Article Text

Abstract
BACKGROUND The pro-mutagenic lesionO 6-methyldeoxyguanosine (O 6-MedG), a marker of exposure to many N-nitroso compounds (NOC), can be detected in normal and tumour DNA isolated from colorectal tissue. The biological significance of this exposure is, as yet, unknown but in situ NOC formation is bacterially catalysed suggesting that NOC formation and potentially DNA alkylation will vary throughout the large bowel.
AIMS To determine ifO 6-MedG levels in colorectal DNA vary within the large bowel.
PATIENTS We studied 62 men and women undergoing surgery for colorectal tumours in the north west of England.
METHODS O 6-MedG levels were measured in paired normal and tumour DNA samples. DNA was digested to nucleosides, fractionated by HPLC, and purified O 6-MedG quantified by a radioimmunoassay.
RESULTS O 6-MedG was detected in 27 out of a total of 62 (43%) normal DNA samples and in 30 of 58 (52%) tumour DNA samples: it was present at concentrations of <0.01–0.94 and <0.01–0.151 μmolO 6-MedG/mol deoxyguanosine for normal and tumour DNA, respectively. Levels ofO 6-MedG in normal, but not tumour, DNA from the proximal colon were lower than those found in DNA from either the sigmoid colon (p=0.03) or rectum (p=0.05). When the analysis was restricted to samples that containedO 6-MedG, similar results were obtained in that O 6-MedG levels in normal DNA were lower in the proximal colon than in the sigmoid colon (p=0.04) or rectum (p=0.03).
CONCLUSIONS DNA alkylation varied within the large bowel possibly due to in situ NOC formation and was highest in areas of the colon and rectum where the highest incidence of large bowel tumours occurs, suggesting that DNA alkylation may play a role in the aetiology of colorectal cancer.
- O6-methylguanine
- alkylating agents
- colorectal cancer
- DNA adducts
Abbreviations used in this paper
- O6-MedG, O6-methyldeoxyguanosine
- dG, deoxyguanosine
- NOC
- N-nitroso compounds
Statistics from Altmetric.com
Colon cancer is one of the most common cancers in the developed world and colon carcinogenesis is a multistep process involving both genetic and environmental factors.1 Major advances have resulted in the identification of cancer susceptibility genes and in delineating the genetic changes that accumulate during induction of sporadic colon cancers.2-4 In contrast, our knowledge of the environmental factors involved is still relatively limited. Numerous epidemiological studies indicate that dietary factors can modify the risk of colon cancer.5 For example, vegetable consumption and fibre intake (possibly by reducing the formation of carcinogens or inhibiting cell replication) are inversely associated with risk, whereas meat protein and fat are consistently and positively associated with risk, possibly by being a source of chemical carcinogens.5
The link between meat protein intake and colorectal cancer risk is usually considered to arise through consumption of heterocyclic amines.6 However, intake of meat protein may also increase the formation of N-nitroso compounds (NOC) in the human colon.7 We have previously shown, in two separate studies, that the pro-mutagenic lesion O 6-methyldeoxyguanosine (O 6-MedG), a marker of exposure to many NOC, can be detected in normal and tumour DNA isolated from individuals with colorectal cancer.8 ,9 As NOC formation is bacterially catalysed,10 ,11 NOC formation within the large bowel will vary and this may alterO 6-MedG levels throughout the large bowel. Hence we have quantifiedO 6-MedG levels in a cross sectional study of patients with colorectal tumours to examine if DNA alkylation varies throughout the large bowel: previously, we reported on the association of K-ras mutations and the frequency of DNA alkylation in this population.9
Methods
SUBJECTS
Paired normal (macroscopically normal tissue taken ∼5 cm from the tumour edge) and tumour tissues were taken from 62 individuals (30 men and 32 women: median age 73 years) presenting with colorectal tumours at hospitals within the Manchester area. Sixteen of these individuals had proximal colon tumours (tumours of the caecum (n=13), transverse colon (n=2), and hepatic flexure (n=1)), 21 had sigmoid colon tumours, and 25 rectal tumours. There were no significant age or sex differences between these groups (data not shown).
QUANTITATION OF O6-METHYLDEOXYGUANOSINE BY RADIOIMMUNOASSAY
DNA was extracted from the tissue samples and enzymatically digested to nucleosides.12 The digest was fractionated by Aminex A7 cation exchange chromatography and theO 6-MedG containing fractions were analysed in two independent radioimmunoassays using mouse monoclonal α-O 6-MedG.12 ,13Only samples with ⩾5% inhibition of tracer antibody binding in both assays were considered positive, as this is the lowest percentage inhibition that is statistically significantly different from zero. The lower limit of detection was ∼0.01 μmolO 6-MedG/mol deoxyguanosine (dG). Sufficient DNA was obtained for analysis ofO 6-MedG by radioimmunoassay in all but four tumour samples.
DATA ANALYSIS
Univariate analysis was carried out for: (i) alkylation frequency using the χ2 test; and (ii) adduct levels using non-parametric methods (Mann-Whitney U test and Kruskal-Wallis one way analysis of variance) as adduct levels were not normally distributed. Adduct data were analysed: (a) using all values for adduct levels, assigning a value of 0.005 μmolO 6-MedG/mol dG (that is, half the assay detection limit) to samples in whichO 6-MedG was not detected; and (b) using only values for whichO 6-MedG was ⩾0.01 μmolO 6-MedG/mol dG (that is, detected). Data in table 1 are presented as mean (SD)O 6-MedG levels for descriptive purposes only. Significance testing was carried out without adjustment for multiple comparisons.
Frequency of alkylation (number (%)) and O6-methylguanine concentrations (mean (SD)) in DNA isolated from macroscopically normal and malignant tissue samples from the colon and rectum
Results
O 6-MedG was detected in 27 out of a total of 62 (43%) normal DNA samples and in 30 of 58 (52%) tumour DNA samples: it was present at concentrations of <0.01–0.94 and <0.01–0.151 μmolO 6-MedG/mol dG for normal and tumour DNA, respectively. The highest value (0.94 μmolO 6-MedG/mol dG) was found in normal DNA from a patient with a sigmoid colon tumour (the positive outlier in table 1 and fig 1). In normal tissue DNA, the concentration of O 6-MedG in the proximal colon (that is, the caecum, transverse colon, and hepatic flexure) was lower than that found in the sigmoid colon (p=0.03) or rectum (p=0.05) when either all samples (fig 1) or only samples containing detectable levels of O 6-MedG were examined (p=0.04 and 0.03 for the sigmoid colon and rectum, respectively) (table 1). Furthermore, the alkylation frequency (25%) in normal DNA from individuals with proximal colon tumours was approximately half that found in individuals with sigmoid colon (52%, p=0.18) or rectal (48%, p=0.25) tumours (table 1). However, in tumour tissue DNA there were no differences in O 6-MedG levels between the proximal colon, sigmoid colon, and rectum when either all samples (fig 1) or only those samples with detectableO 6-MedG levels (table 1) were analysed. The frequency of DNA alkylation in tumour DNA was highest in the proximal colon (60%) and rectum (63%; p=0.80v proximal colon) and lowest in the sigmoid colon (32%; p=0.19 v proximal colon) (table1).
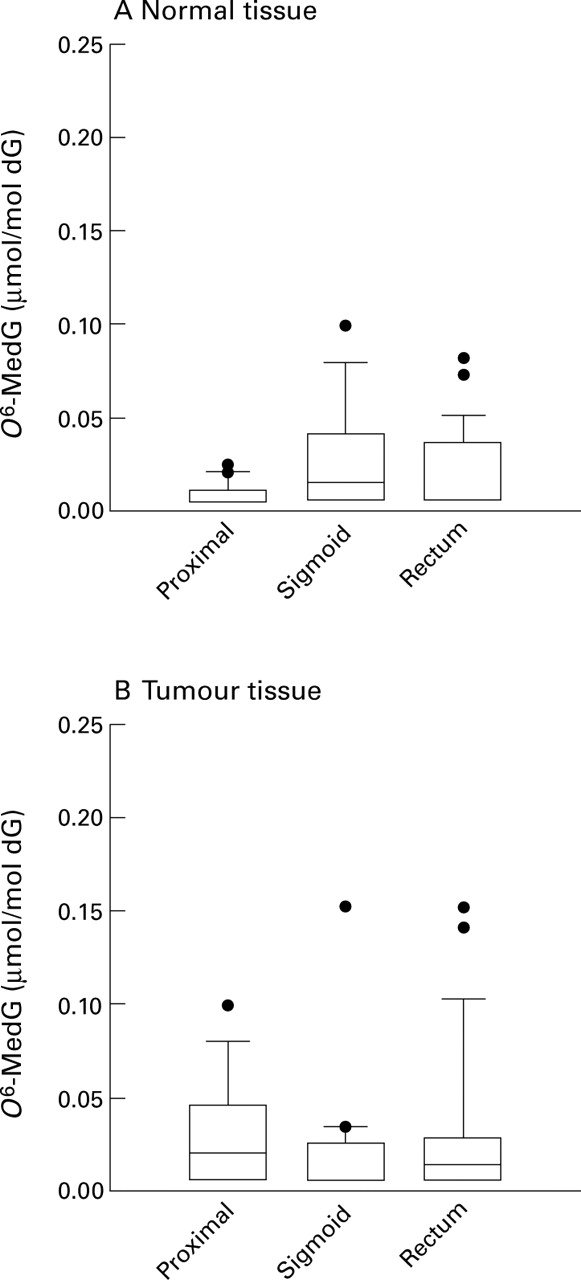
Box and whisker plots showing O6-methyldeoxyguanosine (O6-MedG) concentrations in DNA from individuals with colorectal cancer. (A) Normal tissue from individuals with colorectal tumours. (B) Tumour tissue from individuals with colorectal tumours. Median concentration is denoted by the line in the box (which if not present means that most DNA samples had no detectable O6-MedG), 25–75 percentiles as the box, 10–90% percentiles as capped vertical bars, and outliers as dots. One outlier (0.94 μmol O6-MedG/mol dG for normal DNA from an individual with sigmoid colon cancer) is not shown in (A) and (B) for ease of comparison. O6-MedG concentrations in normal DNA from the proximal colon (that is, the caecum, transverse colon, and hepatic flexure) are lower than those found in DNA from the sigmoid colon (p=0.03, Mann-Whitney U test) and rectum (p=0.05). Exclusion of the sigmoid colon outlier does not alter the comparison between the proximal colon (p=0.05, Mann-Whitney U test)
In the proximal colon, the alkylation frequency in tumour DNA samples (60%) was higher than that in DNA from normal tissue (25%; p =0.11; table 1) and O 6-MedG levels in tumour DNA were higher than levels in normal DNA (p<0.05, Wilcoxon matched pairs test). In contrast, there were no differences between the alkylation frequency and O 6-MedG levels in normal and tumour DNA from individuals with sigmoid colon or rectal tumours (table 1).
In those individuals withO 6-MedG in both normal and tumour DNA, there was no association between levels ofO 6-MedG in normal DNA and those in tumour DNA from the same individual (data not shown). The ratio ofO 6-MedG in normal DNA to that in tumour DNA was 0.53 in the caecum (n=2), 0.96 in the sigmoid colon (n=4), and 1.95 in the rectum (n=10; p=0.19 for differences in the ratios using the Kruskal-Wallis test). Twenty individuals had no detectable O 6-MedG levels in either normal (n) or tumour (t) DNA (n−t−) whereas 16 individuals had O 6-MedG present in both DNA samples (n+t+). In a further eight individuals,O 6-MedG was detected only in normal DNA (n+t−) whereas in 14 individualsO 6-MedG was detected only in tumour DNA (n−t+). The variation in proportions of n-t-, n+t-, n-t+, and n+t+ individuals with tumour site is shown in fig 2. The proportion of individuals with both samples alkylated increased from 13% in the proximal colon to 21% in the sigmoid colon to 42% in the rectum (p=0.05, χ2 test for trend).

{kind=link}
{kind=link}
Relationship between tumour site and frequency of alkylation in paired normal and tumour DNA samples. Normal (n) and tumour (t) DNA samples containing detectable levels of O6-methyldeoxyguanosine (O6-MedG) are identified as n+ and t+, respectively, whereas those with no detectable levels of O6-MedG are identified as n− or t−. The proportion of individuals with both DNA samples alkylated increased from 13% for individuals with proximal colon tumours to 42% for those with rectal tumours (p=0.05, χ2 for linear trend).
Discussion
Previous studies have shown that DNA alkylation is a common event in colorectal tissue but the source of exposure to the methylating agents remains to be identified. These results confirm our previous study in a separate population of patients with colorectal disease,8 in demonstrating that colorectal DNA, obtained from individuals with malignant colorectal disease, contains detectable amounts of the pro-mutagenic and pro-carcinogenic lesionO 6-MedG. Furthermore, evidence from the current study indicates thatO 6-MedG levels vary within the large bowel as (i) DNA alkylation was higher in normal tissue from the sigmoid colon and rectum than in the proximal colon, (ii) only 13% of paired normal and tumour DNA samples obtained from the proximal colon were both alkylated compared with 21% in the sigmoid colon and 42% in the rectum, and (iii) O 6-MedG levels in tumour DNA from the proximal colon (but not the sigmoid colon or rectum) were greater than those in normal DNA.
A number of different mechanisms acting either singly or in combination could account for this variation inO 6-MedG levels within the colon and rectum. Firstly, the methylating agent may arise from bacterial catalysed N-nitrosation reactions within the large bowel as it is known that (i) bacterial composition and their metabolic capacity varies significantly within the intestinal tract14 ,15 and (ii) the lumen of the large bowel contains high concentrations of nitrogen containing compounds16 and is therefore a site for nitrosation reactions, presumably catalysed by resident bacterial flora.10 This has been shown to lead to the formation of large quantities of apparent totalN-nitroso compounds but not volatile nitrosamines such asN-nitrosodimethylamine.7 ,10 ,11It is possible therefore, that these reactions could lead to the in situ formation of a methylating agent within the lower portion of the bowel and hence to increased DNA damage at the sites of formation.
Alternatively, differences in the formation of methylating agents within the colon and rectum may arise through differential expression of carcinogen metabolising enzymes,17 perhaps through dietary induced variations in gene expression.18 Such differences may lead to increased formation or detoxification of the methylating agent(s), thereby leading to differences in adduct levels. In addition, although differences in the levels ofO 6-alkylguanine DNA alkyltransferase within the colon could also account for variations inO 6-medG levels, no association between levels of the repair protein and adduct was observed (data not shown), suggesting that in this case changes in repair capacity were not an important factor in explaining the observed differential. Finally, the increased alkylation in the lower bowel may result simply from increased time of contact (and not formation) with lumenally borne genotoxic agents such as N-nitrosated bile acids.19
Our results also tend to indicate that factors affecting DNA alkylation in tumour tissue vary from those affecting alkylation in normal tissue. There was no gradient inO 6-MedG levels in tumour DNA from the proximal colon to the rectum. Such discrepancies may result from differences in carcinogen metabolising enzymes between normal and tumour tissue.20-22 At one level, this might enable the differential targeting of tumours within the colon by alkylating agents or alternatively lead to the development of agents that reduce exposure specifically to normal tissue: it also means that measurements in normal tissue DNA do not reflect what is happening in tumour tissue DNA.
The biological significance of these observed levels of DNA alkylation in the human colon remains to be determined, particularly as concentrations and frequencies of alkylation are similar in this population to those observed in other tissues from populations with differing and greater cancer risks. However, it is of interest thatO 6-MedG levels were highest in the sigmoid colon and rectum—that is, the sites where most sporadic large bowel tumours occur.1 Such an association, however, may be tenuous, particularly as we measured the adduct in a mixed population of cells from different locations in different individuals and also, these individuals had a tumour which potentially may affect the biomarker. Furthermore, the adduct measurement probably reflects recent/current exposure which may bear no relationship to exposure in the aetiologically relevant time frame. However, in experimental studies, colon tumours induced by dimethylhydrazine in rats occurred in the distal portion of the colon, precisely in the region where DNA adducts are highest (Jackson et al, unpublished work).
There is further evidence to suggest that methylating agent exposure may be important. Vegetable and micronutrient intake, which reduce the risk of large bowel cancer, also reduce endogenous NOC formation and NOC tumorigenicity.23 ,24 Large bowel tumours contain mutational events (for example,K-ras GC-AT transitions and mismatch repair defects25-29) that have been shown to occur in vitro and in vivo following exposure to alkylating agents.30-32Moreover, we have found evidence to indicate that the DNA repair protein, O 6-alkylguanine DNA alkyltransferase, provides protection against the mutagenic effects of alkylating agents in the lower intestinal tract as individuals with lowO 6-alkylguanine DNA alkyltransferase activity were at increased risk of having a GC-AT transition K-ras mutation but not a transversion mutation.33 This result is entirely consistent with the known action of the repair protein in (i) removingO 6-MedG lesions from DNA34 and (ii) reducing the frequency ofK-ras GC-AT transitions in an experimental model of large bowel cancer,35 thereby suggesting a role for methylating agents in the aetiology of these cancers.
Acknowledgments
The work presented in this study was supported by the Cancer Research Campaign and in part by the Egyptian Government (Cultural and Educational Bureau).
Abbreviations used in this paper
- O6-MedG, O6-methyldeoxyguanosine
- dG, deoxyguanosine
- NOC
- N-nitroso compounds