Article Text

Abstract
BACKGROUND Primary sclerosing cholangitis (PSC) is considered to be a chronic autoimmune disease where infiltrating T lymphocytes have been implicated in the destruction of bile ducts. Altered function of these T cells may reflect abnormalities in the immune response leading to tissue damage.
AIM We investigated the proliferative and functional capacity of freshly isolated liver derived T lymphocytes (LDLs) and natural killer (NK) cells from PSC patients.
METHODS The proliferative responses to common mitogens such as phytohaemagglutinin (PHA), concanavalin A (Con A), and lipopolysaccharide (LPS) were studied, and the cytotoxic function of T lymphocytes was measured using allogeneic target cells. NK (CD56+/16+) cytotoxic function was measured using the two cell lines K562 (NK sensitive) and Raji lymphoma cells (NK resistant).
RESULTS Compared with patients with primary biliary cirrhosis (PBC), autoimmune hepatitis (AIH), and normal controls (without liver disease), in PSC: (1) LDLs contained a low percentage of T cells; (2) there was significantly decreased expression of interleukin (IL)-2 receptor (p<0.001) on activated T cells (HLA-DR+); (3) LDLs but not peripheral blood lymphocytes had significantly impaired proliferative responses to mitogens such as PHA, Con A, and LPS (p< 0.001); (4) no cytotoxic activity of PSC liver T and NK cells was recorded; (5) significantly higher levels of tumour necrosis factor α (TNF-α) and IL-1β but lower levels of IL-2, IL-10, and interferon γ were found in the supernatants of mitogen stimulated LDL cultures (p<0.001); (6) higher percentages of freshly isolated PSC LDLs contained intracytoplasmic TNF-α and IL-1β; and (7) pretreatment of PSC LDLs in vitro with neutralising TNF antibodies significantly enhanced proliferative responses and allowed IL-2 receptor expression following stimulation. In addition, the impaired cytolytic activity of both NK and T cells was partially restored. Impaired proliferative or functional capacity of liver derived T cells was not observed in either PBC or AIH patients.
CONCLUSIONS We suggest that reduced T cell reactivity in liver infiltrating cells obtained from patients with PSC is due to high local production of TNF-α. Our findings indicate that the use of anti-TNF antibodies as an alternative treatment for PSC patients should be evaluated.
- autoimmune liver diseases
- biliary epithelial cells
- cytokines
Abbreviations used in this paper
- BECs
- biliary epithelial cells
- IBD
- inflammatory bowel disease
- FITC
- fluorescein isothiocynate
- IL-1β
- interleukin 1β
- IFN-γ
- interferon γ
- LDLs
- liver derived lymphocytes
- PHA
- phytohaemagglutinin
- Con A
- concanavalin A
- LPS
- lipopolysaccharide
- PBC
- primary biliary cirrhosis
- PSC
- primary sclerosing cholangitis
- TNF-α
- tumour necrosis factor α
- AIH
- autoimmune hepatitis
- PBS
- phosphate buffered saline
- NK
- natural killer
- PBLs
- peripheral blood lymphocytes
- cpm
- counts per minute
Statistics from Altmetric.com
Primary sclerosing cholangitis (PSC) is characterised by inflammatory fibrosis of the intrahepatic and extrahepatic biliary tracts.1 ,2 Patients with PSC frequently develop progressive hepatobiliary fibrosis with eventual development of cirrhosis, portal hypertension, liver failure, and premature death. The aetiology of PSC is unknown although lymphocytic infiltration in areas of portal damage suggests an autoimmune mediated destructive process.3 Infiltration by T lymphocytes is a feature shared by several organ specific autoimmune diseases. Periportal mononuclear cell infiltration with at least 80% T lymphocytes is found in most liver biopsy specimens of PSC patients.4 Some of these T cells show evidence of activation, as indicated by expression of HLA-DR.5 In 70% of cases PSC is associated with inflammatory bowel disease (IBD), most commonly ulcerative colitis. This close association further suggests that PSC may be an autoimmune disease.6
The pathophysiological mechanisms underlying the development of PSC are unknown. In PSC the final target of the inflammatory process appears to be the biliary epithelium.7 The cause of epithelial cell destruction is not known. There is increasing evidence that locally produced cytokines may play a critical role, and may be directly responsible for epithelial cell injury by altering epithelial cell function or integrity.8 The effects of cytokines in other chronic inflammatory disorders such as rheumatoid arthritis,9 Crohn's disease,10psoriasis,11 and glomerulonephritis12 have already been reported. In IBD patients, proinflammatory cytokines such as tumour necrosis factor α (TNF-α), interleukin (IL)-1β, and interferon γ (IFN-γ) have been reported to be increased and it is suggested that IBD is attributable to an imbalance in Th1 and Th2 CD4 T cell activity with a predominant Th1 cell response.13-15
Neither the detection or presence of cytokines in the livers of PSC patients nor the role played by cytokines in the destruction of biliary epithelium has been elucidated previously. In addition, so far the effects of proinflammatory cytokines on freshly isolated cells from the sites of inflammation in patients with autoimmune liver diseases have not been studied. In most reports to date, the functional capacity of liver derived lymphocytes (LDLs) have been studied using cells propagated in vitro, indicating the possibility of strong selection bias, and therefore such cells may represent only a fraction of the LDLs present in situ.
Autoimmune injury has been suggested to be responsible for bile duct destruction in PSC16 and enhanced autoreactivity of peripheral T lymphocytes in these patients has been reported.17 The aim of our study was to functionally characterise lymphocytes freshly isolated from the livers of PSC patients to evaluate alterations in the function of LDLs that may reflect abnormalities that could be associated with tissue damage. In all experiments, cell samples were compared with those from normal controls and from patients with other autoimmune liver diseases.
Materials and methods
PATIENTS AND CONTROLS
Seven PSC patients (four females), median age 42 (36–63) years, all with end stage cirrhotic disease, were studied. Five patients also had ulcerative colitis but none had cholangiocarcinoma. The diagnosis of PSC was based on accepted clinical, histological, and cholangiographic criteria.7 Four primary biliary cirrhosis (PBC), three autoimmune hepatitis (AIH) patients, and eight healthy donor livers were also included. Diagnoses of PBC and AIH were based on accepted criteria.18 ,19 All patients with PBC were treated with ursodeoxycholic acid. Patients with AIH were treated with steroids and azathioprine. All patients with PBC and AIH had end stage liver disease with biopsy proved cirrhosis. None had hepatitis B or C infections.
ISOLATION OF MONONUCLEAR CELLS FROM PERIPHERAL BLOOD
Peripheral blood lymphocytes (PBLs) were isolated from patients using density gradient cell separation. PBLs were obtained before any blood products were given. Cells were suspended in RPMI 1640 cell culture medium (Gibco, Paisley, UK) supplemented with 10% heat inactivated fetal calf serum, 2 mM l-glutamine, 25 mM HEPES, and antibiotics (complete medium), and adjusted to a concentration of 1×106 cells/ml.
ISOLATION OF LIVER LYMPHOCYTES
Liver tissues were obtained from organs removed from recipients and organ donors at the time of liver transplantation. Permission was granted from the local ethics committee. Initially, the livers were washed extensively with phosphate buffered saline (PBS) to remove all blood and contaminating peripheral lymphocytes. Tissues were then chopped with sterile scalpels and enzymatically digested with sterile medium containing 0.05% (w/v) collagenase and 0.002% (w/v) DNase (both from Sigma, Steinhem, Germany) in a 37°C water bath for 30 minutes. The digested cell suspensions were passed through a nylon mesh filter (25 μm pore size) to remove undissociated tissue and cell clumps. The isolated cells were washed once with PBS to remove mucous-like material and centrifuged at 350g for 10 minutes. Thereafter lymphocytes were separated on Lymphoprep using density gradient centrifugation.
Except for phenotyping and intracellular cytokine labelling, all experiments were performed after overnight incubation of LDLs on plastic to remove adherent cells. Hence the mononuclear cell populations consisted mainly of T lymphocytes, CD56+/16+cells, and some B cells.
ISOLATION OF CD56+/16+ CELLS FROM LDLs
Cells were added to culture flasks and incubated overnight to remove adherent cells. All non-adherent cells were collected from the flasks, washed once with PBS, and centrifuged at 200g for five minutes. Anti-CD3 antibodies (clone SK7; cat. No 347340; Becton-Dickinson, San Jose, California, USA) 20 μl/106 cells were added to the pellet and incubated for 30 minutes at room temperature. Cells were washed twice with PBS, and anti-CD3 reactive lymphocytes were bound to rat antimouse IgG1 coated magnetic beads (cat. No. 110.12; Dynal Inc., Oslo, Norway) 30 μl beads/106 cells, following incubation on a rock “n” roller for 30 minutes at room temperature. Anti-CD3 reactive lymphocytes were removed by placing the mixture in a magnetic field. The supernatant was centrifuged to obtain cells that were depleted of CD3+ lymphocytes. The use of rat antimouse IgG1 coated magnetic beads was repeated a second time to remove any remaining CD3+ cells. The cells were next incubated with anti-CD19 antibody coated magnetic beads 20 μl/106 cells (cat. No 111.04; Dynal Inc.), and incubated at room temperature on a rock “n” roller for 20 minutes. Anti-CD19 reactive lymphocytes were removed by placing the mixture in a magnetic field. The average phenotype of the negatively selected natural killer (NK) cell enriched fraction was 95% CD3−, CD16+, and CD56+ cells, and less than 1% CD3+ cells, as indicated by flow cytometric analysis.
ISOLATION OF BILIARY EPITHELIAL CELLS
Human intrahepatic biliary epithelial cells (BECs) were isolated from the livers of two normal healthy liver donors using a method previously described.20 In brief, liver tissue was mechanically disrupted and enzymatically digested as described above and the non-hepatocyte fraction was collected by density gradient centrifugation using Percoll. BECs were further purified by immunomagnetic isolation using Dynabeads conjugated to the epithelial cell specific antibody BerEp4 (Dynal). Approximately 2×107 immunomagnetic beads were washed and incubated with the cells extracted from the density gradient interface. The mixture was incubated at 4°C for 30–40 minutes with gentle agitation. Cells coupled to the beads were isolated and washed by applying a magnetic particle concentrator (Dynal). Immunoisolated BECs were resuspended in a 1:1 mixture of Ham's F12 and DMEM media (both from Gibco), supplemented with 5% fetal bovine serum, 5 ng/ml epidermal growth factor, 30 ng/ml cholera toxin, 0.4 μg/ml hydrocortisone hemisuccinate, 2 nM tri-iodo-thyronine and 5 μg/ml insulin (all from Sigma), and 10 ng/ml human recombinant hepatocyte growth factor (R&D Systems, Abingdon, UK), and seeded in gelatin coated tissue culture flasks. Cells were grown at 37°C in a humidified atmosphere of 5% CO2 in air. Cell monolayers were passaged at confluence using trypsin-EDTA.
LABELLING OF PBLs AND LDLs FOR T AND NK CELL MARKERS WITH CONJUGATED ANTIBODIES
Isolated PBLs/LDLs (1×104) were incubated with 5 μl of fluorescein isothiocynate (FITC) or phycoerythrin conjugated monoclonal antibodies in 50 μl of PBS-0.1% sodium azide solution for 30 minutes at 4°C. Cells were washed once with PBS-sodium azide and fixed in 1% (wt/vol) paraformaldehyde solution in PBS.
The monoclonal antibodies (Becton-Dickinson, Mountain View, California, USA) used in the study included the following: anti-CD3, anti-CD4, anti-CD8, anti-CD56, anti-CD16, anti-CD25, and anti-HLA-DR. Mouse isotypes IgG1 and IgG2a were used as controls in all experiments. Two colour analysis was performed on a FACSorter (Becton Dickinson). For intracellular cytokine staining of LDLs, FITC conjugated antihuman TNF-α (IC210F) and anti-human IL-1β (IC201F) (both antibodies from R&D systems) were used. The procedure for intracellular staining was performed according to the manufacturer's instructions.
PHENOTYPING OF BECs
Single colour fluorescence was used to phenotypically characterise the BECs. Primary antibodies for intracellular staining were directed to cytokeratin 7 and 19 (Dako, A/S, Denmark). For labelling of intracellular antigens,21 cells were pelleted and washed once in 300 μl of PBS containing 0.5% (w/v) saponin (Sigma). Primary antibodies at predetermined concentrations were added to the cells and incubated for 30 minutes at 4°C. Cells were washed twice with cold PBS with saponin. An appropriate FITC conjugated goat antimouse secondary antibody (Immunotech, Marseille, France) was added and incubated for 20 minutes at 4°C. Cells were washed once prior to analysis on the flow cytometer. Antibodies against surface receptors included antihuman HEA-125 (epithelial specific antigen) (Serotec, Oxford, UK) used for phenotyping of epithelial cells, FITC labelled antihuman TNF receptor I (TNF RI, clone 16803.1), TNF receptor II (TNF RII, clone 22235.311; both from R&D Systems), and anti-CD95 (Fas) (Becton-Dickinson).
PROLIFERATION ASSAYS
Triplicate cultures of 100 μl of LDLs or PBLs (1×105) suspended in complete medium were distributed into a 96 well tissue culture plate. A sample of 100 μl of either phytohaemagglutinin (PHA 5 μg/ml), concanavalin A (Con A 5 μg/ml), lipopolysaccharide (LPS 10 ng/ml), or medium alone were added to the wells and incubated for 48 hours at 37°C. 3H thymidine was then added, harvested after another 24 hours of incubation with a semiautomated cell harvester (Titertek 550; Flow laboratories, Irvine, Ayrshire, UK), and counted in a liquid scintillation counter (LKB Instruments Inc., Bromma, Sweden). Results are expressed as arithmetic mean counts per minute (cpm) of triplicate cultures (SEM). The experiments were also performed with only CD3+ T cell fractions.
CELL MEDIATED LYMPHOLYSIS ASSAY
Two non-adherent cell lines were used as targets for the cytotoxicity experiments: K562 (NK cell sensitive) and Raji (NK resistant). These cells were obtained from the American Type Culture Collection (ATCC, Rockville, Maryland, USA) and maintained in RPMI 1640 complete medium. Immediately before each cytotoxicity assay, both target cell types were harvested and pelleted by centrifugation (400g, 10 minutes). Target cells (1×106) in 100 μl complete medium were labelled with 100 μCi of 51CrO4Na2 (Amersham, Bromma, Sweden) for one hour at 37°C in a humidified 5% CO2 atmosphere and washed three times before being resuspended in complete medium at a concentration of 1×105 cells/ml. Freshly isolated effector cells (CD56+/16+/CD3− cells from livers of PSC/PBC/AIH/normals) were resuspended at an appropriate concentration and aliquoted (100 μl) in triplicate in 96 U bottom microculture plates (Costar, Cambridge, Massachusetts, USA). The labelled target cells were incubated with the CD56+/16+ cells at effector to target ratios of 50:1, 25:1, 12:1, and 6:1 in a total incubation volume of 200 μl. The plates were centrifuged at 200 g for five minutes, incubated at 37°C for four hours, and subsequently aliquots of supernatant (100 μl) were counted for radioactivity. Spontaneous51Cr release was determined by counting the supernatant radioactivity present in the cultures of target cells in medium alone. Maximal release was determined by incubating the target cells with 5% Triton X-100. Cytotoxicity was expressed as percentage specific51Cr release using the formula:
% specific release = ((experimental cpm−spontaneous cpm)/(maximum cpm–spontaneous cpm))×100.
For detection of the functional capacity of liver derived cytotoxic T cells, LDLs from each PSC/PBC/AIH patient were mixed with allogeneic irradiated (2000 rads) LDLs (from normals) at a cell concentration of 1.5×106/ml and a responder:stimulator ratio of 1:1 (MLC), and incubated in flasks for five days at 37°C. Three days prior to the cell mediated lympholysis assay, PHA (10 μg/ml) was added to fresh allogeneic LDL target cells and incubated at 37°C. On day 6, PHA stimulated LDLs were washed once in PBS and labelled with 100 μCi of 51Cr for one hour at 37°C, washed three times, and resuspended at 1×105 cells/ml in complete medium. Effector cells from the five day MLC cultures were washed, counted, and distributed in U bottomed microtitre plates at effector to target ratios of 50:1, 25:1, 12:1, and 6:1. After incubation for four hours at 37°C, the amounts of radioactivity released into the supernatants were measured and the percentage specific cytotoxicity was calculated as above. Control experiments were performed using LDLs from the four normal livers stimulated against each other.
CYTOKINE ASSAYS
PSC, PBC, AIH, and normal LDL culture supernatants from unstimulated, mitogen stimulated, and cultures neutralised with anti-TNF-α or anti-IL-1β were collected after 48 hours, sterile filtered, and kept frozen at −70°C until assayed. The cytokines TNF-α, IFN-γ, IL-1β, IL-2, IL-10, and IL-12 were measured by standard sandwich ELISA techniques using the Quantikine sandwich enzyme immunoassay from R&D systems. Assays were performed according to the manufacturer's instructions. In brief, samples and standards were incubated in anticytokine antibody coated wells. After washing, a polyclonal antibody conjugated to horseradish peroxidase was added. After another incubation and washing, a chromogen-peroxide substrate solution was added. Colour development was stopped with sulphuric acid and the intensity of the colour was measured.
NEUTRALISING ANTIBODY STUDIES
Anti-hTNF-α (MAB210) and anti-hIL-1β (MAB201) neutralising antibodies were purchased from R&D systems. The anti-hTNF-α monoclonal antibodies can neutralise the biological activity of both soluble and membrane bound TNF-α. LDLs were incubated with neutralising antibodies (10 μg/ml) for two hours and the proliferation assay carried out as described previously. An additional control included cells with the neutralising antibodies alone. For the cytolytic assays, LDLs were incubated with the neutralising antibodies overnight.
STATISTICAL ANALYSIS
When comparing differences in proliferative response and cytotoxic functions between different groups, the Kruskall Wallis test was used. The Mann-Whitney U test was used when differences between PBL and LDLs within the same patient were compared. Calculations were made with Statistica/Wc (Statsoft, Inc, Tulsa, Oklahoma, USA).
Results
SIGNIFICANTLY DECREASED EXPRESSION OF IL-2 RECEPTOR ON ACTIVATED T CELLS DERIVED FROM PSC LIVERS
We phenotypically characterised T and NK cells from PBLs and LDLs, including expression of activation markers, for example IL-2 receptor (CD25) and HLA-DR (table 1). Among the cells obtained from control liver tissues, T lymphocytes and NK cells (CD56+/CD16+/CD3−) represented the two major populations. Lower proportions of CD3+ and NK cells were found in PSC livers compared with PBC, AIH, and normal livers. A significantly higher fraction of liver derived CD3+ cells from all three disease groups expressed HLA DR compared with normals. However, the most interesting difference was very low expression of CD25 on activated T cells from PSC patients compared with PBC, AIH, and normals (p<0.001). In contrast, as shown in table 1, comparing PBLs and LDLs in PSC patients, we found that the proportions of CD3+ and CD25+ T cells in PBLs were normal.
Phenotypic analysis of peripheral blood and liver derived lymphocytes from normal healthy liver donors, and from primary sclerosing cholangitis (PSC), primary biliary cirrhosis (PBC), and autoimmune hepatitis (AIH) patients
TNF-α AND IL-1β EXPOSED BILIARY EPITHELIAL CELLS EXPRESS HIGHER LEVELS OF TNF-RI AND RII
BECs were routinely characterised during growth by immunofluorescence labelling and flow cytometry. Results indicated that the cultures were more than 95% pure for BECs on the basis of expression of cytokeratins 19 and 7. Other contaminating cells (endothelial cells and fibroblasts) and smooth muscle cells were less than 2%.
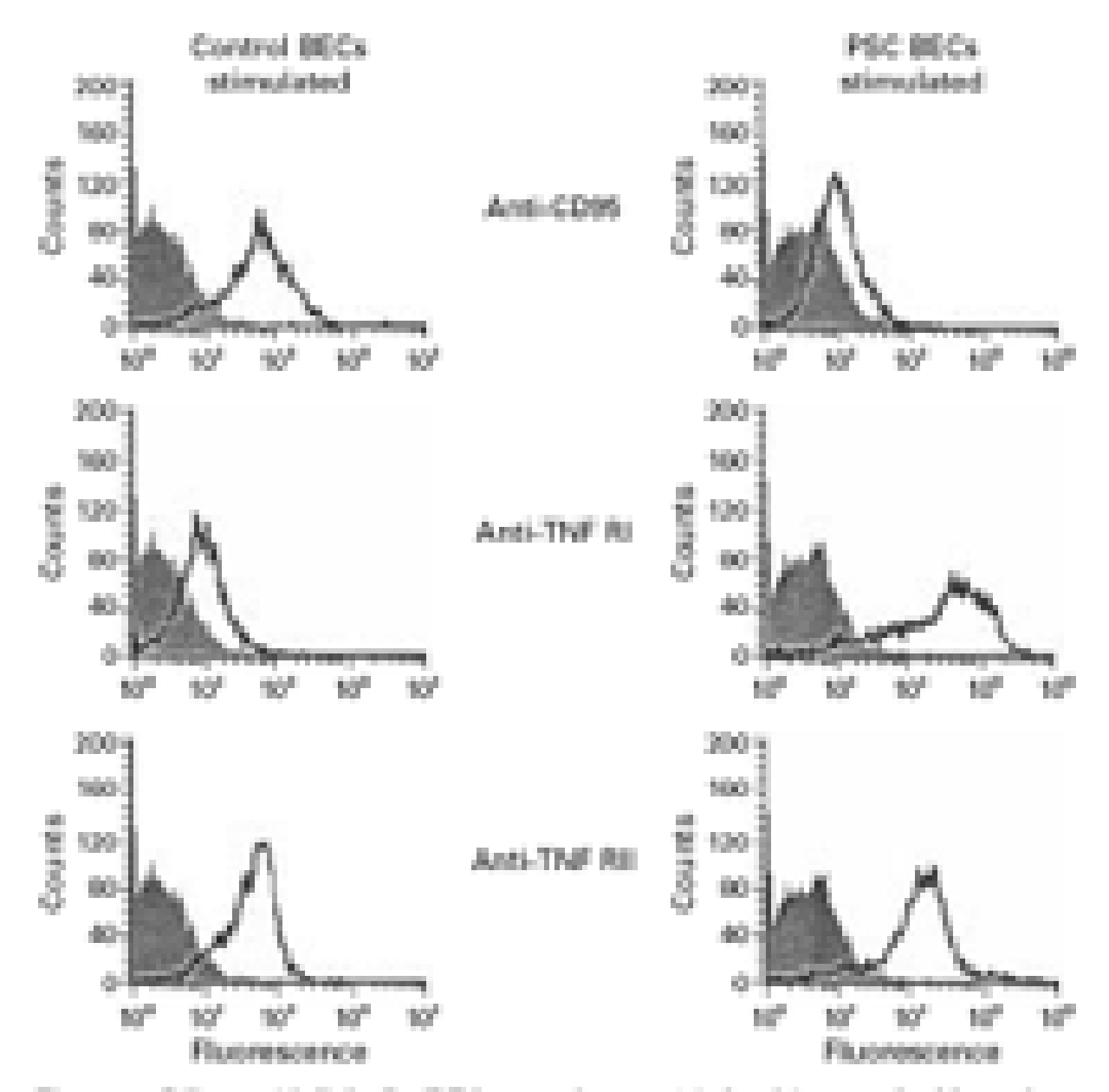
BECs isolated from the livers of three PSC patients and two normal controls (second passage) were tested for expression of CD95 and the two TNF receptors RI and RII. A marked morphological difference was observed in normal BECs compared with PSC BECs. Normal BECs showed typical epitheloid morphology while PSC BECs were more elongated and vacuolated (manuscript in preparation). Freshly isolated BECs from normals or PSC livers did not express either of the TNF receptors. Both types of BECs expressed cytokeratin 7. However, lower expression of cytokeratin 19 (data not shown) and CD95 was observed on PSC BECs compared with control BECs (fig 1). Cytokine stimulation with TNF-α and IL-1β showed that PSC BECs expressed higher levels of the two TNF receptors (RI and RII) with no change in CD95 compared with controls (fig 1).

Biliary epithelial cells (BECs, second passage) isolated from two healthy and three primary sclerosing cholangitis (PSC) livers were tested for expression of CD95 and the two tumour necrosis factor (TNF) receptors to study the role of BECs as targets for lymphocyte mediated or direct cytokine mediated damage. TNF-α and IL-1 stimulated PSC BECs demonstrated increased expression of the TNF RI and RII receptors but low expression of CD95. Normal BECs expressed CD95 and TNF RII but had lower expression of TNF RI.
DIMINISHED PROLIFERATIVE RESPONSES OF MITOGEN STIMULATED T CELLS ISOLATED FROM PSC PATIENTS
As seen in fig 2, a low proliferative response induced by PHA, Con A, and LPS in LDLs isolated from PSC patients was seen. This was significantly different from LDLs obtained from PBC, AIH, and normal livers (p<0.001) (fig 2). Similar results were obtained when pure populations of CD3+ T cells were used (data not shown).

In general, liver derived lymphocytes (LDLs) isolated from controls (n=8) on stimulation with mitogens such as concanavalin A (Con A) and phytohaemagglutinin (PHA) for 72 hours showed lower proliferation responses than those observed in peripheral blood lymphocytes from the same on stimulation. No difference in response was observed after pretreatment with anti-tumour necrosis factor (TNF) monoclonal antibodies. Significantly diminished proliferation of LDLs from the seven primary sclerosing cholangitis (PSC) patients was observed on stimulation which was restored to almost normal by pretreatment with anti-TNF monoclonal antibodies (p<0.001). No significant differences in proliferative responses in mitogen stimulated LDLs from primary biliary cirrhosis (PBC) or autoimmune hepatitis (AIH) patients (either before or after anti-TNF monoclonal antibody pretreatment) was observed compared with controls (NS). Data represent mean counts per minute (cpm) of triplicate cultures. LPS, lipopolysaccharide.
Comparison between mitogen stimulated proliferative responses of PBLs and LDLs from PSC patients showed a significant difference. They were significantly higher in PBLs than in LDLs (fig 3) (p<0.001) but lower compared with PBLs from normal controls. In cells from PBC and AIH patients, no significant differences were observed in proliferative responses of either PBLs or LDLs compared with normals (NS).

Peripheral blood lymphocytes (PBLs) isolated from controls (n=8), primary sclerosing cholangitis (PSC) (n=7), primary biliary cirrhosis (PBC) (n=4), and autoimmune hepatitis (AIH) (n=3) patients showed normal proliferative responses on stimulation with mitogens such as concanavalin A (Con A) and phytohaemagglutinin (PHA) for 72 hours. No difference in responses was observed after pretreatment with anti-tumour necrosis factor (TNF) monoclonal antibodies. Data represent mean counts per minute (cpm) of triplicate cultures. LPS, lipopolysaccharide.
As no proliferative responses were observed in LDLs from PSC patients, we performed the same assays using shorter periods of time (4, 8, 12, 16, 24, 36, and 48 hours). Diminished proliferative responses were observed at all time points (data not shown).
IMPAIRED CYTOLYTIC ACTIVITY OF T LYMPHOCYTES AND NK CELLS ISOLATED FROM THE LIVERS OF PSC PATIENTS
We found that the cytotoxic functions of T and NK cells obtained from the livers of PSC patients were significantly impaired compared with PBC, AIH, and normal controls (p<0.001) (fig 4). PSC liver CD56+/16+ cells did not kill K562 or Raji. However, NK cells from PBC, AIH, and normal livers spontaneously killed K562 but not the Raji cell line, and cytotoxic T cells clearly showed high cytotoxic function against allogeneic target cells (fig4).

Cytotoxic liver derived lymphocytes (LDLs) (T cells) generated by stimulation against normal allogeneic LDLs from control, primary biliary cirrhosis (PBC), and autoimmune hepatitis (AIH) livers showed high cytotoxic function against allogeneic target cells. In the case of controls, only the average per cent lysis (from eight experiments in normals, four in PBC, and three in AIH) is shown. Cytotoxic LDLs (T cells ) isolated from primary sclerosing cholangitis (PSC) livers (n=7) on the other hand did not show any cytotoxic activity against allogeneic target cells. The cytotoxic activity of T cells was thus completely abolished in the livers of PSC patients (p<0.001). Natural killer (NK) cells (CD56+16+) isolated from control, PBC, and AIH livers showed normal cytotoxic activity against K562 (NK cell sensitive cell line) but not against Raji's (NK resistant cell line) (only mean per cent lysis is shown). NK cells isolated from PSC livers (n=7) on the other hand did not show any cytotoxic activity against K562. NK cell cytotoxic activity was thus completely abolished in the livers of PSC patients (p<0.001). However, overnight treatment of cytotoxic LDLs (T cells) isolated from PSC livers with tumour necrosis factor (TNF) antibodies (abs) (10 mg/ml) showed a moderate increase in cytotoxic activity against allogeneic target cells. Similarly, NK cells (CD56+/16+) showed enhanced cytotoxic activity against K562 (NK cell sensitive cell line).
As LDLs derived from the livers of PSC patients had impaired proliferative and functional capacity, we decided to screen supernatants of mitogen stimulated cultures for the presence of cytokines, as certain proinflammatory cytokines may inhibit lymphoproliferative responses in vitro.
HIGH LEVELS OF TNF-α AND IL-1β IN THE SUPERNATANTS OF PHA AND CON A STIMULATED LDLs FROM PSC PATIENTS
Supernatants were collected after 48 hours from cell cultures stimulated with either PHA, Con A, or LPS and analysed for the presence of TNF-α, IFN-γ, IL-1β, IL-2, IL-10, and IL-12. As shown in table2, significantly higher levels of TNF-α and IL-1β, and lower concentrations of IL-2, IL-10, and IFN-γ were detected in the supernatants of LDL cell cultures from PSC patients stimulated with PHA and Con A compared with PBC, AIH, and normal controls (p<0.001). Supernatants collected after 4, 8, 12, 16, 24, and 36 hours gave the same results (data not shown).
Cytokine production in supernatants of mitogen stimulated or anti-tumour necrosis factor (TNF) monoclonal antibody treated lymphocytes from primary sclerosing cholangitis (PSC) and control livers
Flow cytometric analysis of three day mitogen stimulated LDLs showed that T lymphocytes and CD56+/16+ cells were the major populations present in cultures.
INCREASED INTRACELLULAR EXPRESSION OF TNF-α AND IL-1β IN LDLs FROM PSC LIVERS
LDLs from all PSC, PBC, AIH, and control livers were stained for intracellular expression of TNF-α and IL-1β. We found varying but increased percentages of PSC LDLs possessing TNF-α and IL-1β. LDLs from one PBC patient had a high proportion of IL-1β but not of TNF-α secreting cells. Only one control liver showed binding slightly higher than background (table 3). A representative example of the increased intracellular binding of anti-TNF-α and anti-IL-1β in LDLs from one PSC patient compared with one control is shown in fig5.
Percentage of liver derived lymphocytes (LDLs) positive for intracytoplasmic staining of anti-tumour necrosis factor α (TNF-α) and interleukin 1 β (IL-1β) cytokines from primary sclerosing cholangitis (PSC) and control patients

Increased immunofluorescent staining of intracellular cytokines tumour necrosis factor α (TNF-α) and interleukin 1β (IL-1β) was seen in freshly isolated liver derived lymphocytes from all seven primary sclerosing cholangitis (PSC) patients but not from controls. A representative picture of this finding from one PSC patient and one normal control is shown. Cells were initially fixed, permeabilised, and stained with fluorescein isothiocynate conjugated anti-TNF/anti-IL-1β antibodies. Unlabelled anti-cytokine monoclonal antibody was used to decrease high background staining.
TREATMENT WITH ANTI-TNF MONOCLONAL ANTIBODY RESTORES THE PROLIFERATIVE RESPONSES OF PSC LDLs
The final concentration of the anti-TNF monoclonal antibody was determined in initial titration experiments of time and concentrations to detect the optimal time and amount required for neutralising the TNF-α effect (data not shown). Treatment with 10 μg/ml overnight was found to be optimal for our purposes. An isotype matched irrelevant monoclonal antibody (Sigma) was used as a control. No non-specific stimulation using the control antibodies was observed (see fig 6). The anti-TNF-α antibody at a final concentration of 10 μg/ml neutralised the effect of 100 ng/ml TNF-α. Anti-TNF treated cell culture supernatants showed normal levels of IL-2, IL-10, and IFN-γ but lower TNF-α and IL-1β levels (table 2).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
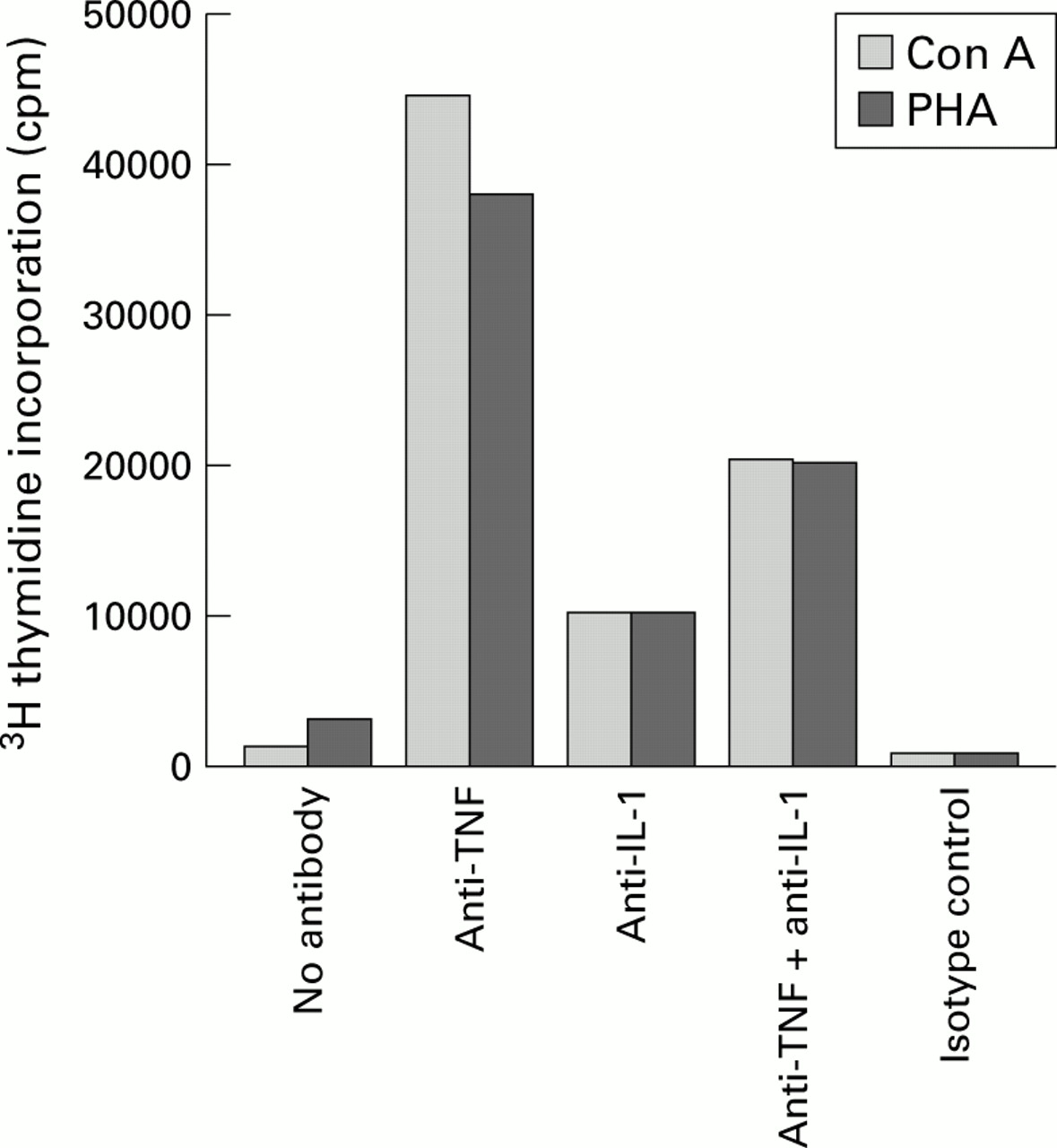
Effect of anti-cytokine antibodies on restoration of diminished proliferative responses to mitogen stimulation by concanavalin A (Con A) and phytohaemagglutinin (PHA). Liver derived lymphocytes (LDLs) from primary sclerosing cholangitis (PSC) livers remained untreated or pretreated with anti-tumour necrosis factor (TNF) monoclonal antibodies (10 μg/ml) and/or interleukin 1 (IL-1) monoclonal antibodies (10 μg/ml), or control monoclonal antibodies (10 μg/ml) overnight. Con A or PHA was added and 3H thymidine incorporation was detected after 72 hours. Anti-TNF monoclonal antibodies alone restored the diminished proliferative responses in the LDLs from PSC patients.
Anti-TNF treated LDLs from PSC patients demonstrated significantly enhanced proliferation (p<0.001) comparable with controls (fig 2). However, no difference in the proliferative responses of PBC, AIH, and normal LDLs before or after anti-TNF treatment was observed (figs 2 and3).
As high levels of IL-1β were also detected in supernatants from mitogen stimulated LDL cultures, we decided to study the effect of anti-IL-1β monoclonal antibody on the proliferative responses of these cells. As in the case of anti-TNF antibody, titration studies indicated that a concentration of 10 μg/ml with overnight treatment was optimal for our purposes (the PBC patient with the high levels of IL-1β was used as a positive control). However, no effect of anti-IL-1β antibody was observed on levels of other cytokines produced and detected in the supernatants or on the proliferative responses of LDLs from PSC, PBC, or AIH.
Although a combination of anti-TNF and anti-IL-1β enhanced the proliferative responses of LDLs to some extent, the best effect was obtained using anti-TNF alone. Thus anti-IL-1β alone or in combination with anti-TNF was not as effective in restoring impaired proliferative responses as anti-TNF alone (fig 6).
TREATMENT WITH ANTI-TNF MONOCLONAL ANTIBODY RESTORES PARTIALLY THE CYTOLYTIC CAPACITY OF PSC T AND NK CELLS
Anti-TNF treated cytotoxic T and CD56+16+cells from PSC patients demonstrated a moderate increase in cytotoxic activity to allogeneic target cells and K562, respectively, compared with activity prior to anti-TNF antibody treatment (fig 4).
Discussion
In the present study we functionally characterised freshly isolated cells from blood and the site of inflammation (that is, the liver) in PSC patients. Our results show that cell proliferation and the function of liver derived T lymphocytes is impaired in PSC patients compared with normal controls and other patients with autoimmune liver diseases. We believe this is a consequence of exposure (maybe even chronic) to high levels of TNF in vivo, as (a) freshly isolated LDLs from PSC patients showed increased expression of intracellular TNF-α, (b) in vitro, supernatants from mitogen stimulated, freshly isolated LDLs showed the presence of significantly high levels of TNF-α compared with controls, and (c) addition of anti-TNF antibodies restored the diminished proliferative responses in vitro. Alternatively, one may speculate that T cells had undergone TNF-α induced apoptosis and that addition of anti-TNF antibodies inhibited apoptosis. However, our investigations showed that even though a small number of cells underwent apoptosis (approximately 15% using annexin V), the rest of the cells survived (data not shown) at the end of the assay but did not proliferate.
Our results are supported by the findings of Spengler and colleagues22 who reported the presence of relatively high levels of TNF-α in culture supernatants of PHA stimulated T cell lines from liver biopsies in patients with different stages of PSC while decreased levels were observed in PBC patients. These results indicate that increased levels of TNF-α are already present in PSC patients even in the early stages of the disease and are not restricted to only the late stages. In addition, Berg and colleagues,23 based on the cytokine profile obtained using supernatants from blood lymphocytes, have classified PSC as TH1 (T helper 1 subclass of T cells) mediated as the TH2/TH1 ratio was found to be 1:4 while in PBC the ratio was 1:1. TH1 cells produce proinflammatory cytokines, including IFN-γ, IL-2, and potentially TNF. Therefore, based on reports from other studies, including the current one, it may be hypothesised that LDLs in PSC are chronically exposed to TNF-α in vivo and therefore this accounts for the impaired in vitro proliferative and functional capacities. Thus even though the current observations were made in patients with end stage liver disease, they may reflect immunological changes already occurring during the initial stages of the disease. Our findings are specific to PSC patients as impaired proliferation was not observed in either PBC or AIH patients with end stage liver disease and is therefore not only explained by the end stage nature of the disease.
In addition, we found significantly higher levels of TNF-α and IL-1β but significantly lower levels of IL-2, IL-10, and IFN-γ in the supernatants of mitogen stimulated LDLs obtained from PSC livers compared with controls. This effect was reversible as exposure of cells (overnight) to the neutralising effect of anti-TNF-α antibodies restored the proliferative responses. Addition of neutralising anti-IL-1β antibodies alone did not reverse impaired proliferative responses to mitogen stimulation. It should be noted that the anti-TNF-α monoclonal antibodies used neutralise both soluble and membrane bound TNF-α while the anti-IL-1β antibody neutralises only soluble IL-1β and thus is probably less effective in restoring impaired responses.
The impaired cytolytic activity of both NK and T cells was partially restored after treatment with TNF antibodies. It may be possible that repeated treatment with higher doses or longer incubation time with TNF antibodies may be required to see complete reversal of the cytotoxic capacity of these cells. Due to lack of sufficient cells, such studies could not be performed.
Currently, we do not know what stimulates LDLs in PSC to produce high levels of TNF-α. Elevated levels of TNF-α in PSC may have a genetic background. Genetic variation in the TNF-α gene may influence inflammatory and immune mediated diseases. The −308 polymorphism in the TNF-α promoter region (the TNF2 allele) is associated with a six to sevenfold increase in baseline and inducible levels of TNF-α production.24 Recently, Bernal and colleagues25 studied the association of TNF polymorphism with PSC and concluded that genetic susceptibility to PSC may be determined by polymorphism within the TNF genes particularly with the TNF2 allele. Our results support this finding. However, it is important to mention that as studies were performed with material obtained from patients with end stage cirrhotic disease, our findings may not reflect the earlier state of the disease.
The elegant in vitro and in vivo (animal) studies of T lymphocytes chronically exposed to TNF performed by Cope and colleagues26 ,27 showed that chronic exposure to TNF suppresses a broad range of T cell responses and may target T cell receptor signalling28 through partial rather than complete uncoupling of early signalling events within T cells. The proposed immunosuppressive effects of prolonged exposure to TNF are, among others, antiproliferative effects, which influence the precursor frequency of peripheral autoreactive lymphocytes, defective effector T and/or B cell responses, and attenuation of antigen-receptor signalling.29 Our findings of decreased proliferative responses, low levels of IL-2 receptor expression, as well as decreased levels of IL-2, IL-10, and IFN-γ in the supernatants of mitogen stimulated LDLs obtained from the site of inflammation in PSC patients support the findings of Cope et al.
The results of this study also emphasise the importance of using the right source of cells for phenotypic and functional studies as PBLs and LDLs from PSC patients showed significantly varying results. The most striking finding of the phenotypic analysis of LDLs was the very low expression of the IL-2 receptor on activated T lymphocytes from PSC livers compared with PBC, AIH, and normal controls, even though LDLs from all three disease groups had significantly high levels of activated T cells (CD3+/HLA-DR+).
Damage of intrahepatic bile ducts by activated T cells is assumed to be a key event in the pathogenesis of PSC.17 Based on our findings we put forward two hypotheses. Firstly, assessment of the normal functional capacity of freshly isolated cytotoxic PSC LDLs demonstrated decreased or no cytolytic activity. The possibility that T cells mediate biliary epithelial cell injury in PSC by a direct cytotoxic effect seems less likely in view of our inability to detect any normal cytotoxic activity among LDLs from these patients. NK cells may not be involved in the destruction as their normal killing capacity (based on results obtained with K562 cells) was also abolished. These effector cells may play an important role in the early stages of the disease but their role may diminish with time because of chronic exposure to TNF-α and possibly IL-1β. It is conceivable that TNF-α alone or in combination with IL-1β may directly mediate BEC injury. In addition, expression of receptor TNF-R1 on BECs further supports the possibility of induction of apoptosis in BECs by high levels of TNF-α in PSC patients (fig 1). Induction of apoptosis in BECs by TNF-α containing supernatants from PSC LDL cultures is currently under investigation (manuscript in preparation). Even if TNF-α does not directly mediate BEC injury, it may adversely affect BEC in many ways.30 Another important clinical implication is the absence of IL-2 and its role in PSC. IL-2 deficient mice spontaneously develop a chronic, non-granulomatous inflammation of the submucosa and mucosa of the colon, and caecum, which is reported to resemble ulcerative colitis in humans.31 Ciacciet al have shown that intestinal epithelial cells have IL-2 receptors and proliferate in the presence of IL-2.32 Thus the absence of IL-2 in concert with chronic TNF and IL-1β production may further cause BEC injury. The second hypothesis is that as already activated cells respond poorly to mitogen stimulation, allostimulation, and produce less of certain cytokines, it is likely that we are detecting the presence of an antigen specific T cell clone that is expanded in PSC patients. Broomeet al have reported the preferential usage of the Vβ3 region of the T cell receptor in liver T lymphocytes in PSC patients.3 This study needs to be confirmed and further analysis using all of the currently available monoclonal antibodies against the V regions of the T cell receptor needs to be performed.
Our findings may have important clinical implications in that it may be possible to treat PSC patients with anti-TNF-α antibodies. Significant clinical benefits have been observed in patients with rheumatoid arthritis, multiple sclerosis, and Crohn's disease using anti-TNF therapy. However, whether long term chronic TNF blockade aggravates the autoimmune disease process has still to be elucidated. As yet there are no convincing clinical data suggesting that long term TNF blockade might exacerbate autoimmunity. Nevertheless, based on our results we suggest that treatment of PSC patients using anti-TNF antibodies needs to be evaluated in the future.
Acknowledgments
The study was financed by grants from the Magnus Bergvall foundation and the Ruth and Richard Julins foundation, Clas Grotchinsky foundation, the Swedish Medical Society, the Karolinska Institute, and the Nanna Svartz Foundation. We thank Professor Erna Möller and Dr Jan Holgersson for constructive criticism of the manuscript.
Abbreviations used in this paper
- BECs
- biliary epithelial cells
- IBD
- inflammatory bowel disease
- FITC
- fluorescein isothiocynate
- IL-1β
- interleukin 1β
- IFN-γ
- interferon γ
- LDLs
- liver derived lymphocytes
- PHA
- phytohaemagglutinin
- Con A
- concanavalin A
- LPS
- lipopolysaccharide
- PBC
- primary biliary cirrhosis
- PSC
- primary sclerosing cholangitis
- TNF-α
- tumour necrosis factor α
- AIH
- autoimmune hepatitis
- PBS
- phosphate buffered saline
- NK
- natural killer
- PBLs
- peripheral blood lymphocytes
- cpm
- counts per minute