indirect, through the end products of their degradation by bacteria. Short chain fatty acids, which play important physiological roles in the colon, are the major products of colonic fermentation.12-14 Other components such as sulphides are also produced from sulphated fibres on degradation by colonic bacteria.15 16 Nothing is known about the influence of these end products on goblet cell secretory activity.
Several non-nutritive constituents such as polyphenols are also contained in a variety of fruits and vegetables. Dietary phenolic compounds seem to exert several pharmacological effects such as antioxidative and anticarcinogenic ones.17-19 Whether these compounds are involved in the regulation of colonic mucus secretion was therefore tested.
It thus appears that colonic goblet cells may be under the control of numerous primary or secondary luminal factors, which makes the interpretation of most results gathered from in vivo experiments difficult. The present study was thus undertaken to investigate further the discharge of colonic goblet cells in response to several compounds occurring at the apical side of the goblet cells. For this purpose, the isolated vascularly perfused rat colon was used. This model provides a unique opportunity to study the secretion of mucus in response to well defined luminal stimuli in a manner that eliminates the influences potentially encountered in vivo. The polarised mucus cells may be thus submitted to specific stimulation with fibres or products of carbohydrate fermentation. The discharge of mucus was evaluated by a sandwich ELISA and histological analysis.
Materials and methods
MATERIALS
Bovine serum albumin was purchased from Biovalori (Cassens, France). Hyperamine 25, a mixture of amino acids, was obtained from Braun Medical (Boulogne, France). The following reagents were purchased from Sigma (Saint Quentin-Fallavier, France): cellulose, pectin, gum arabic, carrageenin iota, carrageenin kappa, and carrageenin lambda, fucoidan, glucuronic acid, galacturonic acid,n-butyrate, acetate, propionate, resveratrol, quercetin, epicatechin, sodium mercaptoacetate, sodium hydrogen sulphide, and dithiothreitol. Alginic acid sodium salt was purchased from Fluka (Saint Quentin-Fallavier, France). Ulvan (a new source of dietary fibre extracted from green seaweed) was kindly provided by C Cherbut (INRA, Nantes, France). Microtitre plates (NUNC-Immunoplate) were obtained from Polylabo (Paul block & cie, Strasbourg, France).
SURGICAL PREPARATION
The operative procedure to prepare an isolated vascularly perfused rat colon was performed as previously described.20Briefly, male Wistar rats (250–350 g), purchased from Le Centre d'Elevage Dépré (Saint Doulchard, France), were anaesthetised with sodium pentobarbital (50 mg/kg, intraperitoneally). The proximal and transverse colons were freed from their visceral fixations. The ileocolic veins and arteries were tied. Both ends of the colonic loop (10 cm in length) were then fitted with Silastic tubing. A metal cannula and a Silastic cannula were then quickly inserted into the superior mesenteric artery and portal vein respectively. Arterial perfusion was started immediately at a rate of 2.5 ml/min with Krebs-Henseleit buffer (2 mM CaCl2, 6 mM KCl, 3.18 mM NaH2PO4, 104 mM NaCl, 1 mM MgSO4, 41.6 mM NaHCO3, pH 7.4) containing 25% washed bovine erythrocytes, 3% bovine serum albumin, 5 mM glucose, and 1% (v/v) hyperamine 25. The perfusion pressure, continuously recorded with a mercury manometer, ranged from 55 to 75 mm Hg. The colonic lumen was flushed out with prewarmed isotonic saline and then with air, and both ends of the intestinal loop were ligated. The preparation was then removed and transferred to a bath containing isotonic saline at 37°C.
EXPERIMENTAL PROTOCOL
The experiments consisted of a 10 minute equilibration period, followed by a 30 minute period of stimulation. Immediately after the equilibration period, the loops were filled by injection with 1.0 ml of prewarmed isotonic saline (control groups) or with 1 ml of luminal factors at 37°C. All luminal agents to be tested were prepared freshly before the stimulations started. The pH of each luminal compound was adjusted to 7–7.5, and the osmolarity was increased when required to 300 mosmol/kg by the addition of appropriate amounts of NaCl. Control data were obtained from separate groups of animals.
At the end of the experimental period, loops were cut at both ends. Fluid content was collected. To remove the mucus adherent to the mucosal surface, loops were carefully emptied by manual massage, subsequently flushed with 15–20 ml air, and drained. Luminal contents (fluid contents + adherent mucus gel) were weighed, sonicated, and frozen at −20°C for subsequent determination of mucin-like immunoreactivity and luminal DNA content. The adherent mucus gel was taken into account because it represents a substantial part of the secreted immunoreactive material. The empty colonic loops were weighed and the length measured (cm). They were then homogenised for one minute in phosphate buffered saline (Ultra-Turrax; Janke and Kundel, Staufen, Germany), and stored at −20°C. Tissue homogenates were analysed for DNA content.
ANALYSIS OF SAMPLES
Samples of luminal contents were incubated for 24 hours with 100 mM dithiothreitol to produce reduction and then assayed by a sandwich type ELISA as previously described.21 Briefly, the polyclonal antibody 45C was raised in a rabbit against rat colonic mucus glycoprotein obtained from CsCl gradient purified material. An immunoglobulin rich fraction was prepared using DEAE-Sephadex A50 (pH 7.2), and then used to coat ELISA plates. A part of this immunoglobulin rich fraction was also used to prepare biotinylated labelled antibody using a succinimide ester of biotin.21 Mucin content of the samples was determined from standard curves prepared from purified rat colonic mucin. The luminal contents were tested at three dilutions (1:4000; 1:8000, and 1:16 000), each in duplicate. Samples of luminal stimulants were also assayed concurrently to verify that they did not generate false positive results in ELISA.
Colonic loop homogenates were thawed and briefly homogenised. Aliquots were sonicated for 20 seconds, and then analysed for DNA content using the fluorometric method of Hinegardner.22 The amount of mucin secreted from each loop was expressed as μg mucin/mg tissue DNA. Samples of luminal content were also analysed for DNA content. This analysis served as an indirect measure of tissue viability, and loops were discarded when luminal DNA content represented more than 2% of the total DNA (tissue DNA + luminal DNA). Total luminal content (fluid content + adherent mucus gel) recovered from the lumen was expressed as mg luminal contents/cm colonic loop.
HISTOLOGICAL ANALYSIS AND GOBLET CELL MORPHOLOGY
Histological technique
The effect of several dietary components was also monitored by histological analysis, as described previously.23 Briefly, pieces of colon (2–2.5 cm) taken at the end of the isolated colon experiment were fixed into Karnovsky's fixative, processed in Epon, cut into 2 μm sections, stained with alcian blue (pH 2.5) followed by the periodic acid-Schiff reaction and then counterstained with haematoxylin. The alcian blue/periodic acid-Schiff method yielded a blue colour when mostly acidic mucins were present, purple when neutral mucins were also present, and magenta when mainly neutral mucins were present. The corresponding cells were designated stained mucus cells.
Quantitative histology
It is generally accepted that a reduced number of mucus cells indicates release of mucins from the colonic mucosa, and cavitation of mucus cells is the hallmark of accelerated mucus secretion.1 24 25 Thus the extent of mucus discharge may be quantified both by assessing the number of mucus cells per crypt section and the percentage of cavitated mucus cells per preparation. For this purpose, 15–30 crypts per slide were analysed, and 500–1000 mucus cells were counted per sample of colon. A crypt was considered when it was cut along or nearly along the length of the crypt lumen (at least two thirds of the length of the crypt). Progressing from the crypt base to crypt opening, all stained mucus cells were counted on both sides of the lumen. All mucus cells were evaluated for the presence of cavitation of their apical membrane. A mucus cell with clear apical indentation into the intracellular store of mucus granules was taken as showing signs of the cavitation that accompanies recent compound exocytotic activity. Four to seven rat colons were analysed for each set of experiments. All slides were analysed by a single investigator who was blinded to the drug treatment.
STATISTICAL ANALYSIS
Data are presented as mean (SE), and compared using analysis of variance with a Bonferroni test. p<0.05 was considered significant.
Results
EFFECT OF FIBRES ON COLONIC MUCUS SECRETION
Non-sulphated fibres
Luminal administration of sodium alginate (25 mg/l) in the isolated vascularly perfused rat colon induced colonic mucin release (310.1 (24.3) μg/mg DNA (n = 10) v 131.9 (6.5) μg/mg DNA for the control group (n = 10) p<0.05). Histological analysis disclosed that colonic preparations exposed to 25 mg/l sodium alginate for 30 minutes exhibited numerous cavitated goblet cells (28.7 (4.4)% of cavitated mucus cells (n = 6) v10.6 (2.4)% of cavitated mucus cells for the control group (n = 6) p<0.05). In contrast, sodium alginate did not modify the number of stained mucus cells per crypt: 26.2 (0.6) mucus cells per cryptv 29.8 (0.6) mucus cells per crypt for the control group (n = 6). Stimulation with 1 or 10 mg/l sodium alginate did not elicit mucus discharge (175.2 (21.9) μg/mg DNA (n = 5) and 190.2 (14.8) μg/mg DNA (n = 6) respectively).
Compared with sodium alginate, luminal administration of pectin, gum arabic, or cellulose (25 mg/l) did not induce significant release of mucins (table 1). Even at final concentrations of up to 10 g/l, pectin and cellulose did not significantly stimulate the discharge of mucus (152.1 (4.6) μg/mg DNA (n = 5) and 120.5 (33.1) μg/mg DNA (n = 6) respectively). High concentrations of gum arabic were not used because of the high viscosity of the resulting mixture.
Mucin and total luminal content of fluid after administration of non-sulphated fibres
Sulphated polymers
Intracolonic administration of ulvan (25 mg/l), a sulphated algal polysaccharide, produced a sharp rise in the discharge of immunoreactive material (fig 1). This mucin discharge was associated with a rise in the percentage of cavitated mucus cells (table 2, fig2). As shown in fig 1, the effect of ulvan was concentration dependent over the range 1–25 mg/l. The first significant response was obtained with 10 mg/l ulvan. In contrast, ulvan did not elicit any significant increase in total fluid luminal content (table 2).

Colonic mucin-like immunoreactivity (μg/mg DNA) to luminal administration of increasing amounts of ulvan in the isolated perfused rat colon. The secretion of mucin was evaluated by a sandwich ELISA as described in Materials and methods. Data are mean (SE). *p<0.05 v controls.
Quantitative histology, ELISA, and total fluid luminal content after administration of several algal sulphated polymers (25 mg/l)
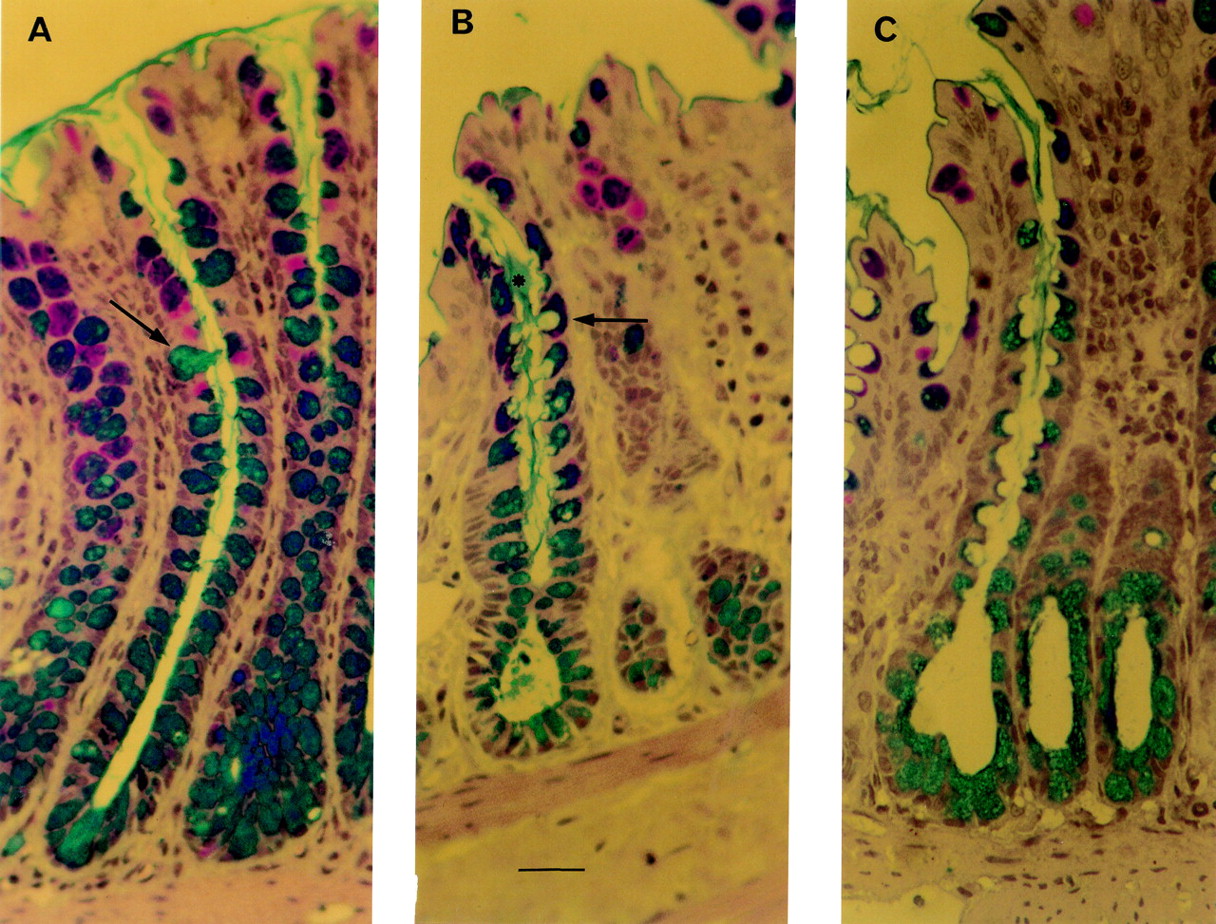
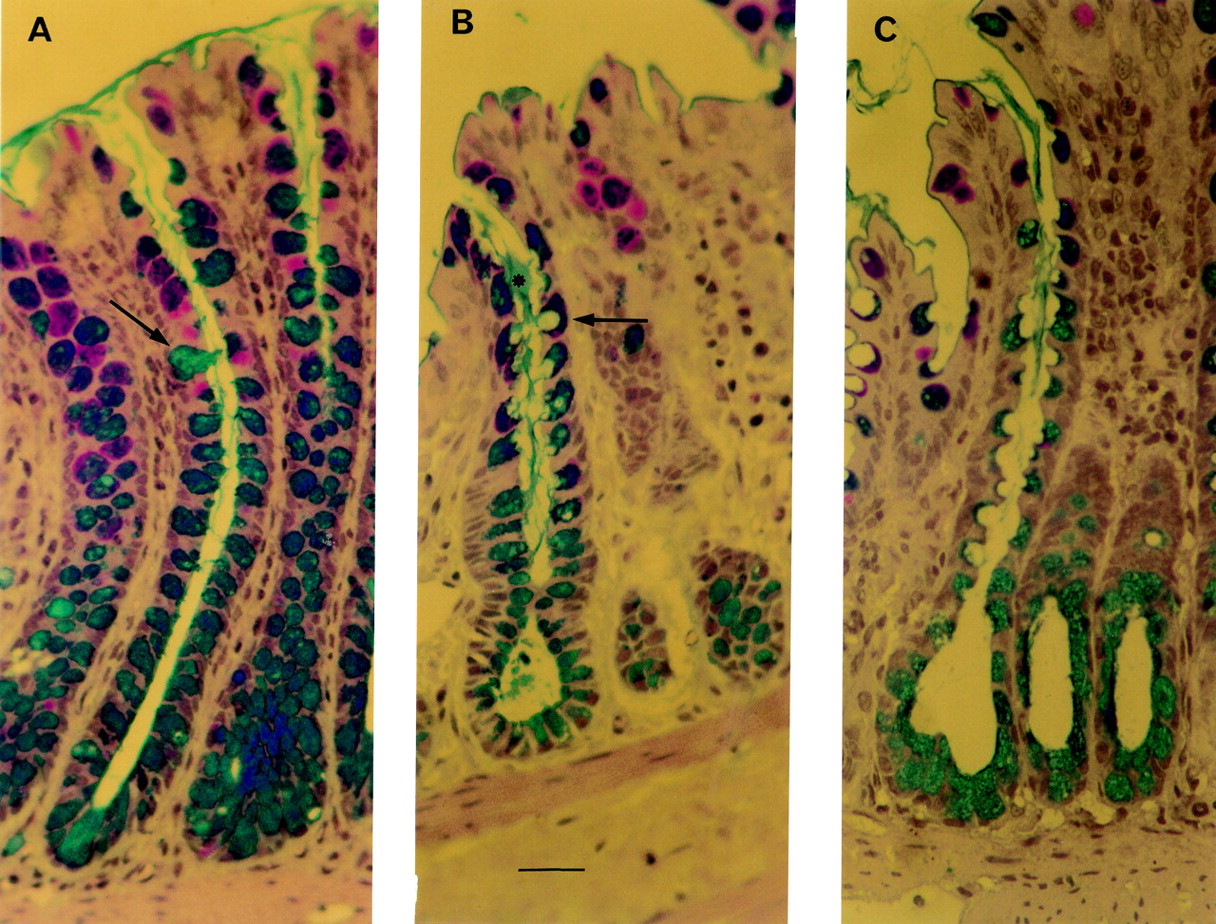
Thin section histology of perfused rat colon. Bar = 10 μm. (A) Crypt of a control preparation. This colonic loop underwent a 10 minute equilibration period, followed by a 30 minute period during which 1 ml isotonic saline was administered to the lumen of the colonic loop. Goblet cells (alcian blue/periodic acid-Schiff stained) are filled with mucus. Arrows indicate examples of stained mucus cells. (B) Colonic crypt after administration of ulvan (25 mg/l) for 30 minutes. Stained mucus cells exhibit deep apical membrane cavitation (arrow) from which streams of stained mucus (asterisk) emerged to join the mucus blanket in the crypt lumen. (C) Crypt of perfused rat colon after 30 minutes of stimulation with 5 mM butyrate. Numerous mucus cells with deep cavitation of the apical surface are easily observed.
Fucoidan (25 mg/l), another sulphated polymer, showed a tendency to increase mucin secretion (254.3 (30.2) μg/mg DNA, n = 6) although this increase did not achieve statistical significance (table2).
Undegraded iota, kappa, and lambda carrageenans, which are used as alimentary additives (gelling agents), contain one, two, or three SO4 2− groups respectively.10Undegraded iota, kappa, and lambda carrageenans at final concentrations of up to 25 mg/l did not produce any discharge of mucin whereas a significant increase in total luminal content of fluid was observed with the kappa and lambda carrageenans (table 2).
Uronic acids
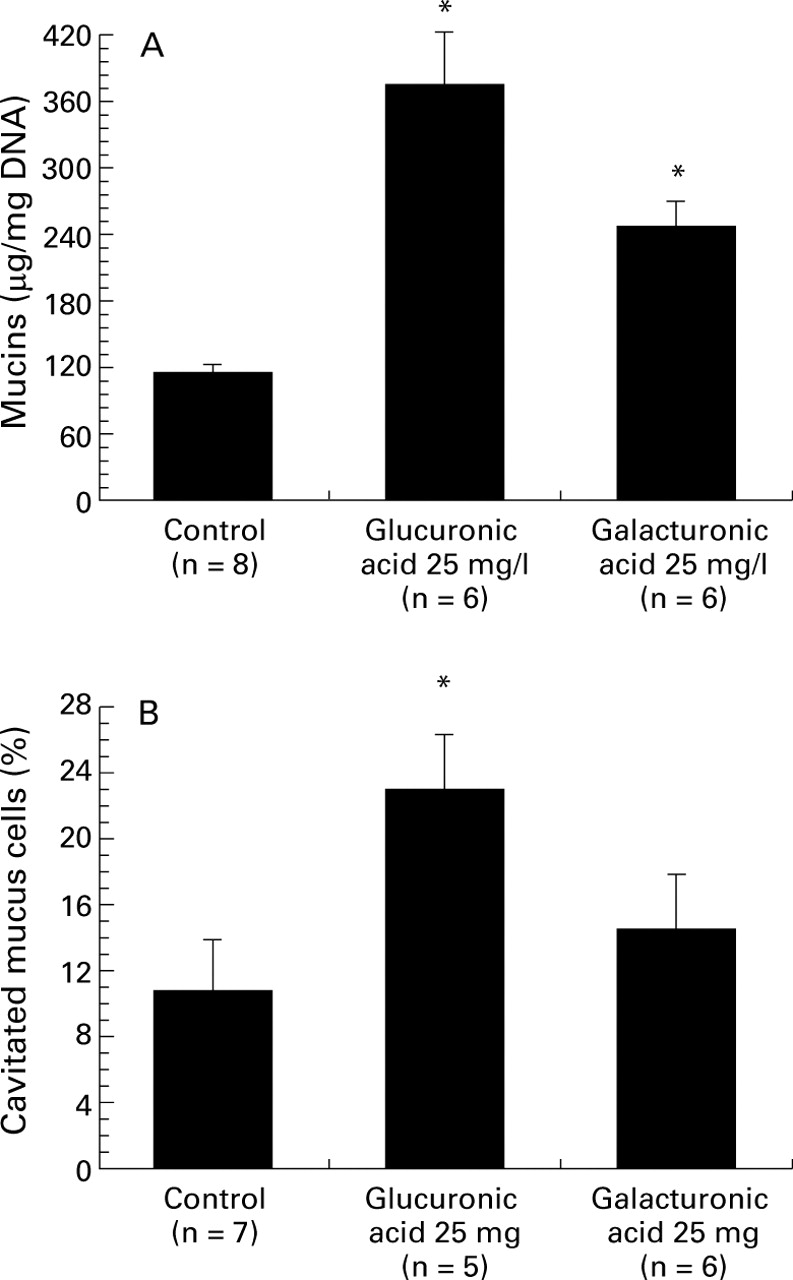
As outlined in fig 3, luminal administration of 25 mg/l glucuronic acid (one of the major constituents of ulvan) produced a sharp rise in the secretion of mucin (376.5 (45.6) μg/mg DNA (n = 6)v 115.2 (7.2) μg/mg (n = 8) for the control group, p<0.05; fig 3A). Numerous goblet cells became cavitated after 30 minutes of 25 mg/l glucuronic acid stimulation (fig 3B). On stimulation with 25 mg/l galacturonic acid, the rise in mucin release was less pronounced (247.1 (22.8) μg/ml DNA; n = 6, p<0.05; fig 3A). Morphological analysis indicated that galacturonic acid tended to increase the percentage of cavitated mucus cells, although this value did not reach statistical significance (fig 3B).
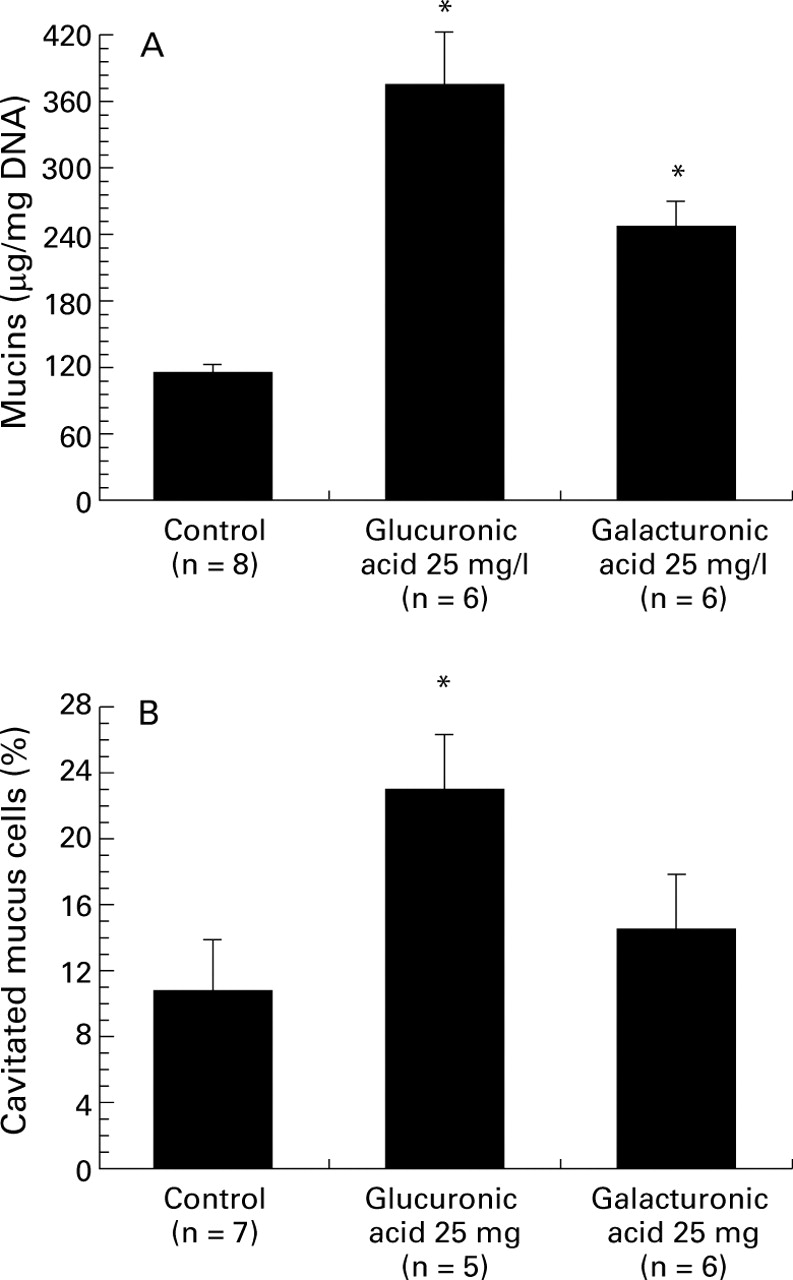
Effects of two uronic acids on colonic mucus cells. (A) Effects of glucuronic acid (25 mg/l) and galacturonic acid (25 mg/l) on luminal mucin secretion in the isolated vascularly perfused rat colon. *p<0.05 v the control group. (B) Effects of glucuronic acid (25 mg/l) and galacturonic acid (25 mg/l) on the percentage of cavitated mucus cells. The colonic mucosa was analysed by light microscopy as described in Materials and methods. Some 500–1000 stained mucus cells per slide were evaluated for the presence of apical membrane cavitation. A mucus cell with clear apical indentation into the intracellular store was taken as showing signs of cavitation. Data are mean (SE). *p<0.05 compared with the control group.
RELEASE OF COLONIC MUCIN BY FACTORS PRODUCED BY THE COLONIC MICROFLORA
Short chain fatty acids
Luminal administration ofn-butyrate (5 mM) provoked a significant mucin discharge (338.1 (63.6) μg/mg DNA, n = 9, p<0.05v 144.7 (13.5) μg/mg DNA for the control group, n = 9, p<0.05). Increasing the concentration ofn-butyrate to 100 mM decreased the mucus response (fig 4A). Compared with control loops, the mucus cells of colonic preparations exposed to 5 mM butyrate for 30 minutes exhibited deep apical membrane cavitation (fig 2 and fig 4B). In contrast, luminal administration of 5 mM n-butyrate failed to elicit any significant modification in the number of stained mucus cells per crypt (table 3).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Effects of increasing the concentration of butyrate on colonic mucin secretion as evaluated by using a sandwich enzyme linked immunosorbent assay and by histological analysis. Data are mean (SE). *p<0.05 v the control group.
Quantitative histological examination and ELISA of isolated vascularly perfused rat colon after luminal administration of short chain fatty acids
As shown in table 3, administration of 5 mM acetate was without effect on mucus discharge whereas mucus secretion was observed when 100 mM acetate was administered. On administration of 100 mM acetate, an increase in the number of mucus cells with apical cavitation was observed (table 3). Neither 5 nor 100 mM propionate elicited any significant discharge of mucus (table 3).
Sulphur compounds
Mercaptides (sodium hydrogen sulphide) and mercapto fatty acids (mercaptoacetate) are readily produced by colonic bacteria from sulphated polysaccharides, cruciferous vegetables, sulphates, and dietary amino acids.15 Luminal administration of sodium hydrogen sulphide (40 mM) or sodium mercaptoacetate (40 mM) in the isolated vascularly perfused rat colon elicited only a modest increase in mucus secretion (table 4). Table 4 shows total luminal contents in response to sulphur compounds. Sodium hydrogen sulphide produced a twofold increase in total luminal content recovered from the lumen, although luminal DNA content was similar to that observed in the control group (data not shown).
Mucin and total fluid luminal content after administration of reducing sulphur compounds normally present in colonic luminal content
EFFECT OF DIETARY PHENOLIC COMPOUNDS ON MUCIN DISCHARGE
Numerous polyphenols are widely distributed in higher plants and are ingested by humans with regular food. These include trihydroxystilbene (resveratrol) and flavonoids (epicatechin, quercetin).26 Resveratrol, epicatechin, and quercetin at concentrations up to 20 mM were all unable to induce any significant increase in colonic mucus secretion (120.5 (14.7) μg/mg DNA, n = 5, 112.9 (7.5) μg/mg DNA, n = 7, and 135.4 (15.1) μg/mg DNA, n = 5 respectively v 110.1 (17.2) μg/mg DNA for the control group, n = 6).
Discussion
The mechanisms responsible for the secretion of colonic mucins are still unclear. In particular, little is known about the discharge of mucin in response to luminal factors. The preparation of isolated vascularly perfused rat colon was then used as a relevant model to analyse the changes in colonic mucin secretion in response to dietary compounds (algal fibres, dietary fibres, short chain fatty acids, sulphides, phenolic compounds).
Previous investigations suggested that dietary supplementation with fibres may modify the composition of mucin27 or the number of goblet cells.28 Such hypotheses received support from in vivo studies in various animals. Indeed, Vahouny and colleagues7 and Schneeman and colleagues6observed that a wheat bran-containing diet increased incorporation of [3H]glucose and [35S]sulphate into rat intestinal glycoproteins. A morphometric study by Lundin and colleagues28 also suggested that fibre feeding raised the number of goblet cells in the small intestine of Syrian Golden Hamsters. High fibre diets also induced a rise in the number of goblet cells in rat colon.29 30 Furthermore, dietary supplementation with 10% psyllium, a highly branched arabino-xylan polysaccharide used as a fibre supplement, evoked an increase in luminal mucin levels in rat colon, suggesting a higher goblet cell secretory activity.9 In contrast, the present data show that pectin and cellulose did not induce a short term effect on mucin discharge.
An attractive hypothesis that could explain these discrepancies is that dietary fibres induce colonic mucus discharge in vivo through an indirect mechanism that is not present in our model. This mechanism could involve short chain fatty acids, which are formed in the colon by bacterial fermentation of dietary fibre.12 We therefore tested the three major short chain fatty acids (acetate, propionate, and butyrate) present in the colon. We found for the first time that a physiological concentration of butyrate (5 mM) or acetate (100 mM) caused a threefold increase in mucus secretion in the lumen of isolated perfused rat colon, suggesting that fermentable dietary fibres may stimulate colonic mucin secretion in vivo through the production of short chain fatty acids. Moreover, Finnie and colleagues31 showed that sodium butyrate induces a striking increase in mucin synthesis by human colonic biopsy specimens. These observations suggest that short chain fatty acids are influential in increasing the synthesis and secretion of colonic mucin. If this hypothesis adequately explains modulation of mucus secretion by dietary fibres, other mechanisms are also possible. These may include physical characteristics of colonic content, viscosity properties, modification of nutrient absorption, and indirect effects through hormonal regulation.
In this study, 5 mM butyrate produced colonic mucus secretion whereas the highest concentrations of butyrate (100 mM) resulted in a lower mucus response. This result agrees with earlier observations showing that the maximal effect of butyrate on the colonic secretion of the hormonal peptide PYY was observed around 5–10 mM.32 33On the basis of these data, it may be speculated that 100 mM butyrate induces the secretion of some inhibitory factors of colonic secretions. Somatostatin appears to be a good candidate, as endocrine D cells are scattered throughout the epithelium of distal bowel and it inhibits exocrine and endocrine secretions in the gut.34 Additional experiments with the isolated perfused rat colon are required to validate this hypothesis.
In response to stimulation, intestinal goblet cells may accelerate their discharge of mucin by way of two processes. In many mucus cells, a phenomenon of compound exocytosis is induced, resulting in deep cavitation of the apical membrane surface of mucus cells.1 25 Some intestinal mucus cells can also respond to stimulation by another process that decreases the intracellular store of mucus granules but without cavitation.1 25 When the latter mechanism leads to total depletion of mucus granules, goblet cells are no longer stained by alcian blue/periodic acid-Schiff and appear indistinguishable from other epithelial cells. This study shows for the first time that luminal acetate and butyrate induce mucus discharge by compound exocytosis. Supporting this hypothesis, the results of Sakata and Engelhardt24 and ours showed that short chain fatty acids did not significantly influence the number of mucus cells staining for mucin.
Our interest focused then on algal polysaccharides because of the widespread use of some of these fibres (alginate, carrageenans) by food manufacturers as gelling agents, thickeners, stabilisers, or emulsifiers.10 The doses (1–25 mg/l) of algal fibres used in this study were chosen to approximate the estimated human intake as food additive.35 36 The present data show for the first time that alginate (25 mg/l) appreciably increased the discharge of colonic mucin. Interestingly, the magnitude of mucus release stimulated by alginate was only threefold less than that stimulated by bethanechol (a cholinergic agonist) or the neuropeptide bombesin, which were the most powerful agents used in our earlier study.21Together, these data indicate that alginate is an effective secretagogue of colonic mucus, suggesting its potential therapeutic usefulness in dietary supplementation. However, the physiological consequences of alginate ingestion are far from being understood and may require in particular long term studies. Indeed, changes in dietary habits may modify the chemical composition and functional characteristics of mucins.5 7 37
Ulvan, a fibre extracted from green seaweed, has a high content of sulphate and glucuronate moieties.10 Our results establish for the first time that ulvan induces mucin secretion. Interestingly, glucuronic acid was also found to stimulate mucus discharge, suggesting that this substrate may be essential for mucus secretory activity. These studies also show that two other sulphated polysaccharides (carrageenin and fucoidan) did not induce a significant increase in mucus secretion. The presence of sulphated groups thus does not seem to influence colonic mucin discharge significantly.
A consequence of the presence of sulphated polysaccharides is the production of sulphide in the colonic lumen.16 Luminal concentrations of sulphides in humans vary between 0.1 and 10 mM.15 In this study, we found that supraphysiological doses of two sulphides did not stimulate mucin discharge. Nonetheless, it should be pointed out that sodium hydrogen sulphide provoked a sharp increase in luminal fluid accumulation whereas sodium mercaptoacetate was without effect. These results agree with early observations that hydrogen sulphide, but not mercaptoacetate, inhibited sodium transport in the colonic mucosa.38
In conclusion, we have provided evidence that the preparation of isolated vascularly perfused rat colon offers a unique ex vivo model for assessing the quantitative changes in mucin secretory activity of colonic goblet cells in response to well defined luminal factors. This approach has enabled us to show that some dietary fibres (alginate, ulvan), uronic acids (glucuronic acid, galacturonic acid), and short chain fatty acids may be useful for stimulating colonic mucus secretion. Our findings could be of considerable interest in gastrointestinal physiology and nutrition. They need to be supported by additional studies with in vivo and in vitro models to validate the hypothesis that some dietary fibres and short chain fatty acids may be protective against various intestinal diseases.