Article Text

Abstract
Background—Increased production of reactive metabolites of oxygen and nitrogen has been implicated in chronic inflammation of the gut. The object of this study was to examine the magnitude and location of nitric oxide synthase (NOS) activity and peroxynitrite formation in the colonic mucosa of patients with ulcerative colitis in relation to the degree of inflammation.
Subjects—Thirty three patients with active ulcerative colitis (17 with mild or moderate inflammation, 16 with severe inflammation).
Methods—Inducible NOS activity was determined in the colonic mucosa by measuring the conversion ofl-arginine to citrulline in the absence of calcium. The localisation of NOS and nitrotyrosine immunoreactivity was assessed immunohistochemically using the labelled streptavidin biotin method.
Results—Inducible NOS activity increased in parallell with the degree of inflammation of the mucosa. Expression of inducible NOS was found not only in the lamina propria, but also in the surface of the epithelium. Peroxynitrite formation as assessed by nitrotyrosine staining was frequently observed in the lamina propria of actively inflamed mucosa.
Conclusions—Nitric oxide and peroxynitrite formation may play an important role in causing irreversible cellular injury to the colonic mucosa in patients with active ulcerative colitis.
- nitric oxide
- peroxynitrite
- nitric oxide synthase
- ulcerative colitis
- colonic mucosa
Statistics from Altmetric.com
Ulcerative colitis (UC) is characterised by recurrent diffuse inflammation which is confined to the colon and rectum. Although the precise aetiology and pathogenesis of the disease remains to be elucidated, there is a growing body of both experimental and clinical evidence to suggest that chronic gut inflammation is associated with enhanced production of reactive metabolites of oxygen and nitrogen.1-9 It is thought that, along with reactive oxygen metabolites, NO-derived metabolites contribute greatly to the tissue dysfunction produced by chronic inflammation.
NO, one of the smallest biologically active molecules, is formed froml-arginine by the enzyme nitric oxide synthase (NOS) and there are two major classes of NOS isoenzymes10 ,11 termed constitutive (calcium-dependent; cNOS) and inducible NOS (calcium-independent; iNOS). cNOS is present primarily in endothelial cells and neuronal cell bodies.12 It produces a small amount of NO and is thought to be important in the maintenance of homoeostasis of cardiovascular and nervous systems.10iNOS, on the other hand, is present in various types of cell, including phagocytic leucocytes, and can be induced by a variety of different proinflammatory cytokines and mediators as well as by certain bacterial products.12 ,13 iNOS produces much more NO and is thought to be important in the host defence against intracellular pathogens and tumour cells.14 ,15 However, high NO concentrations may be toxic and damage normal tissues. Tissue injury may result from a combination of NO with superoxide anion, both of which are actively produced in inflamed tissue, yielding the highly toxic species, peroxynitrite.16
It has been increasingly shown that chronic inflammation of the intestine and colon can be ameliorated by the inhibition of NOS in experimental animals. Miller et al 1 have demonstrated that chronic ileitis induced by trinitrobenzenesulphonic acid can be ameliorated by the inhibition of NOS in guinea pigs. Similarly, Grisham et al 5 showed that chronic inhibition of NOS byl-arginine monomethyl ester or aminoguanidine attenuated significantly the peptidoglycan/polysaccharide induced increases in macroscopic colonic inflammation scores and colonic myeloperoxidase activity in rats. Thus it is conceivable that NO synthesis is augmented in chronic inflammation of the intestine, influencing the disease process. In human rectal tissues, an increase in concentrations of citrulline, a coproduct of NOS activity, and enhanced colonic NO generation and NOS activity have been reported in patients with active UC.2 ,8
However, the sources for this enhanced NO generation have not been fully identified. There is some evidence that the phagocytic leucocytes (neutrophils, monocytes, macrophages), which are known to accumulate within the inflamed mucosa of patients with UC, may represent an important source of reactive nitrogen metabolites. Miles et al 17 showed that extravasated but not circulating rat neutrophils produced much larger amounts of NO by the upregulation of both iNOS mRNA and iNOS enzyme activity. Intestinal or colonic inflammation in human inflammatory bowel disease is known to be associated with increases in tissue-derived cytokines, such as tumour necrosis factor, interferon-γ, and interleukin 1.18 Some of these cytokines are potent inducers of NO synthase in macrophages, neutrophils, and endothelial cells. On the other hand, cultured intestinal epithelial cells have been reported to release a large quantity of NO in response to endotoxin and a variety of cytokines.19 ,20 It was noted that iNOS expression by epithelial cells was associated with diminished cell viability, suggesting that NO, possibly via peroxynitrite formation, may lead to irreversible cellular injury.
The objective of the present study is to assess the role that NO may play in mediating colonic inflammation in active UC by assessing the site, enzyme source, and magnitude of NO production. Moreover, as an indicator of peroxynitrite formation through NO, nitration of tyrosine was assessed immunohistochemically and the results were compared with the localisation of iNOS.
Methods
PATIENTS
Thirty three patients with active UC (24 men and 9 women; mean age 37.3 years; range 18–57 years) were enrolled in the study. UC was diagnosed on the basis of standard diagnostic criteria, and disease activity was calculated as described in a previous report.21 Namely, scores were given on the basis of frequency of bowel movement, body temperature, pulse rate, presence of blood in the stool, and abdominal pain. The scores ranged from 0 to 8. Patients with scores above 3 were regarded as in the active stage. Disease activity was moderate or severe in 16 patients and mild in 17. Twelve patients had proctosigmoiditis, 13 had left sided colitis, and eight had extensive disease. Medications being taken at the time of colonoscopy were: prednisolone (10–30 mg three times a day) in 16 patients; sulphasalazine (2–6 g three times a day) in 26 patients; and 6-mercaptopurine (20–30 mg three times a day) in eight patients. Three were receiving topical corticosteroids. Thirteen patients received a combination of these at the time of colonoscopy.
The control group comprised 18 patients (ten men and eight women; mean age 40.8 years) with colonic polyps. Biopsy specimens of their colonic mucosa were histologically normal.
Patients underwent colonoscopy after standard bowel preparation. In each patient with UC, colonic biopsy specimens were taken from both endoscopically inflamed and endoscopically normal mucosa. All subjects gave their informed consent and the study was approved by the ethical committee of our university.
DETERMINATION OF iNOS ACTIVITY IN INTESTINAL MUCOSA
iNOS activity in biopsy specimens was determined by monitoring the conversion of l-arginine to citrulline in the presence of l-arginine analogue and in the absence of calcium.22 Briefly, tissues were homogenised in 50 mM Tris buffer (pH 7.0) containing 1 mM EDTA, 1 mM dithiothreitol, 10 μg/ml leupeptin, and 0.2 mM phenylmethanesulphonyl fluoride. The homogenates were centrifuged at 30 000 rpm and the supernatant and pellet were saved on ice. The supernatant was assayed using the N G-monomethyl-l-arginine (L-NMMA; Sigma, St Louis, MO, USA)-inhibitable conversion of14C-labelled l-arginine to [14C]citrulline. Small aliquots (0.05 ml) of the supernatants were added to the reaction mixture containing 50 mM Hepes (pH 7.2), 1.0 mM EGTA, 1.0 mM MgCl2, 10 μMl-[14C]arginine, 1 mM citrulline (to inhibit catabolism of [14C]citrulline), 10 mMl-valine (to inhibit non-specific arginase activity), 1 μM FAD, and 0.1 mM NADPH to a total volume of 200 μl. The reaction mixture was incubated for 60 minutes at 37°C. After completion of the incubation period, 1 ml of a 1:1 suspension of Dowex (AG 50W-8) and buffer was added to bind all unchanged radiolabelledl-arginine. Formation of citrulline in the presence of excess L-NMMA (1 mM) was also determined in order to be able to subtract background citrulline formation.
IMMUNOPRECIPITATION
Equal amounts of protein from each homogenised sample were treated with the detergent Nonidet P40 (1%) and briefly sonicated. The solubilised protein (500 μg) was then incubated with monoclonal antibodies against iNOS (macNOS) (mouse IgG2a; Transduction Laboratories) (5 μg) for ten hours at 4°C in a total volume of 500 μl of immunoprecipitation buffer (10 mM Tris/HCl, pH 7.4, 1% Nonidet P40, 1 mM EDTA, 0.5% BSA, and 0.2 mM phenylmethanesulphonyl fluoride). After addition of 5 μg of rabbit anti-mouse immunoglobulin G antibody, the incubation mixture was vortex-mixed and the incubation continued for an additional hour at 4°C. Then 100 μl of a 10% suspension of fixed Staphylococcus aureus (Zymed Co., South San Francisco, CA, USA) was added to the mixture, which was incubated for 30 min and centrifuged at 2500 g for 20 minutes. iNOS activity remaining in the supernatant was determined as described above.
HISTOLOGICAL AND IMMUNOHISTOCHEMICAL EXAMINATION FOR NO SYNTHASE AND NITROTYROSINE
Biopsy specimens from each patient were fixed in 10% buffered formalin and embedded in paraffin after hydration. Serial 4 μm sections were stained with haematoxylin and eosin. The histological scores reflect the degree of inflammatory cell infiltration and tissue destruction as assessed on a five point scale corresponding to that described by Riley et al 23: 1, normal; 2, mild increase in chronic inflammatory cell infiltrate, with no tissue destruction; 3, moderate increase in chronic inflammatory cell infiltrate, with no tissue destruction; 4, marked increase in chronic inflammatory cell infiltrate, with mild tissue destruction; and 5, marked increase in chronic inflammatory cell infiltrate, with obvious tissue destruction.
NOS activity and nitrotyrosine were assessed immunohistochemically using the labelled streptavidin biotin method. Biopsy specimens from each patient were fixed in periodate/lysine/paraformaldehyde solution, and embedded in “optimum cutting temperature” (OCT) compound (Miles Inc, Elkhart, IN, USA) before being frozen in dry ice and acetone. Cryostat sections 6 μm thick were transferred to gelatin coated slides and air dried for one hour at 20°C. After they had been washed in phosphate buffered saline, pH 7.4 (PBS) containing 1% Triton X-100 for five minutes, sections were incubated in 1% normal goat serum in PBS. Monoclonal antibodies against iNOS (macNOS) and nitrotyrosine (mouse immunoglobulin G; Update Biotechnology Inc, Lake Placid, NY, USA) were diluted 50 to 100 times with PBS and layered on the section overnight at 4°C. Sections were incubated with secondary antibody (biotinylated anti-mouse immunoglobulin G class antibody; Amersham International plc, Amersham, Bucks, UK) for one hour at 37°C. Then sections were incubated with fluorescein isothiocyanate conjugated streptavidin (Streptavidin-fluorescein; Amersham International plc) for 30 minutes at room temperature. A rinse with PBS containing 1% bovine serum albumin was performed at each step. A coverslip was applied using glycerol jelly. These sections were observed under a fluorescence microscope (Olympus, Tokyo, Japan). As negative controls, sections were treated with the same procedure except for the absence of primary antibodies.
WESTERN BLOT ANALYSIS
Immunoblotting experiments using antibodies used for immunofluorescence staining were performed to confirm the specificity of antibodies. Biopsy specimens were homogenised using a 15 ml type B blender homogeniser at 2000 rpm with ten complete strokes in five volumes of boiling lysis buffer (1% sodium dodecyl sulphate, 10 mM Tris/HCl, pH 7.4) and microwaving for 10–15 seconds. Aliquots (100 μg protein) of the sample were resolved by sodium dodecyl sulphate/polyacrylamide gel electrophoresis (12% gel (Protogel); National Diagnostics, Atlanta, GA, USA). The protein was then immobilised on nitrocellulose membrane by electrotransfer. The membrane was incubated with 3% non-fat dried milk for 20 minutes at room temperature to block the non-specific binding sites. The membrane was then incubated with monoclonal antibodies directed against iNOS overnight with agitation at 4°C and then with colloidal gold-conjugated secondary antibodies (goat anti-mouse immunoglobulin G; 1:200 dilution) in PBS/3% non-fat dried milk for 30 minutes at room temperature with agitation. Membrane bound antibodies were detected using a silver enhancing kit (Bioclin, UK).
STATISTICAL ANALYSIS
All results are expressed as means and SEM. Differences among groups were evaluated using one way analysis of variance and Fisher’s post-hoc test.
Results
To determine whether iNOS antibody can detect iNOS protein specifically, Western blotting with anti-iNOS antibody was performed on tissue from normal colon and tissue from a region of active UC (fig 1). In the normal colon, iNOS was not detected (lane a). In the active UC mucosa with severe inflammation (grade 4), anti-iNOS antibody labelled a single major band which migrated at the 130 kDa position (lane b), whereas only a weak band was detected in the mucosa with mild inflammation (grade 2) (lane c).
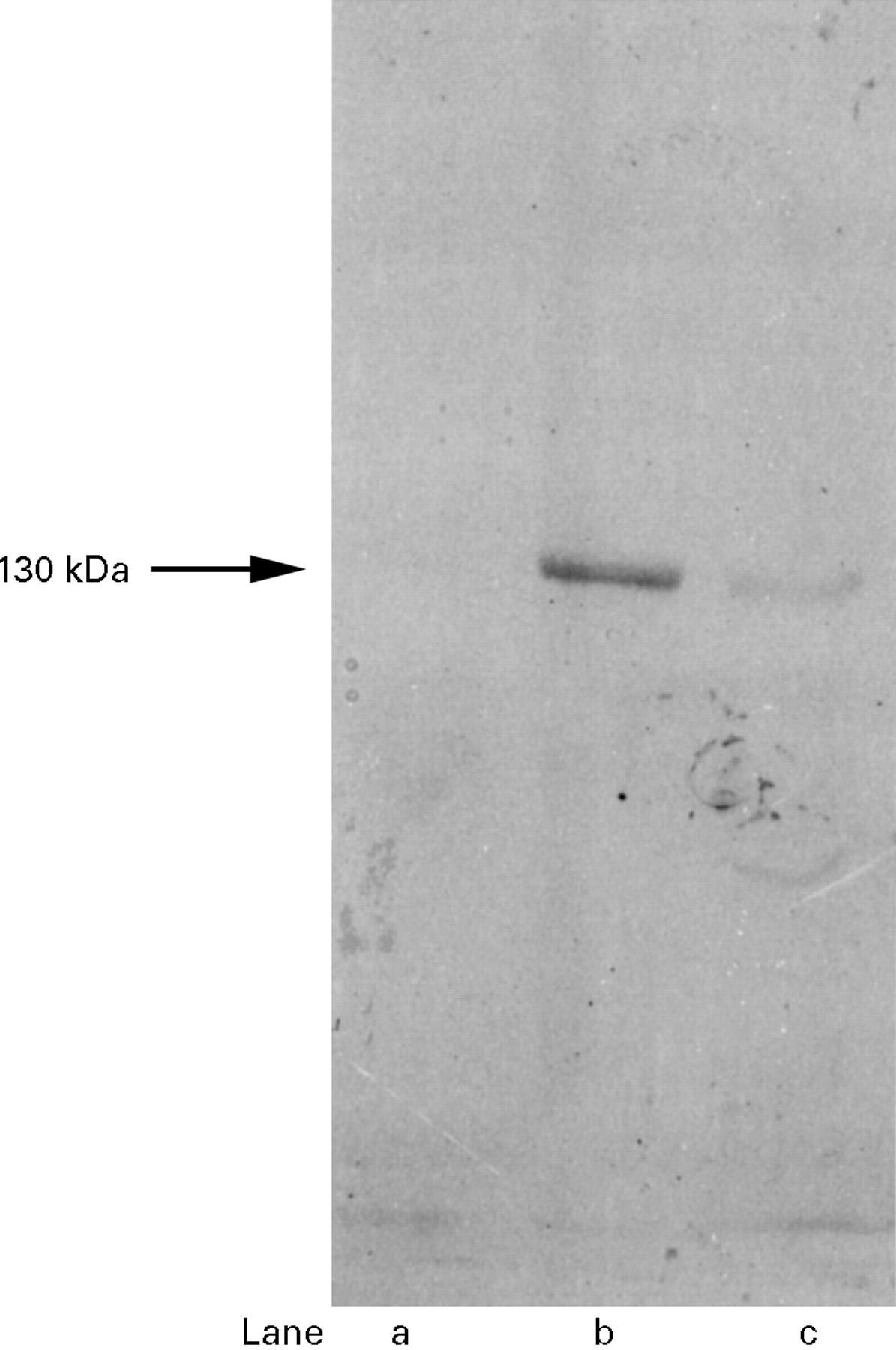
Detection of inducible nitric oxide synthase (iNOS) protein in the biopsy specimen of colonic mucosa by Western blotting with anti-iNOS antibody. Lane a, iNOS was not detected in homogenate of histologically normal mucosa from a control subject. Lane b, in the active UC mucosa with severe inflammation (grade 4), anti-iNOS antibody labelled a single major band which migrated at the 130 kDa position. Lane c, only a weak band was detected in the UC mucosa with mild inflammation (grade 2).
Figure 2 illustrates colonic NOS activity in patients with UC. In control subjects there was no detectable iNOS activity in the colonic mucosa. Biopsy specimens from endoscopically normal mucosa of UC patients showed iNOS values of less than 10 pmol/h per mg protein, while iNOS activity was significantly increased in actively inflamed mucosa compared with quiescent mucosa. iNOS activity of the diseased mucosa from patients with active UC correlated well with the severity of inflammation. Colonic iNOS activity determined in mucosa of histological grade 4 or 5 was significantly higher than that in mucosa with grade 2 or 3 inflammation. iNOS activity was less than 5% of that of both inflamed and quiescent mucosa in the immunoprecipitated subjects, suggesting that monoclonal antibody against iNOS actually reacts with iNOS protein.

Inducible nitric oxide synthase (iNOS) activity in colonic mucosa. iNOS activity was determined in biopsy specimens from endoscopically normal (quiescent) mucosa and actively inflamed mucosa by monitoring the conversion of l-arginine to citrulline. Enzyme activities associated with mild or moderate inflammation (grade 2 or 3) and severe inflammation (grade 4 or 5) were compared. Values are expressed as means and SEM. *p<0.05 compared with quiescent mucosa. †p<0.05 compared with colonic mucosa with grade 2 or 3 inflammation.
Immunohistochemical study of iNOS in colonic tissue from patients with UC showed the presence of iNOS activity in the lamina propria as well as in the superficial epithelium. In control subjects, positive iNOS immunoreactivity was not detected in either the surface epithelium or the lamina propria. Table 1 shows the rate of positive iNOS staining in colonic tissues in patients with active UC. Localisation of iNOS activity was compared in patients with mild or moderate inflammation (grade 2 or 3) and those with severe inflammation (grade 4 or 5). Only 54% of patients with grade 2 or 3 inflammation showed positive iNOS activity. In these patients, iNOS activity was observed mainly in the lamina propria and only 19% exhibited iNOS activity in the surface epithelial cells. In the apparently normal mucosa of these patients, positive iNOS activity was found in the lamina propria in only one case. On the other hand, there was a considerable difference in iNOS distribution in the colonic mucosa of patients with severe inflammation (grade 4 or 5): iNOS activity was observed in all of the inflamed mucosa. It should be noted that, in the inflamed mucosa of grade 4 or 5, a significant increase was demonstrated in positive iNOS reactivity in the surface epithelium of the colonic mucosa as well as in the lamina propria as shown in fig 3(19% in grade 2 or 3 v 81% in grade 4 or 5; p<0.05). In two patients with actively inflamed mucosa of grade 4 or 5, positive iNOS reactivity was confined to the epithelium, as shown in fig 4. Even in patients with grade 4 or 5 inflammation, iNOS activity was not detected in apparently normal mucosa except in one case.
Localisation and positive rate of immunoreactivity of inducible nitric oxide synthase in colonic tissues in patients with active ulcerative colitis

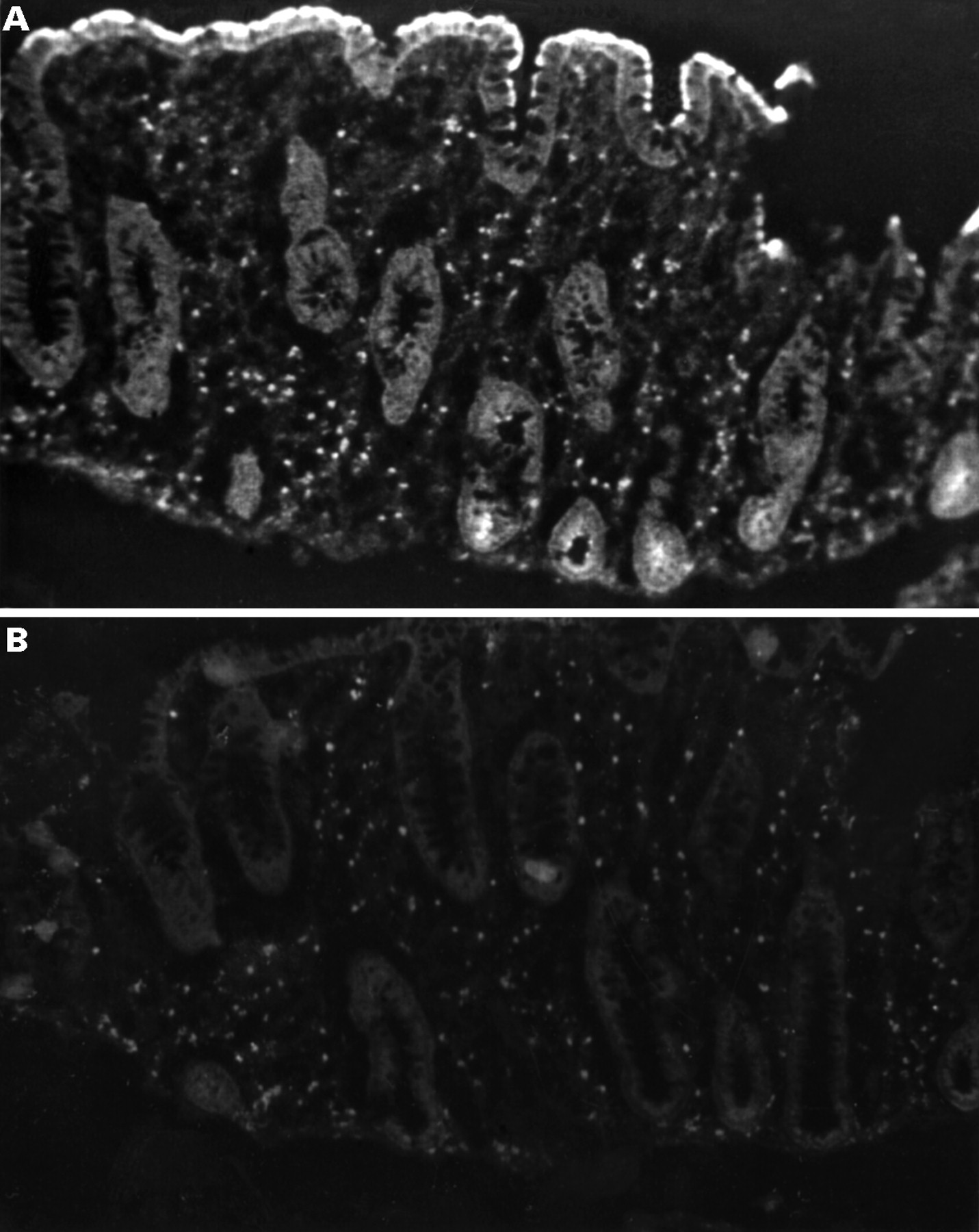
Immunohistochemical study of inducible nitric oxide synthase (iNOS) activity in actively inflamed ulcerative colitis mucosa. Localisation of iNOS immunoreactivity was examined using the labelled streptavidin biotin method with antibodies against iNOS (macNOS). The biopsy specimen was obtained from actively inflamed mucosa grade 4. (A) Significant iNOS reactivity was demonstrated in the surface epithelium of the colonic mucosa as well as in the lamina propria. (B) A negative control picture without primary antibody is also shown.(Original magnification × 100.)

Immunohistochemical study of inducible nitric oxide synthase (iNOS) activity in actively inflamed ulcerative colitis mucosa. Localisation of iNOS immunoreactivity was examined using the labelled streptavidin biotin method employing antibodies against iNOS (macNOS). The biopsy specimen was obtained from actively inflamed mucosa with grade 5 inflammation. In this patient, iNOS reactivity was confirmed in the surface epithelium of the colonic mucosa (A). A negative control picture without primary antibody is shown (B). (Original magnification × 400.)
Figure 5 shows the relation between iNOS activity and the site of iNOS immunoreactivity in the inflamed colonic mucosa of patients with active UC. Mucosa exhibiting positive iNOS immunoreactivity showed significantly higher iNOS activity than did mucosa exhibiting no immunoreactivity. In addition, the inflamed mucosa that exhibited iNOS immunoreactivity in both the lamina propria and the surface epithelial cells showed a significantly higher iNOS enzyme activity than mucosa in which iNOS activity was confined to the lamina propria.

Relation between inducible nitric oxide synthase (iNOS) enzyme activity and the site of iNOS immunoreactivity in the inflamed colonic mucosa of patients with active ulcerative colitis. Values are expressed as means and SEM. *p<0.05 compared with colonic mucosa without iNOS immunoreactivity. †p<0.05 compared with the colonic mucosa with positive iNOS immunoreactivity observed in both lamina propria and epithelial cells.
Table 2 shows the rate of positive nitrotyrosine staining in colonic tissues in patients with active UC. Nitrotyrosine staining of colonic tissue indicates the presence of tyrosine nitration and was compared in patients with mild or moderate inflammation (grade 2 or 3) and severe inflammation (grade 4 or 5). In the control subjects, positive nitrotyrosine immunoreactivity was not detected in either the surface epithelium or the lamina propria. In patients classified as having grade 2 or 3 inflammation, positive nitrotyrosine activity was observed in 41%, always in the lamina propria.
Localisation and positive rate of nitrotyrosine immunoreactivity in colonic tissues in patients with active ulcerative colitis
Figure 6 shows typical positive nitrotyrosine immunoreactivity in the lamina propria of colonic mucosa from patients with grade 4 inflammation. In patients classified as grade 4 or 5, we were able to find positive nitrotyrosine reactivity in all of the biopsy specimens from the inflamed mucosa. Some 75% of this nitrotyrosine activity was found only in the lamina propria of the intestine, whereas the others also had nitrotyrosine activity in the surface epithelium, as shown in fig 7.

Immunohistochemical study of nitrotyrosine activity in actively inflamed ulcerative colitis mucosa. Localisation of nitrotyrosine immunoreactivity was examined using the labelled streptavidin biotin method with antibodies against nitrotyrosine. The biopsy specimen was obtained from actively inflamed mucosa of grade 4 inflammation. Nitrotyrosine immunoreactivity was observed in the lamina propria of colonic mucosa (A). A negative control picture without primary antibody is shown (B). (Original magnification × 400.)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Immunohistochemical study of nitrotyrosine activity in actively inflamed ulcerative colitis mucosa. Localisation of nitrotyrosine immunoreactivity was examined using the labelled streptavidin biotin method with antibodies against nitrotyrosine. The biopsy specimen was obtained from grade 5 actively inflamed mucosa. In this patient, nitrotyrosine immunoreactivity was observed not only in the lamina propria, but also in the surface epithelium (A). A negative control picture without primary antibody is shown (B). (Original magnification × 400.)
Discussion
Chronic inflammation of the colonic mucosa in patients with active UC has been reported to be associated with enhanced production of NO,2 ,7 ,8 which may play an important role in pathogenetic processes of inflammation induced tissue injury. In the present study, we show that colonic mucosa from patients with UC associated histologically with severe inflammation more often displayed positive immunoreactivity for iNOS than did tissue derived from patients with lower grade inflammation. Enhanced NO production was confined to the actively inflamed mucosa, whereas iNOS immunoreactivity was virtually undetectable in the apparently normal mucosa of these patients. Miller et al 24 have reported that the inclusion of aminoguanidine in the drinking water resulted in dose-dependent inhibition of inflammatory responses in trinitrobenzenesulphonate-induced guinea pig ileitis.24 We hypothesise that enhanced NO synthesis by iNOS in the inflamed intestine may play a key role in further aggravation of the inflammation and may be closely associated with the development of tissue injury.
In our present study, no iNOS immunoreactivity was observed in actively inflamed mucosa from patients with colitis, although in such mucosa iNOS activity was detected by enzymic assays. The reason for this discrepancy is not known. However, it is conceivable that histological examination with a specific antibody for iNOS is less sensitive, and low grade activity cannot be differentiated from non-specific background activity.
The source of the enhanced NO generation in the colonic mucosa has not been exactly identified. However, there is some evidence that phagocytic leucocytes (neutrophils, monocytes, macrophages), which are known to accumulate mainly within the lamina propria of the inflamed mucosa,5 ,25 represent important sources of these potentially injurious free radicals. Endothelial cells, fibroblasts, and mast cells may also contribute to NO generation in inflamed tissues, although much of their activity is generally regarded as being calcium dependent.11 Tepperman et al 19 have shown that iNOS activity can be induced in cultured intestinal epithelial cells in response to endotoxin and various cytokines, and this was observed in the absence of superoxide-producing cells, such as macrophages. Increased NO generation caused by cytokines, bacterial products, and mitogens has also been reported by other groups in cultured small intestinal epithelial cells.20 In concert with these observations, we found in this study that enhanced NO synthesis by iNOS occurs not only in the lamina propria, but also in the superficial epithelium of the inflamed colonic mucosa. It should be noted that expression of iNOS activity in the surface epithelium was increased with disease activity. This increased iNOS expression in the colonic epithelial cells may contribute largely to the enhanced iNOS activity observed in the actively inflamed mucosa of patients with UC and may be associated closely with the diminished cellular viability of the epithelium itself.
The peroxynitrite anion is a potent oxidant formed by a rapid reaction of superoxide with NO.16 ,26 Peroxynitrite reacts with proteins to form a stable product, nitrotyrosine, by addition of a nitro group to the 3’ position adjacent to the hydroxyl group of tyrosine. Although theoretically possible, the in vivo nitration of protein tyrosines by other NO species such as nitrogen dioxide (NO2) is likely to be trivial.27 The marked increase in immunoreactivity of peroxynitrite has been reported in the rat aorta and in the lung during acute endotoxaemia.28 ,29Recently, Kooy et al 30 showed that nitrogen-derived oxidants are formed in human acute lung injuries and suggest that peroxynitrite may be an important oxidant in inflammatory lung diseases. In experimental guinea pig ileitis, Miller et al 24 showed immunohistochemically that nitrotyrosine and iNOS were co-localised and that inihibition of NO formation prevented nitrotyrosine formation. By using specific antibodies to nitrotyrosine residues, we have shown that nitration of protein tyrosines actually occurs in the colonic mucosa of patients with active UC, thereby demonsrating the effects of NO-derived oxidants, most likely peroxynitrite, in the injury of inflamed mucosa of patients with active UC.
The positive rate of nitrotyrosine staining also increased with the increase in disease activity and may correlate with irreversible tissue injury in patients with active UC. However, nitrotyrosine staining was observed mainly in the lamina propria. Since NO itself does not result in nitration of tyrosine, it is reasonable that nitrotyrosine staining is not frequently seen in the surface epithelium, to which superoxide anion from phagocytic leucocytes cannot easily gain access.
It should be pointed out that many of the patients from whom the biopsy specimens were obtained were receiving corticosteroids or salazosulphapyridine preparations, each of which is known to be capable of inhibiting the release of inflammatory mediators. Corticosteroids, in particular, are well known inhibitors of iNOS activity. It is conceivable therefore that our results were biased by such effects. However, we could not find a correlation between iNOS immunoreactivity and the medications taken in any group of patients (data not shown). Further, such therapy would be expected to minimise the inflammatory response, and hence would not account for the enhanced production of NO that we observed.
NO may promote inflammation by virtue of its ability to mediate chemotaxis of both neutrophils and monocytes.31 ,32 Other mechanisms by which NO promotes tissue injury and inflammation include the release of intracellular iron, inhibition of mitochondrial function, and inhibition of DNA synthesis.33-35 Weitzmanet al 36 and Thomas and co-workers37 have shown that non-toxic concentrations of certain leucocyte-derived oxidants are potent mutagens in vitro, suggesting a possible link between chronic inflammation and neoplastic transformation in the distal bowel. In addition, auto-oxidation products of NO, such as N2O3, are potent nitrosating agents and mutagens in vitro.38 ,39 With primary or secondary amines, which are abundantly produced by bacteria at low pH, the luminal condition in UC is important for the formation of N-nitroso compounds.40 The presence of significant NO stress in the colonic mucosa, as shown in this study, may promote malignant transformation in the chronically inflamed colon, possibly through the contribution of NO-dependent nitrosative and oxidative chemistry.
Acknowledgments
This study was supported in part by grants from the Ministry of Education, Science and Culture of Japan and from Keio University.