Article Text

Abstract
Background—Genetic polymorphisms inN-acetyltransferase (NAT2) can change the normally fast acetylation of substrates to slow acetylation, and have been associated with the development of some cancers. The NAT2 locus may also suffer dysregulation during cancer progression, as the gene resides on chromosome 8p22, a region which is frequently deleted in colorectal cancer.
Subjects and Methods—A polymerase chain reaction based method was used to determine NAT2 genotype in 275 patients with colon cancer and 343 normal control DNAs. Within the cancer group, 65 cases known to contain deletions in chromosome 8p were examined for loss of heterozygosity at the NAT2 locus.
Results—Overall, there was no statistical difference in frequency or distribution of NAT2 alleles and genotype between colon cancer and control groups. There was a significant association between the slow acetylation genotype and early age of onset. NAT2 genotype did not vary with other clinical features of colon cancer, which included Dukes’s stage, site of tumour, and sex. Of 48 informative cases, only three (6%) showed loss of heterozygosity, indicating that the NAT2 locus is not commonly deleted in colorectal cancer. This suggests that NAT2 is retained during the process of allele loss possibly because of its proximity to a gene necessary for cell viability.
Conclusions—NAT2 does not play a major role in colorectal cancer risk, but may influence risk in some age groups. The nature of the loss of heterozygosity at the chromosome 8p site is complex and is worthy of further study.
- xenobiotic
- N-acetlytransferase 2
- polymorphism
- colorectal cancer
- loss of heterozygosity
- tumour suppressor gene
Statistics from Altmetric.com
- xenobiotic
- N-acetlytransferase 2
- polymorphism
- colorectal cancer
- loss of heterozygosity
- tumour suppressor gene
The polygenic and multifactorial nature of colorectal cancer is clear. It is also apparent that an individual’s reaction to chemical carcinogens or mutagens in the diet is relevant to subsequent development of pathology.1 ,2 For some cancers the risk to the individual may depend on the activation or deactivation of chemical carcinogens by xenobiotic enzymes. The intestine is a primary site of contact with potentially carcinogenic compounds, and colonic crypt cells express several important xenobiotic metabolising enzymes implicated in the development of cancer—for example, CYP1A1 and GSTM1.3 ,4 Functional polymorphisms in the genes coding for enzymes glutathione S-transferase M1 (GSTM1) and T1 (GSTT1) have been associated with an increased risk of colon carcinogenesis,5-7 but recent studies suggest that those initial findings considerably overestimated the risk involved.8
Activity of the cytosolic enzyme N-acetyltransferase 2 (NAT2) has been associated with an increased cancer risk in the bladder and colon.9-11 The NAT2 enzyme acetylates arylamine and hydrazine substrates and is expressed predominantly in the liver. It is also present in epithelial cells of the intestine,3 and may influence the local presentation of carcinogens to intestinal crypt cells, such as the arylamines present in cooked meats.12Polymorphism in the expression and activity of xenobiotic enzymes may therefore be a major factor in the variation seen in the incidence of colorectal cancer in different regions of the world.13 ,14
In the Caucasian population genetic polymorphisms in NAT2 can confer a slow acetylation phenotype by either reducing the stability of the enzyme or reducing transcription of the gene.15-18Analysis of polymorphisms at the NAT2 gene locus can identify individuals with rapid, intermediate, or slow acetylator phenotypes. Each slow allele contains a combination of one or two nucleotide substitutions which occur at positions bp481 (C to T), bp590 (G to A, Arg to Gln), bp803 (A to G, Lys to Arg), and bp857 (G to A, Gly to Glu) of the gene sequence.19 ,20 The substitutions at positions 481 and 803 predominantly co-segregate, whereas substitutions at positions 590 and 857 segregate independently of positions 481, 857, and each other. Additional functional polymorphic sites have been identified in Japanese and African populations, but are extremely rare in European populations.21 ,22
Biochemical studies of the NAT2 phenotype have suggested an increased colorectal cancer risk associated with the fast acetylation phenotype.5 ,10 ,11 However, the increased risk was relatively small and it is possible that the phenotype evaluation was influenced by the specificity of substrates used and by overlapping activity from NAT1 which is also known to be polymorphic.17 In other studies using restriction fragment length polymorphism analysis, the NAT2 genotypes determining fast, intermediate, or slow acetylation were not associated with cancer risk in either colonic adenomas or carcinomas.3 ,23 ,24 These contradictory results may be indicative of the small number of colorectal cancer cases examined but the role of NAT2 acetylation in colorectal cancer risk remains unclear.
The NAT2 locus may also be subjected to dysregulation during tumour progression as the gene resides on chromosome 8p22, a region frequently deleted in colorectal cancer.25-28 Loss of the NAT2 locus has the potential to alter local enzyme activity and could affect subsequent carcinogen presentation. Several separate regions within chromosome 8p have been identified as the minimum regions of overlap of loss of heterozygosity (LOH) in sporadic colorectal carcinomas: the smallest is close to the lipoprotein lipase (LPL) gene, at 8p22; another is in the vicinity of the neurofilament light (NEFL) polypeptide gene at 8p21,25 ,29 and there are apparently distinct consistently deleted regions at 8pter-8p22 and 8p21.3-8p11.22.27 ,29 Analysis of NAT2 LOH may also further localise the region of allele loss on chromosome 8, and putative tumour suppressor gene(s).
Using a polymerase chain reaction (PCR) based NAT2 genotyping assay we have examined NAT2 genotype in constitutive DNA from a group of consecutive patients with colorectal cancer and investigated loss of NAT2 heterozygosity in cancers known to have chromosome 8p loss.
Methods
COLORECTAL CANCER CASES
Peripheral blood and colorectal cancer tissue was collected from a consecutive series of operable patients with colorectal cancer after surgery in four local hospitals between 1988 and 1993. Diagnosis was confirmed histopathologically and patients were classified according to Dukes’s stages (A, B, C), and according to position of cancer in the colon as either right (caecum, transverse, or ascending) or left (sigmoid, descending, or rectum) side. Control samples of peripheral blood were obtained from healthy individuals attending routine occupational screening clinics.30 They were Caucasian, aged 20–75 years, and equally distributed for males and females. Both cancer and control groups were from the same population base. DNA was extracted from peripheral blood lymphocytes and colorectal tissue according to standard methods.31 In 65 patients, in whom there was known to be LOH at 8p, DNA was also extracted from colon cancer tissue. For each colorectal cancer sample a serial fragment of tissue was taken for histological examination.
PCR ASSAY FOR DETECTION OF NAT2 POLYMORPHISMS
A PCR assay was developed using primers designed from the published human gene sequence.32 The primer pair P1 (5′-GCTGGGTCTGGAAGCTCCTC-3′, sense: nucleotides 367–386) and P2 (5′-TTGGGTGATACATACACAAGGG-3′, antisense: nucleotides 891–913) generated a 547bp NAT2 specific fragment spanning the polymorphic sites. Polymorphisms were detected using restriction enzymes KpnI, DdeI, TaqI, and BamHI. PCR was performed on a Hybaid Omnigene™ thermal cycler using 200 ng genomic DNA, 200 ng of each primer, 200 mM dNTPs (Pharmacia, UK), ×1 polymerase buffer (Promega, UK), 1.5 mM MgCl2, 4% DMSO, and 2 units of Taq polymerase (Promega) in a total volume of 100 μl. Twenty microlitres of each PCR reaction was digested separately with 5 units of restriction enzymes KpnI, DdeI, TaqI, and BamHI (Gibco BRL, Paisley, UK). Digested PCR products were separated by size on 1.8% Metaphor™ (Hoeffer Scientific Ltd) agarose gel. Bands were visualised with ethidium bromide staining and ultraviolet illumination. For each case NAT2 LOH was assessed at all four polymorphic sites.
DATA ANALYSIS
Genotype distributions of the colon cancer and control groups were compared using the χ2 test and Fisher’s exact test where appropriate. Allele frequencies for each group were calculated and genotype distribution for cancers and controls were tested for goodness of fit with the Hardy-Weinberg equilibrium.33 Any deviations from this equilibrium were calculated using a χ2test.
Results
COLORECTAL CANCER STUDY PATIENTS
NAT2 genotype was examined in 275 patients with primary sporadic colorectal cancer. These cancers were staged according to Dukes’s classification: 32 stage A, 127 stage B, and 116 stage C. The group comprised 135 (49%) women and 140 (51%) men, with an average age of cancer onset of 70 years (range 28–95 years). In 65 patients both constitutional and tumour DNA was examined. Histological examination of colorectal cancer tissue confirmed the absence of non-neoplastic epithelial tissue. Non-neoplastic stroma represented less than 5% of the total amount of tissue extracted.
PCR ASSAY FOR NAT2 GENOTYPE
Using previously described restriction fragment length polymorphisms in the NAT2 gene we designed PCR primers which amplified a 547bp fragment of the gene spanning the four major polymorphic sites thus simplifying analysis and reducing reaction numbers. Digestion of PCR products with KpnI, DdeI, TaqI, and BamHI produced diagnostic restriction fragments which identified each of six common alleles in Caucasians, NAT2*4, NAT2*5 (A, B, and C), NAT2*6B, and NAT2*7A according to the combined restriction pattern of the four enzymes (fig1 and table 1). The consolidated classification system was used for NAT2 alleles.34 All restriction fragments corresponded to predicted sizes. Alleles NAT2*5 (A, B, and C), NAT2*6B, and NAT2*7A all contain base changes which result in slow acetylation phenotypes, therefore NAT2 phenotype could be predicted from genotype (table 2). NAT2*4 homozygotes conform to wild type fast, NAT2*4/ NAT2*5, 6, or 7 heterozygotes to intermediate, and NAT2*5, 6, or 7 homozygotes to slow acetylators.35 The phenotype distribution for colon cancers and controls was not significantly different (χ2=1.902, df=2, p=0.387), and indicated that phenotype was not associated with colon cancer aetiology or development in this heterogeneous cancer group. Allele frequencies were similar in both cancer and control groups (χ2=2.571, df=3, p=0.463; table3). Alleles which formed the NAT2*5 classification, B and the rare alleles A and C, were grouped together to simplify analysis.

: NAT2 PCR products from constitutive DNA of patients with colorectal cancer, following digestion with restriction enzymes: (A) Kpn I; (B) DdeI; (C) TaqI; and (D) BamHI, illustrating four NAT2 polymorphic sites. Band sizes (in base pairs) are given for polymorphic sites (+ and –) and constant bands (c). Restriction patterns for each enzyme are illustrated: homozygous wild type; heterozygous; and homozygous mutant. MkV denotes molecular weight marker.
NAT2 allele identification according to restriction pattern and corresponding enzyme phenotype. + or − signs refer to the presence of restriction sites within the NAT2 gene
NAT2 genotype in 275 patients with colorectal caner and 343 controls. Alleles NAT2*4, NAT2*5(A, B, and C), NAT2*6B, and NAT2*7A correspond to alleles desribed in table 1
NAT2 genotype with clinical features of the colorectal cancer group
NAT2 GENOTYPE IN CANCER AND CONTROL GROUPS
Nine genotypes, of a possible 10, were identified within both cancer and control groups. Homozygous NAT2*7A was not present in either group. The distribution of NAT2 genotype in 275 cancers and 343 controls is shown in table 2. For each group the distribution of genotype is consistent with the Hardy-Weinberg equilibrium (rejected at p<0.05), and implies that genotype frequencies are constant and not affected by mutation and selection within each group. Overall, we found no association between NAT2 genotype and the presence of colorectal cancer (χ2=5.94, df=3, p>0.05).
Predicted fast and intermediate NAT2 phenotype classifications were combined for subsequent analysis of NAT2 genotype with sex, age of onset, site of cancer, and Dukes’s stage (table 3). Patients who developed colorectal cancer before the age of 70 were more frequently slow acetylators, with a relative risk of 1.69 (95% confidence intervals 1.1–2.6, p=0.018). NAT2 phenotype showed no association with sex (χ2=3.458, df=1, p=0.063), or Dukes’s stage (χ2=0.33, df=2, p=0.848). Cancers on the right side of the colon did not differ in genotype distribution from those on the left (χ2=1.784, df=1, p=0.182), or from the control group (χ2=0.006, df=1, p=0.941).
NAT2 AND LOSS OF HETEROZYGOSITY AT 8P
We investigated NAT2 polymorphisms in 65 colorectal cancers with loss of one or more polymorphic markers on 8p in order to ascertain whether the genotype changes during tumour development. Forty eight cases were informative for the NAT2 gene and in only three (6%) was there evidence that the predicted loss had occurred. NAT2 loss was uncommon, even in individual cases where markers both proximal and distal to the NAT2 locus at 8p22 showed LOH. Allele loss in these cases was determined in a previous study using 12 genomic probes,25 and the complex pattern of loss on 8p is illustrated in fig 2. Figure 3 demonstrates NAT2 allele loss, and retention in two patients with colorectal cancer known to have 8p LOH.
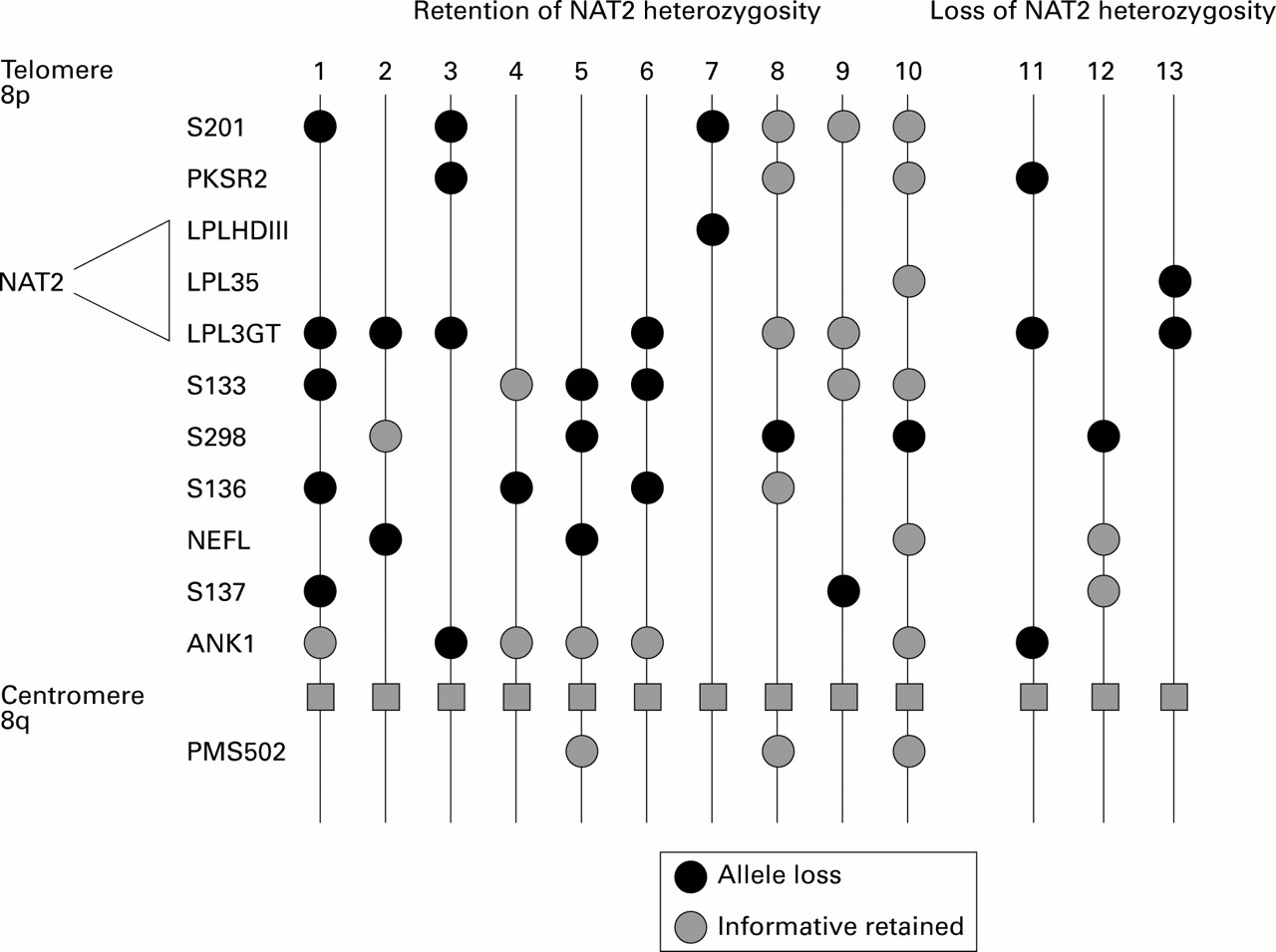
: Heterozygosity of NAT2 was retained in 94% of cancers which had demonstrated LOH on chromosome 8p in a previous study of colon cancer. Gene markers used in that study are listed according to approximate chromosomal location from telomere to centromere and heterozygosity status was examined using constitutive and tumour DNA in PCR and Southern blotting studies.25 The approximate region to which NAT2 maps is indicated on the left of the figure. The pattern of allele loss in 13 patients with colorectal cancer is shown. In patients 1–10 NAT2 heterozygosity was retained, and in patients 11–13 NAT2 heterozygosity was lost.

{kind=link}
{kind=link}
{kind=link}
: NAT2 alleles of two patients with colorectal cancer which had previously shown LOH on chromosome 8p. Patient A demonstrates allele retention and patient B allele loss. Restriction digests for each enzyme, KpnI, DdeI, TaqI, and BamHI are shown for matched pairs of constitutional DNA (N) and tumour DNA (T). Arrows indicate alleles showing loss.
Discussion
We have examined polymorphisms of the NAT2 gene in constitutive DNA derived from patients with colorectal cancer and from a control group of normal individuals. Overall, we found no association between the NAT2 genotype and colorectal cancer risk in patients aged over 69, with sex, site of tumour, or Dukes’s stage. However, in patients who had developed colorectal cancer before the age of 70, there was a significant increase in risk associated with a slow NAT2 genotype. As changes in NAT2 activity may alter carcinogen metabolism within tumour cells we examined the NAT2 locus for deletion. In only 6% (3/48) of colorectal cancers with loss of heterozygosity at chromosome 8p was there a change in the inferred NAT2 phenotype in the tumour.
The fast and slow acetylation phenotypes of NAT2 have both been associated with cancer aetiology,5 ,9-11 indicating that activation of, or failure to deactivate arylamine carcinogens may be important in different cancer types. In agreement with other biochemical and genetic studies,24 ,36 we failed to observe any overall association between NAT2 genotype and colorectal cancer risk. It is possible that an activation of one or more carcinogens by fast NAT2 O-acetylation ofN-hydroxy metabolites can be important in colorectal cancer aetiology.5 ,10 Unfortunately, these latter studies only analysed the NAT2 phenotype in small numbers (less than 50 patients) of colorectal cancer cases, wherein some bias may have occurred. Furthermore, the sensitivity of the biochemical test used and substrate specificity are confounding factors in determination of NAT2 phenotype and may have contributed to the apparent disparity between some studies. Attempts to pool data from these studies37should be viewed cautiously as differences in experimental technique, population base, and diet, particularly from disparate countries, may have influenced the outcome. An additional variable is the activity of NAT1. This shows a degree of functional overlap with NAT2 in terms of substrate specificity. This gene was previously thought to be monomorphic, but it is now clear that it also has functional polymorphism.38
Comparative studies have clearly indicated that NAT2 phenotype can be accurately predicted from its genotype.35 Predicted slow NAT2 acetylator genotypes were more common in colorectal cancer patients under 70 years of age than in controls (odds ratio 1.69), suggesting that slow acetylation is a risk factor for cancers in younger people and hence that fast acetylation is protective. This is a role similar to that described in bladder cancer, but for that disease phenotype did not vary with age in either the bladder cancer or control groups.9 Although variation in allele frequency with age is known to occur in some genes in normal populations,39studies of NAT2 phenotype have indicated that this does not occur for this gene.24 Another cytosolic enzyme, glutathione S-transferase T1, has also shown differences in its genotype distribution with age of onset of colorectal cancer.8 As the process of colorectal carcinogenesis may take a number of years, this suggests that NAT2 polymorphism may increase the sensitivity to initial genotoxic insult and increase the rate of tumour progression.
Previous reports have suggested that polymorphisms of xenobiotic enzymes in the colon can affect the distribution and location of tumours.7 ,8 The acetylation activity of NAT2 enzyme has been demonstrated in colonic cytosols,3 and remains a potentially important regulator of genotoxic insult to colonic crypt cells. However, we failed to find any strong association between NAT2 genotype and cancer site.
Repeated exposure to carcinogens produces additional genetic lesions which contribute to tumour progression. We examined the possibility that changes in NAT2 may reflect altered sensitivity to xenobiotics during tumorigenesis. The NAT2 gene was retained in 94% of colon cancers which showed allele loss at chromosome 8p. This is surprising as the NAT2 gene has been mapped to 8p22,40 ,41 which overlaps the region of chromosome 8 which is most frequently deleted in colorectal cancer.25-27 The location of NAT2 has been clearly established at 8p22, telomeric to the LPL loci, using somatic cell hybrids, and by cosmid cloning.40 ,41 Independent studies have suggested that there are at least two tumour suppressor genes present on chromosome 8p at 8p23.2-8p22 and 8p21.3-8p11.22.27-29 Loss of both these sites in an individual cancer was common,27 indicating significant loss of chromosome 8p. The precise order of loci used in mapping chromosome 8 has not been fully determined, therefore the patterns of loss illustrated in fig 2 will be refined when new mapping information is available. Despite this complex pattern of allele loss on 8p, the retention of NAT2 in some cancers suggests either that some acetylator activity may be essential for cell survival or that NAT2 may be linked to a gene which is essential for maintenance of cell viability. The complex patterns obtained in our analysis of 8p deletions raise the possibility of its translocation to another chromosome. Such events are common in colorectal cancer.
NAT2 genotype may be a risk factor in cancers arising in younger people. That the increased risk is small reflects the polygenic nature of colorectal cancer and the functional redundancy of many xenobiotic metabolising enzymes. NAT2 is situated within a chromosomal region involved in complex rearrangements in colorectal cancer and tends to be retained. It may prove to be a useful marker for further investigation of genes present at 8p involved in regulation of cell growth.
Acknowledgments
Some data in this paper were presented to the Pathological Society of Great Britain and Ireland (J Pathol 1995;176 (suppl): 18A). This study was supported by Cancer Research Campaign grant number SP2288. CM was an SOHHD Summer Vacation Student.