Article Text

Abstract
BACKGROUND The enteric nervous system is important in the pathophysiology of intestinal fluid secretion induced by cholera toxin (CT),Escherichia coli heat labile (LT), and heat stable (STa) toxins. The neurotransmitters involved are not fully elucidated. Vasoactive intestinal polypeptide (VIP), a potent intestinal secretagogue present in the enteric nervous system, is increased after exposure of the cat intestine to CT. Whether VIP is involved in the pathogenesis of cholera and other toxins in not known.
AIM To study in vivo the effect of VIP antagonism on jejunal fluid secretion induced by CT, LT, and STa.
METHODS CT, LT (25 μg), or 0.9% NaCl was instilled in an isolated 25 cm segment of rat jejunum, and the VIP antagonist (VIPa) [4Cl-D-Phe6, Leu17]-VIP (0.2 or 2 μg/kg/min) or 0.9% NaCl was given intravenously. Two hours later, single pass in vivo jejunal perfusion was performed to assess fluid movement. In STa experiments, intravenous VIPa or 0.9% NaCl was given and 30 minutes later the jejunal segment was perfused with a solution containing STa 200 μg/l.
RESULTS VIPa had no effect on basal intestinal fluid absorption. CT induced net fluid secretion (median −68 μl/min/g dry intestinal weight (interquartile range −80 to −56)) which was dose dependently reversed by VIPa (6.2 (−16 to 34) and 29 (17 to 42); p<0.01). Similarly, LT induced secretion (−63 (−73 to −30)) was attenuated by VIPa (0.2 μg/kg/min) (−15 (−24 to −1); p<0.01) and totally reversed to normal levels by VIPa (2 μg/kg/min) (37 (28–56); p<0.01 compared with LT and not significant compared with normal controls). STa induced secretion (−17 (−19 to −2)) was also reversed by VIPa (12 (9–23) and 14 (0–26); p<0.01).
CONCLUSION VIP plays an important role in CT, LT, and STa induced intestinal secretion and may be the final putative neurotransmitter in the pathophysiology of these toxins.
- cholera toxin; Escherichia colitoxins
- intestinal secretion
- vasoactive intestinal polypeptide
- enteric nervous system
Abbreviations used in this paper
- CT
- cholera toxin
- LT
- Escherichia coli heat labile toxin
- STa
- Escherichia heat stable toxin
- VIP
- vasoactive intestinal polypeptide
- VIPa
- vasoactive intestinal polypeptide antagonist
- 5-HT
- 5-hydroxytryptamine
- PES
- plasma electrolyte solution
- PEG
- polyethylene glycol
Statistics from Altmetric.com
- cholera toxin; Escherichia colitoxins
- intestinal secretion
- vasoactive intestinal polypeptide
- enteric nervous system
Intestinal secretion induced by cholera toxin (CT) and the structurally related enterotoxin, Escherichia coli heat labile toxin (LT), has been attributed solely to direct activation of adenylate cyclase with a corresponding increase in intracellular cAMP. However, evidence has been accumulating on the importance of other mediators such as 5-hydroxytryptamine (5-HT), prostaglandins, neurotensin, and of the enteric nervous system in the pathophysiology of CT induced secretion.1-4 In addition, vasoactive intestinal polypeptide (VIP) has been found to be released in response to administration of CT.5 ,6 VIP is present in high concentrations in the subepithelial neural plexuses and is a potent secretagogue which acts via specific adenylate cyclase linked receptors.7 For these reasons, it seems likely that VIP plays an important physiological and pathophysiological role in the intestine. The proposed mechanism for CT induced secretory diarrhoea includes a neural reflex with a common final pathway through VIP to the epithelial cells.8 However, this proposal remains to be confirmed in studies proving that VIP is important in the pathophysiology of CT induced secretion and is not just an innocent bystander.
The pathophysiology of diarrhoea caused by other enterotoxins has not been as extensively investigated. LT, which is structurally similar to CT, binds to the same receptor on enterocytes (GM1 ganglioside) and has been shown to stimulate adenylate cyclase with a corresponding increase in cAMP production.9 ,10 However, the two toxins are not identical in their amino acid composition9-12 and their binding affinity is different.11 ,13 In addition, we have previously demonstrated that 5-HT plays an important role in CT but not LT induced secretion suggesting that there are fundamental differences in the pathophysiology of fluid secretion caused by these toxins.14 ,15 Furthermore, Nzegwu and Levin have shown that intraluminal capsaicin inhibits LT induced secretion implying that LT acts through afferent nerve fibres.16 The neurotransmitters involved in the efferent loop of this neural reflex are not known. Whether VIP plays a role in LT induced secretion has not been previously investigated.
The role of cGMP in Escherichia coli heat stable toxin (STa) induced intestinal secretion is well established.17-19 It has been shown that, in common with CT, the enteric nervous system is involved in STa induced secretion as demonstrated by the inhibitory effect of hexamethonium, lidocaine, tetrodotoxin, and capsaicin.20-23 Furthermore, we have previously shown a differential effect of 5-HT antagonism on CT and STa induced secretion and demonstrated that 5-HT, unlike in CT, is not involved in STa secretion.14 It would be interesting to know whether the neurotransmitter VIP plays a role in this domain.
Thus the aim of our study was to examine the importance of VIP in the pathophysiology of CT, LT, and STa by examining the effect of VIP antagonism on intestinal secretion induced by these toxins.
Materials and methods
CT AND LT EXPERIMENTS
Adult Sprague-Dawley rats (180–220 g body weight) fasted for 18 hours were anaesthetised with intraperitoneal injection of sodium pentobarbitone (60 mg/kg) and maintained throughout the experiments by interval intraperitoneal injections (15–30 mg/kg) as necessary. The abdomen was opened through a midline incision and cannulae inserted into the small intestine proximally (5 cm distal to the duodenojejunal junction) and 25 cm distally in the jejunum and fixed by ligation as previously described.24 CT, LT (25 μg in 3 ml of 0.9% NaCl), or 3 ml of 0.9% NaCl alone (controls) was instilled in the isolated segments followed by clamping of both the proximal and distal cannulae. The intestine was returned to the abdominal cavity and the abdomen closed. After two hours the clamps were removed and the intestine perfused at a rate of 0.5 ml/min with a plasma electrolyte solution (PES) containing Na 140, K 4, Cl 104, and HCO3 40 mmol/l to which 5 g of cold polyethylene glycol 4000 (PEG) and 4 μCi/l of [14C]-PEG were added. Thirty minutes were allowed to elapse to ensure establishment of a steady state following which consecutive 10 minute collections of the effluent were obtained from the distal cannula for one hour. All rats had femoral vein cannulation and an infusion of the VIP antagonist [4Cl-D-Phe6,Leu17]VIP25 ,26 at a dose of 0.2 (VIPa0.2) or 2 μg/kg/min (VIPa2), or 0.9% NaCl was started directly after instillation of CT or LT. At the end of the experiments, the animals were killed by an overdose of pentobarbitone and the perfused intestinal segment removed, rinsed, blotted, and desiccated in an oven at 100°C to obtain the dry weight. The samples of effluent were analysed immediately or kept frozen at −20°C and analysed within two weeks.
STa EXPERIMENTS
In experiments with Escherichia coliheat stable toxin (STa), a 25 cm segment of jejunum starting 5 cm distal to the duodenojejunal junction was perfused with PES containing [14C]-PEG to which 200 μg/l of ST (equivalent to 50 000 mouse units)27 were added. After 30 minutes of perfusion to establish a steady state, consecutive 10 minute collections of the effluent were obtained for one hour. Infusion of the VIP antagonist or 0.9% NaCl was started 30 minutes before the perfusion.
VIP EXPERIMENTS
In a different groups of animals, both femoral veins were cannulated. VIPa infusion (0.2 or 2 μg/kg/min) was started in one femoral vein and 30 minutes later VIP infusion (0.02 or 0.2 μg/kg/min) was stared in the second femoral vein. One hour later, intestinal perfusion was performed as mentioned above to calculate fluid and electrolyte movement.
In a different group of animals, the effect of VIPa (2 μg/kg/min) on intestinal secretion induced by 1 mM carbachol added to the perfusate was tested.
Steady state conditions in all experiments were demonstrated by less than 5% variation in fluid movement between consecutive 10 minute collections. Also, values were accepted only if recovery of radioactive PEG was 95–105%.24
ANALYTICAL METHODS
[14C]-PEG concentrations in the effluent were measured in duplicate by liquid scintillation spectroscopy in an LKB Wallac 1409 scintillation counter. Sodium and chloride concentrations were determined by an Hitachi 911 analyser.
The mean of net fluid and solute movement of six consecutive effluent samples were calculated and expressed, respectively, as μl/min/g and μmol/min/g of dry intestinal weight. Positive values denote net absorption and negative values net secretion.
STATISTICS
Results are expressed as median (interquartile range) in each group of animals studied. Differences in fluid and solute movement between pairs were tested using the Wilcoxon rank sum test.
MATERIALS
Cholera toxin, Escherichia coli heat stable and heat labile toxins, VIP, the VIP antagonist, and carbachol were obtained from Sigma Chemical Company (St Louis, Missouri, USA). Radiolabelled polyethylene glycol ([14C]-PEG 4000) was obtained from Amersham International (Buckinghamshire, UK) and all other chemicals were supplied by British Drug House (BDH Chemicals, Poole, UK).
Results
Basal fluid absorption (median 61 μl/min/g (interquartile range 49–81); n=11) was not affected by the VIP antagonist at 0.2 or 2 μg/kg/min (48 (28–79), n=7, and 55 (34–75), n=6, respectively). Chloride and sodium movement paralleled that of fluid (table1).
Effect of vasoactive intestinal polypeptide (VIP) antagonism on net Na+ and Cl− flux in normal controls, in cholera toxin (CT), Escherichia coli heat labile toxin (LT), and Escherichia heat stable toxin (STa) induced secretion, and on VIP induced decrease in absorption
CT AND LT EXPERIMENTS
Net fluid secretion occurred in all animals who received CT (−68 μl/min/g (−80 to −56), n=7) (fig 1). The VIP antagonist at a dose of 0.2 μg/kg/min reversed net fluid secretion to absorption (6.2 (−16 to 34), n=7; p<0.01), and at 2 μg/kg/min further increased fluid absorption (29 (17–42), n=7; p<0.01 compared with CT alone, p<0.01 compared with normal controls, and not significant compared with VIPa0.2). Chloride and sodium secretion were also significantly reduced in a dose dependent manner by the VIP antagonist (table1).

Effect of the VIP antagonist [4Cl-D-Phe6, Leu17 ]VIP given intravenously at a dose of 0.2 and 2 μg/kg/min (VIPa0.2 and VIPa2, respectively) on cholera toxin (CT) induced secretion. Results are expressed as median (interquartile range); positive values denote absorption and negative values denote secretion. **p<0.01 compared with controls; ††p<0.01 compared with CT and controls.
LT caused marked fluid secretion (−63 (−73 to −30), n=6) which was significantly attenuated by VIPa 0.2 μg/kg/min (−15 (−24 to −1), n=7; p<0.01). Fluid secretion was completely prevented by VIPa 2 μg/kg/min (37 (28–56), n=6; p<0.01 compared with LT alone, and not significant compared with normal controls) (fig2).

Effect of the VIP antagonist [4Cl-D-Phe6, Leu17 ]VIP given intravenously at a dose of 0.2 and 2 μg/kg/min (VIPa0.2 and VIPa2, respectively) on Escherichia coli heat labile toxin (LT) induced secretion. Results are expressed as median (interquartile range); positive values denote absorption and negative values denote secretion. **p<0.01 compared with controls; ††p<0.01 compared with LT and controls; ‡‡p<0.01 compared with LT and p>0.05 compared with controls.
STa EXPERIMENTS
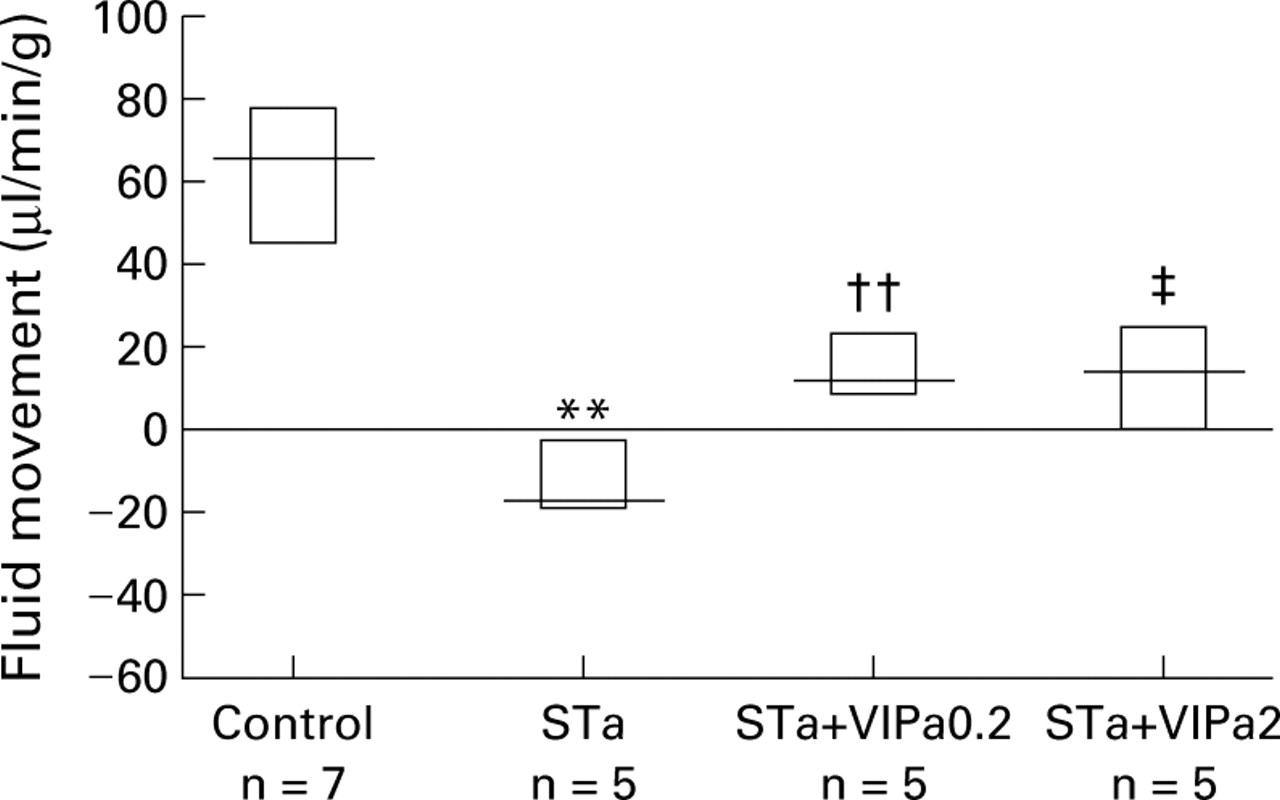
Perfusing the small intestine with a solution containing STa caused marked fluid, chloride, and sodium secretion (−17 (−19 to −2) μl/min/g, −10 (−15 to −5.5), and −7 (−9.2 to −4) μmol/min/g, respectively; n=5) (fig 3, table 1). VIPa0.2 and VIPa2 reversed fluid secretion to absorption which was still less than normal controls (12 (9–23), n=5, p<0.01; and 14 (0–26), n=5, p<0.05, respectively). Sodium and chloride movement paralleled that of fluid (table 1).

Effect of the VIP antagonist [4Cl-D-Phe6, Leu17 ]VIP given intravenously at a dose of 0.2 and 2 μg/kg/min (VIPa0.2 and VIPa2, respectively) on Escherichia coli heat stable toxin (STa) induced secretion. Results are expressed as median (interquartile range); positive values denote absorption and negative values denote secretion. **p<0.01 compared with controls; ††p<0.01 compared with STa and controls; ‡p<0.05 compared with STa and p<0.01 compared with controls.
VIP EXPERIMENTS
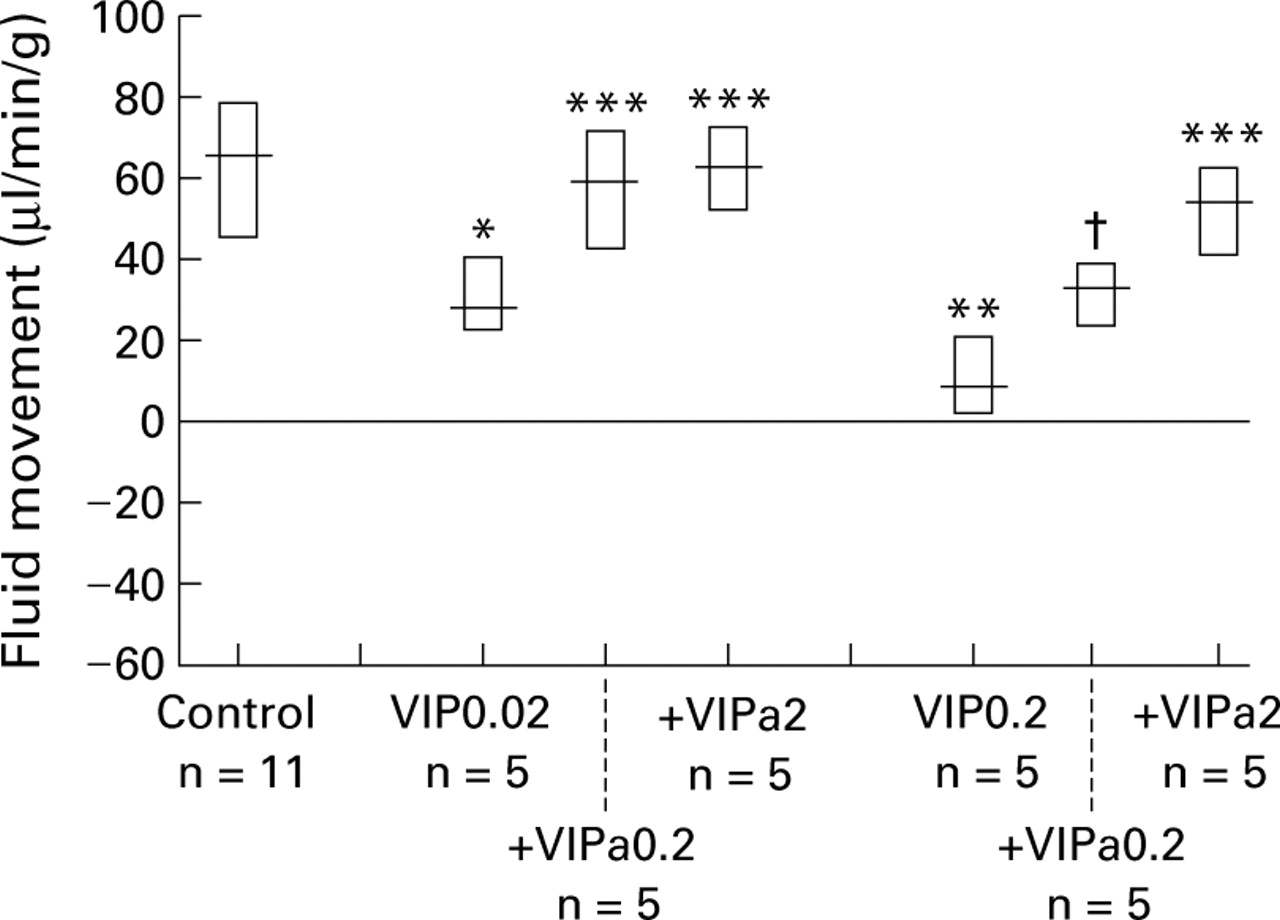
Fluid absorption in normal controls was significantly decreased by intravenous VIP at a dose of 0.02 μg/kg/min (28 (22–40), n=5; p<0.05). VIPa0.2 and VIPa2 completely abolished the effect of VIP (59 (42–71), n=5, and 62 (28–59), n=5). VIP at a dose of 0.2 μg/kg/min caused a further decrease in fluid absorption (8 (2–20), n=5) which was partially attenuated by VIPa0.2 (32 (23–38), n=5; p<0.05 compared with VIP), and completely prevented by VIPa2 (53 (40–61), n=5; p<0.05 compared with VIP and not significant compared with normal controls) (fig 4).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Effect of the VIP antagonist [4Cl-D-Phe6, Leu17 ]VIP given intravenously at a dose of 0.2 and 2 μg/kg/min (VIPa0.2 and VIPa2, respectively) on VIP induced decrease in fluid absorption at 0.02 and 0.2 μg/kg/min given intravenously. Results are expressed as median (interquartile range). *p<0.05, ***p>0.05 and **p<0.01 compared with controls; †p<0.05 compared with VIP0.2 and controls
Carbachol induced fluid secretion (−17 (−19 to −9), n=4) was not affected by VIPa2 (−22 (−25 to −14)).
Discussion
The present experiments demonstrate that VIP plays an important role in intestinal fluid and electrolyte secretion induced by CT andE coli enterotoxins. It has previously been shown that activation of the enteric nervous system is crucial for the action of these three toxins.4 ,15 ,20 ,22 However, how each toxin stimulates the enteric nervous system or what the final mediator(s) of secretion is, is not yet fully understood. It seems that the three toxins do not share the same neuronal pathway activation. Capsaicin afferent fibres have been found to play an important role in LT and STa16 ,23 but not in CT induced secretion28 where the afferent neuronal reflex is activated by 5-HT through neuronal 5-HT3receptors.3 ,14 ,15 In addition, C fibres may be activated through tachykinin NK2 receptors to stimulate secretion in STa29; however, whether or not the same receptors are activated by the other toxins is not known. Furthermore, toxins may evoke release of peptides from endocrine cells which may activate the enteric nervous system via receptors on dendrites located just beneath the epithelium.30 Acetylcholine and VIP have been implicated as putative neurotransmitters in the secretomotor efferent limb of the neuronal reflex.5 ,8 ,31 VIP is a neurotransmitter found in the myenteric plexus, lamina propria, and mucosa of the small and large intestine in different animal species.6 ,32 ,33 Waldman and colleagues34have demonstrated that VIP is released from the intestine in response to secretomotor reflexes, where it acts on the mucosa to activate adenylate cyclase. VIP given intra-arterially or intravenously induced water secretion or reduced water absorption in the small intestine.25 ,35 In addition, the fact that VIP may be implicated in the pathophysiology of intestinal secretion induced by CT came from the following observations. Firstly, VIP is released from the cat small intestine exposed to CT.6 Secondly, inhibition of CT induced secretion by methionine-enkephalin and somatostatin is paralleled by a decrease in VIP release.36 Whether VIP has a direct effect on inducing secretion in CT and in other toxins needs to be confirmed by examining the effect of VIP inhibition on the secretory effect of these toxins. The recent advent of VIP antagonists has permitted a better understanding of the importance of VIP in many physiological and pathophysiological states. While some in vitro studies failed to show these antagonists as competitive inhibitors of VIP,37 ,38 in vivo studies have demonstrated that they can antagonise or reverse the secretory effect of VIP in the intestine.25 ,26 In our study, the validity and specificity of [4Cl-D-Phe6,Leu17]VIP as a VIP antagonist was tested by its effect on reversing the VIP induced decrease in jejunal fluid absorption and by its failure to reverse carbachol induced secretion. Moreover, there are no reports that show that VIPa crosses the blood-brain barrier. Accordingly, we assume that the effect of VIPa is peripheral, although a central effect cannot be excluded.
In cholera, the secretagogue 5-HT has been identified as an indispensable factor in the pathophysiology of small intestinal secretion.2 ,3 ,14 ,15 Thus 5-HT released from enterochromaffin cells activate, among other receptors, neuronal (5-HT3) receptors leading to intestinal secretion.3 ,14 ,15 However, whether or not activation is direct or via release of other neurotransmitters or activation of other neurones is not clear. Neurotransmitters, such as substance P, have been found to be recruited by CT39 implying that 5-HT is not the only transmitter involved in CT. In the present experiments, VIP antagonism dose dependently decreased fluid secretion induced by CT and partially reversed it to absorption, implying that VIP plays an important role in CT induced secretion but some other mediators may also be involved.
LT is a toxin structurally similar to CT and was thought to act via the same pathways. However, recent evidence suggests that these two toxins, although both stimulate adenylate cyclase, have totally different pathophysiological mechanisms. It has been shown definitively that 5-HT is not involved in LT induced secretion.14 ,15 In addition, blocking substance P receptors does not affect LT induced secretion.39 Hence, how does LT stimulate secretion? LT activates intracellular cAMP but this does not explain inhibition of secretion by neuronal blockade. By binding to some receptor sites on enterocytes, it may release an unknown transmitter which activates the neuronal reflex. Alternatively, it has been postulated that the toxin may bind directly to enteric neurones and thus activate secretion. Although the afferent capsaicin sensitive neurones are stimulated during LT exposure,16 it is not known if LT binds directly to these neurones. Cholera toxin B subunit was found to bind to specific receptors exclusively on VIP containing neurones40; however, there are no such data for LT. In the present experiments, we demonstrated that VIP plays an important role in this secretory process as VIP antagonism completely prevented LT induced secretion.
STa is structurally different from the two other toxins and induces intestinal secretion through an increase in cGMP.17-19 In addition, it has been demonstrated that, in common with CT and LT, the enteric nervous system is important in STa induced secretion as shown by the inhibitory effect of neuronal blockade.17 ,19-23The neuronal mediators involved have not been fully elucidated. 5-HT was not found to be implicated in STa induced secretion.14 ,22 However, Rolfe and Levin have recently shown that nitric oxide synthase inhibition as well as capsaicin inhibit STa secretion in the rat ileum and concluded that STa activates a nitric oxide dependent myenteric plexus secretory reflex mediated by capsaicin sensitive C fibres.22 ,41 In addition, it seems that tachykinin NK2 receptors are activated to induce secretion during STa exposure.29 In the present experiments, VIP antagonism partially prevented STa induced secretion demonstrating that VIPergic neurones are activated during STa induced secretion. VIPergic neurones and nitric oxide synthase containing neurones coexist in the enteric nervous system.42 In addition, nitric oxide may release VIP from nerve terminals,43 and VIP may cause release of nitric oxide.44 Thus we may speculate that nitric oxide and VIP interact to induce secretion in STa. This speculation needs to be confirmed in further studies.
In conclusion, our findings support the view that VIP participates in cholera toxin and E coli enterotoxin induced secretion, presumably through activation of an efferent limb of the secretory neuronal reflex. These findings may have clinical applications in the future when VIP antagonists for human use become available.
Abbreviations used in this paper
- CT
- cholera toxin
- LT
- Escherichia coli heat labile toxin
- STa
- Escherichia heat stable toxin
- VIP
- vasoactive intestinal polypeptide
- VIPa
- vasoactive intestinal polypeptide antagonist
- 5-HT
- 5-hydroxytryptamine
- PES
- plasma electrolyte solution
- PEG
- polyethylene glycol