Article Text

Abstract
Background—Production of chemoattractant factors by the intestinal epithelium may contribute to mucosal infiltration by inflammatory cells in inflammatory bowel disease. Secretion of the α chemokine interleukin 8 (IL-8), a neutrophil chemoattractant, has been widely studied, but little is known about epithelial secretion of β chemokines, which are preferentially involved in recruiting monocytes.
Aims—To investigate the profiles of α and β chemokine secretion in colonic cell lines and their differential modulation by interferon γ (IFN-γ), a product of activated T lymphocytes and natural killer cells.
Methods and results—HT29-19A, a model of the Cl− secretory crypt cell, exhibited a parallel secretion of the α chemokines IL-8 and GROα, which could be markedly upregulated by tumour necrosis factor α (TNF-α) and IL-1β. These cells showed no significant expression of the β chemokines RANTES (regulated upon activation T cell expressed and secreted), MIP-1α (macrophage inflammatory protein 1α), and MCP-1 (monocyte chemotactic protein 1) under these conditions, but IFN-γ in combination with TNF-α caused a dose dependent induction of RANTES and MCP-1 secretion. This was accompanied by a marked increase of RANTES mRNA. In contrast, IFN-γ had no significant effect on TNF-α stimulated IL-8 secretion. Caco-2 cells, with features more typical of villus absorptive cells, were relatively poor secretors of α chemokines but secreted high levels of MCP-1 in response to IL-1β. IFN-γ did not influence α or β chemokine secretion in these cells.
Conclusions—These studies suggest that intestinal epithelial cells may produce chemokines capable of attracting both neutrophils and monocytes. The ability of IFN-γ to activate the expression of β chemokines preferentially could facilitate the development of chronic inflammatory infiltrates.
- inflammatory bowel disease
- RANTES
- interferon gamma
- chemokine
Statistics from Altmetric.com
Inflammatory bowel disease (IBD) is characterised by an increase in the number and activity of inflammatory cells in the gut mucosa. Their physical presence and secreted products contribute to the associated tissue damage and ulceration. Recent evidence has suggested that the gut epithelium may itself be involved in initiating leucocyte recruitment into the mucosa by synthesising and secreting chemokines in response to stimuli in the microenvironment. Studies in cultured epithelia have shown that exposure of cells to both proinflammatory cytokines1 and products derived from common enteric bacteria2 stimulates the synthesis and release of interleukin 8 (IL-8), a chemokine which acts as a powerful chemoattractant for, and activator of, neutrophils. A role for IL-8 in vivo is further indicated by studies showing increased IL-8 in the gut epithelium of patients with IBD.3 ,4
Although IL-8 is implicated in the pathogenesis of IBD it is unlikely to be the sole chemoattractant present in diseased tissue. IL-8 is a member of the α subfamily of chemokines whose members preferentially attract and activate neutrophils. The β subfamily of chemokines, including RANTES (regulated upon activation T cell expressed and secreted), MIP-1 (macrophage inflammatory protein 1), and MCP-1 (monocyte chemotactic protein 1) preferentially recruit and activate mononuclear leucocytes and lymphoid cells, which are also present in elevated numbers in inflamed mucosa. There is evidence that both α and β chemokines can be produced in response to proinflammatory cytokines in cultured pulmonary smooth muscle and endothelial cells5 and bronchial epithelial cells,6 and in response to bacterial invasion of colonic epithelial cells.7 In addition, there is preliminary evidence to suggest that α and β chemokines may be increased in the mucosa of patients with IBD.8 ,9
Interferon γ (IFN-γ) is a product of activated Th1 lymphocytes present in increased amounts in the gut wall of patients with IBD. It has been shown to act synergistically with other proinflammatory cytokines, particularly tumour necrosis factor α (TNF-α), to stimulate chemokine secretion in several systems.5 ,6 ,10 Interferon γ and TNF-α may interact to modulate epithelial barrier function11 but there is little information on the extent to which they modulate the secretion of the different classes of epithelial chemokines.
In this study, the ability of cultured intestinal epithelial cell lines (HT29-19A, Caco-2) to secrete the α chemokines, IL-8 and GROα, and the β chemokines, MCP-1, RANTES, and MIP-1α, in response to inflammatory mediators was investigated. We also examined whether IFN-γ is involved in upregulating chemokine secretion in these cells and whether it has differential effects on α and β chemokines.
Materials and methods
MATERIALS
Mouse anti-IL-8 antibodies were purchased from CLB, The Netherlands. Other chemokine antibodies and Quantikine enzyme linked immunosorbent assay (ELISA) kits were from R&D Systems, Abingdon, UK. All cytokines used were human recombinant peptides. Interleukin 8 was obtained from the National Institute of Biological Standards and Control, South Mimms (Ref. No. 89/520). Interleukin 6, TNF-α, and IFN-γ were from R&D Systems; transforming growth factor (TGF) β and other biochemicals were purchased from Sigma Chemical Co., Poole, UK. Cell culture plastics were obtained from Costar, UK, and media constituents from Gibco BRL, Paisley, Scotland. HT29-19A cells were kindly provided by Dr C Laboisse, INSERM, Paris, and Caco-2 were obtained from the American Tissue Culture Collection. Cell lines were tested periodically for mycoplasma contamination using a commercial ELISA system (Boehringer Mannheim, Lewes, UK).
CELL CULTURE
The HT29-19A (passages 38–52) and Caco-2 (passages 98–110) colonic cell lines were cultured in Dulbecco’s Modified Eagles medium (DMEM) containing 2 mM glutamine, 50 IU/ml penicillin, 50 mg/ml streptomycin, and 10% fetal calf serum. Caco-2 growth medium was supplemented with non-essential amino acids while HT29-19A medium contained 15 mM hydroxyethylpiperazine ethanesulphonic acid (HEPES). For chemokine measurements, HT29-19A or Caco-2 cells were seeded into 24 well tissue culture plates (0.6–1.2×105cells/cm2) and grown for three to seven days. In some experiments, cells were grown on 75 mm microporous filter supports (Transwell, 0.3 μm pore size; 4–8×105cells/cm2) for 10–14 days. Under these conditions, both cell lines form polarised monolayers.12 ,13
CELL STIMULATION
For analysis of chemokine secretion cell layers were washed with growth medium, followed by incubation in fresh medium for 2–24 hours with either cytokines (TNF-α, 0.1–250 ng/ml; IL-1β, 1–250 U/ml; IL-6, 1–50 ng/ml; TGF-β, 0.1–100 ng/ml; IFN-γ, 10–1000 U/ml) or bacterial products (lipopolysaccharide from Escherichia coli 055 (LPS); lipoteichoic acid (LTA); muramyl dipeptide (MDP); N-formyl methionyl leucine phenylalanine (fMLP), 0.1–1 μg/ml). Medium was removed at various times, stored at −70°C, and assayed for α and β chemokines. In experiments with filter grown cells, stimuli (TNF-α, IL-1, IFN-γ) were added to the basolateral compartment and aliquots of both apical and basolateral medium were assayed for chemokines.
CHEMOKINE IMMUNOASSAY
Culture supernatants were analysed for IL-8 by a sandwich ELISA using a commercially available IL-8 monoclonal capture antibody, goat polyclonal second antibody, and a peroxidase conjugated donkey antigoat detection antibody (Jackson Labs, Philadelphia, USA) witho-phenylenediamine (OPD)/H2O2 as substrate. Plates were read at 450 nm on an ELISA plate reader. The detection limit was 80 pg/ml. Analysis of GROα, MCP-1, MIP-1, and RANTES was by commercial sandwich ELISA (R&D Systems, Abingdon, UK) following the manufacturer’s instructions. The limit of detection for these assays was 2–5 pg/ml.
EXPRESSION OF RANTES mRNA
RANTES mRNA expression in HT29-19A was assessed using a reverse transcription polymerase chain reaction (RT-PCR) technique. Total RNA was isolated from cell monolayers using the guanidinium isothiocyanate/phenol/chloroform extraction method of Chomczynski and Sacchi.14 Complementary DNA (cDNA) was synthesised from 2 μg RNA by incubation for 60 minutes at 37°C in a total volume of 20 μl with Moloney murine leukaemia virus reverse transcriptase as part of a commercial kit (Pharmacia Biotech, St Albans). PCR was performed in a final volume of 50 μl containing 1 μl of varying dilutions of the reverse transcription reaction as indicated (equivalent to 1–50 ng RNA), 50 μM deoxynucleoside trisphosphate, 1.5 mM MgCl2, 50 mM Tris-HCl (pH 8.0), 2.5 IU Taq polymerase, and 0.2 μM sense and antisense RANTES primers. The primer sequences used were those published by Snowden et al,15 giving a specific product of 243 bp. PCR was performed in a Techne PHC-3 thermal cycler for 37 cycles consisting of denaturation at 94°C for 0.5 minutes, annealing at 60°C for one minute, and extension at 72°C for one minute. Amplification was terminated by a final extension step at 72°C for five minutes. PCR products were separated by electrophoresis on 2% agarose gels and visualised by ethidium bromide. The integrity of the RNA used was checked by confirming expression of actin mRNA, using actin specific primers16 and a PCR protocol of 94°C for one minute, 60°C for one minute, and 72°C for one minute for 25 cycles followed by a final extension step at 72°C for five minutes.
MONOLAYER INTEGRITY
Transepithelial electrical resistance (Rt), measured using an EVOM dual electrode voltohmmeter (WPI Ltd, Aston, Hertfordshire), was used to assess the integrity of filter grown monolayers prior to and following stimulation with inflammatory agents.
STATISTICAL ANALYSIS
Values are expressed as mean (SEM) for the number of observations indicated. Statistical significance was assessed using Student’st test.
Results
SECRETION OF α AND β CHEMOKINES
Initial studies compared the profile of α and β chemokine secretion in HT29-19A and Caco-2 colonic cell lines. The HT29-19A cell line constitutively secreted low levels of the α chemokines IL-8 and GROα which could be up regulated by several inflammatory agents (fig1A). The most effective activators of secretion were the proinflammatory cytokines TNF-α and IL-1β, which at concentrations of 2 ng/ml and 100 U/ml, respectively, elicited 20–30-fold increases in both IL-8 and GROα secretion over a 24 hour period. Neither IL-6 (0.1–10 ng/ml) nor TGF-β (0.1–10 ng/ml) stimulated significant α chemokine secretion in HT29-19A cells (results not shown).

Secretion of α and β chemokines in (A) HT29-19A and (B) Caco-2 human colonic cell lines. Cultures were incubated for 24 hours with control media alone or in the presence of TNF-α (2 ng/ml in HT29-19A and 50 ng/ml in Caco-2), IL-1 (100 IU/ml), LPS (1 μg/ml), or LTA (1 μg/ml). Values are mean (SEM) of 3–6 observations in each group. *p<0.05; **p<0.01.
A range of bacterial derived products, lipopolysaccharide (LPS), lipoteichoic acid (LTA), muramyl dipeptide (MDP), and fMet-Leu-Phe (fMLP) were also tested for their ability to stimulate secretion. Incubation with LPS (isolated from Gram negative bacteria) or LTA (from Gram positive bacteria) at a concentration of 1 μg/ml caused a two- to fourfold increase in both IL-8 and GROα secretion (fig 1A). Increasing the concentration of LPS and LTA to 10 μg/ml did not induce further increases in α chemokine secretion. Similar results were obtained using MDP and fMLP (data not shown). HT29-19A cells did not secrete appreciable levels of the major β chemokines, RANTES and MCP-1, either constitutively or following exposure to bacterial products or proinflammatory cytokines at the concentrations indicated (fig 1A). Higher concentrations of TNF-α (up to 50 ng/ml) failed to elicit significant secretion of RANTES but stimulated a low level of MCP-1 secretion (45 pg/ml after 24 hours). There was no evidence for expression of MIP-1α by HT29-19A cells.
Caco-2 cells exhibited a markedly different pattern of chemokine secretion. Constitutive production of the α chemokines, IL-8 and GROα, was fivefold lower than observed in HT29-19A cells (fig 1B). In this cell line, IL-1β (100 IU/ml) was the only agent capable of significantly increasing chemokine secretion. Neither TNF-α (50 ng/ml) nor bacterial products (up to 10 μg/ml) stimulated significant secretion of IL-8 or GROα. In contrast to low α chemokine secretion, Caco-2 cells secreted significant amounts of the β chemokine MCP-1. Constitutive secretion of 0.12 ng/ml MCP-1 over a 24 hour period could be upregulated 20–30 fold by 100 IU/ml IL-1. TNF-α (50 ng/ml) also activated MCP-1 secretion causing a sixfold increase over control. Incubation with bacterial products produced no detectable increase in MCP-1 secretion. There was no evidence for significant MIP-1α or RANTES expression, either constitutively or following exposure to TNF-α, IL-1, or bacterial products.
Dose response curves for IL-1β stimulation of IL-8 and GROα secretion in the two cell lines confirm that Caco-2 are relatively poor secretors of α chemokines compared with HT29-19A (fig 2). In each cell line IL-1β was equally effective in stimulating IL-8 and GROα secretion, suggesting that the release of these chemokines is likely to be through a common pathway. IL-1β activated secretion of the β chemokine MCP-1 over a similar dose range in Caco-2 (results not shown).
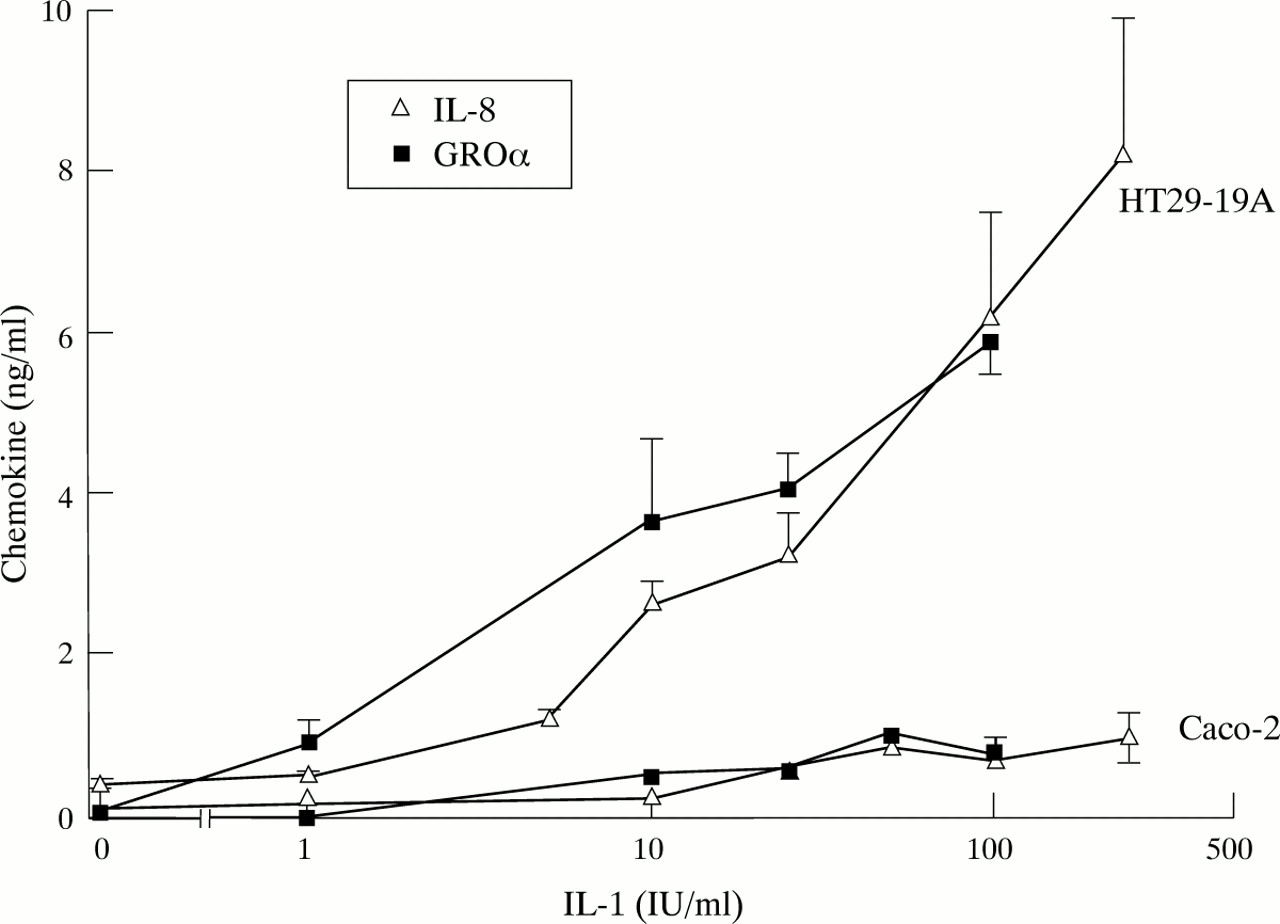
Dose dependent stimulation of α chemokine secretion by IL-1 in HT29-19A and Caco-2 cells. Chemokine levels were measured 24 hours after addition of IL-1 to confluent cultures. Values are mean (SEM) for 3–8 observations in each group.
SYNERGISTIC EFFECTS OF IFN-γ AND TNF-α
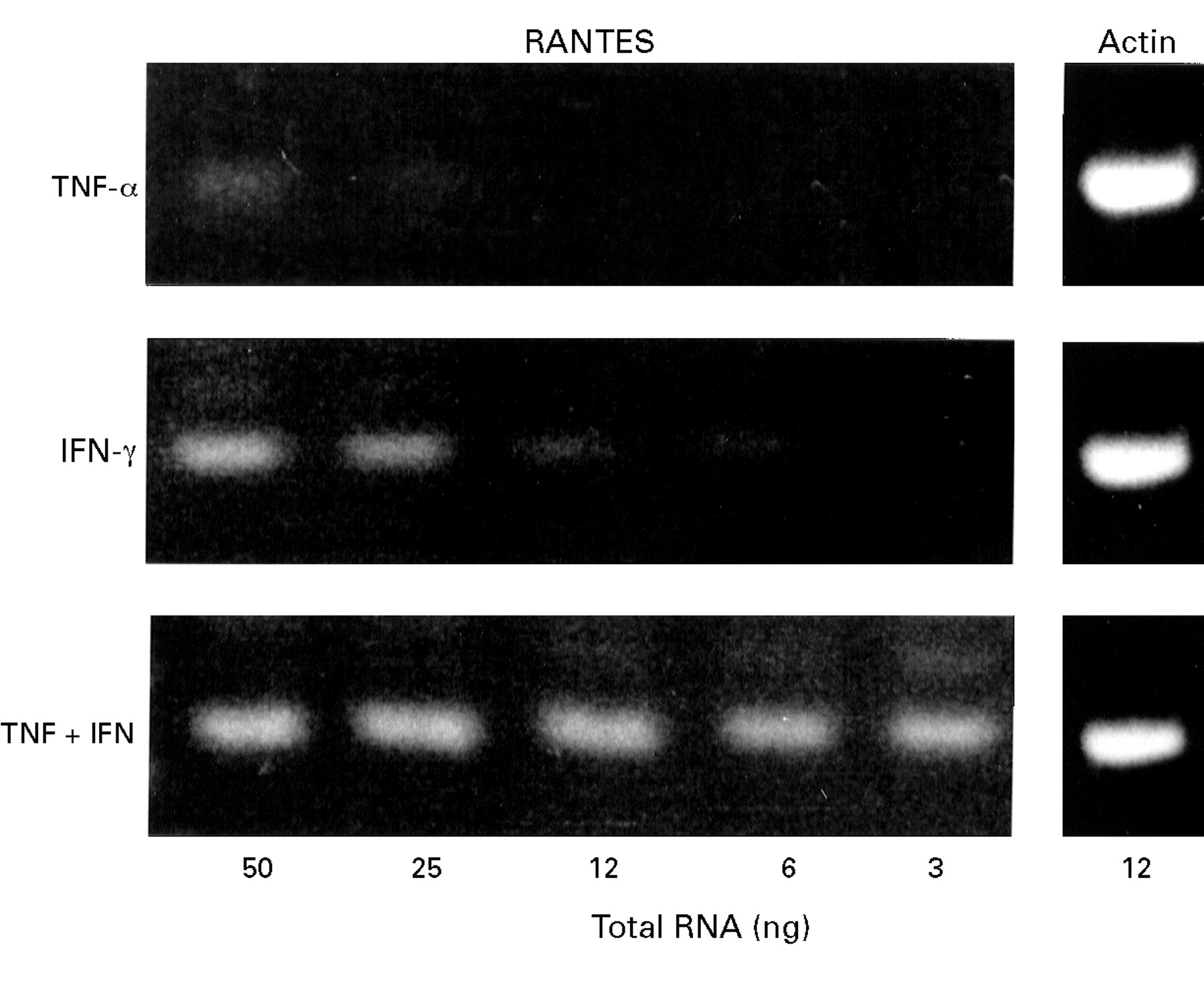
The effects of IFN-γ on chemokine secretion in these systems was investigated. Incubation of HT29-19A cells with 50 U/ml IFN-γ for 24 hours produced no significant increase in basal or TNF-α stimulated secretion of the α chemokine IL-8 or basal secretion of the β chemokines RANTES and MCP-1 (fig 3). However, in cells treated with 50 ng/ml TNF-α, IFN-γ stimulated a dramatic upregulation of both RANTES and MCP-1 secretion. TNF-α stimulated MCP-1 secretion by 20-fold in the presence of IFN-γ while RANTES secretion, which was virtually undetectable in cells exposed to TNF-α, increased by 80-fold when IFN-γ was also present (fig 3). RT-PCR of serial dilutions of HT29-19A RNA showed that the upregulation of RANTES secretion by combinations of IFN-γ and TNF-α was associated with a synergistic increase in RANTES mRNA expression compared with TNF-α or IFN-γ alone (fig 4). Up regulation of RANTES secretion was dependent on the concentrations of both TNF-α and IFN-γ (fig 5). In the presence of IFN-γ (10 U/ml), RANTES secretion was stimulated over a dose range of TNF-α with an approximated EC50 of 20–25 ng/ml TNF-α. The EC50 for IFN-γ stimulation of RANTES secretion in the presence of 50 ng/ml TNF-α was 50 U/ml (fig 5inset). Synergism between IFN-γ and TNF-α was also observed in HT29-19A cells grown as polarised monolayers on filter supports (fig 6).

Influence of IFN-γ on basal and TNF-α stimulated secretion of α and β chemokines in HT29-19A cells. Chemokines were measured 24 hours after incubation with control media or 50 ng/ml TNF-α in the presence or absence of 50 IU/ml IFN-γ. Results are mean (SEM) of four observations in each group.

Synergistic effect of TNF-α and IFN-γ on RANTES mRNA expression in the HT29-19A cell line. Cells were incubated with TNF-α alone (50 ng/ml), IFN-γ alone (50 IU/ml), or TNF-α + IFN-γ for 24 hours. The expression of RANTES specific product (243 bp) is shown after RT-PCR analysis of varying amounts (3–50 ng) of total RNA isolated from each treatment group. RT-PCR for actin using a single concentration (12 ng) confirmed the integrity of the RNA in each group.

Dose dependence of RANTES secretion in response to combinations of TNF-α and IFN-γ in the HT29-19A cell line. Cells were incubated with varying concentrations of TNF-α, either alone or in the presence of 10 or 50 U/ml IFN-γ. Results are mean (SEM) of four experiments. (Inset: IFN-γ dose response for RANTES secretion in the presence of 50 ng/ml TNF-α showing mean (SEM) for 6–8 observations in each group.)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
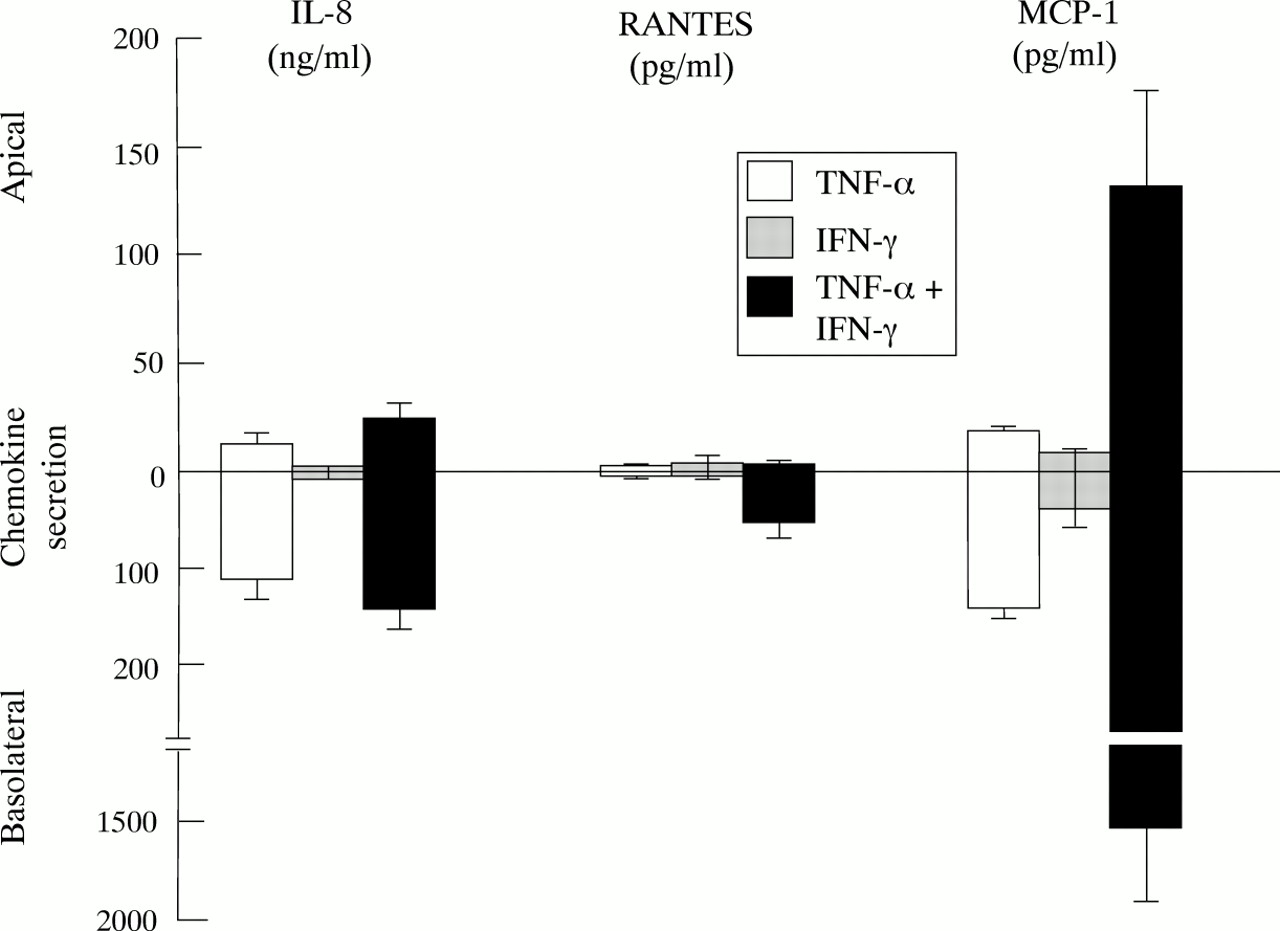
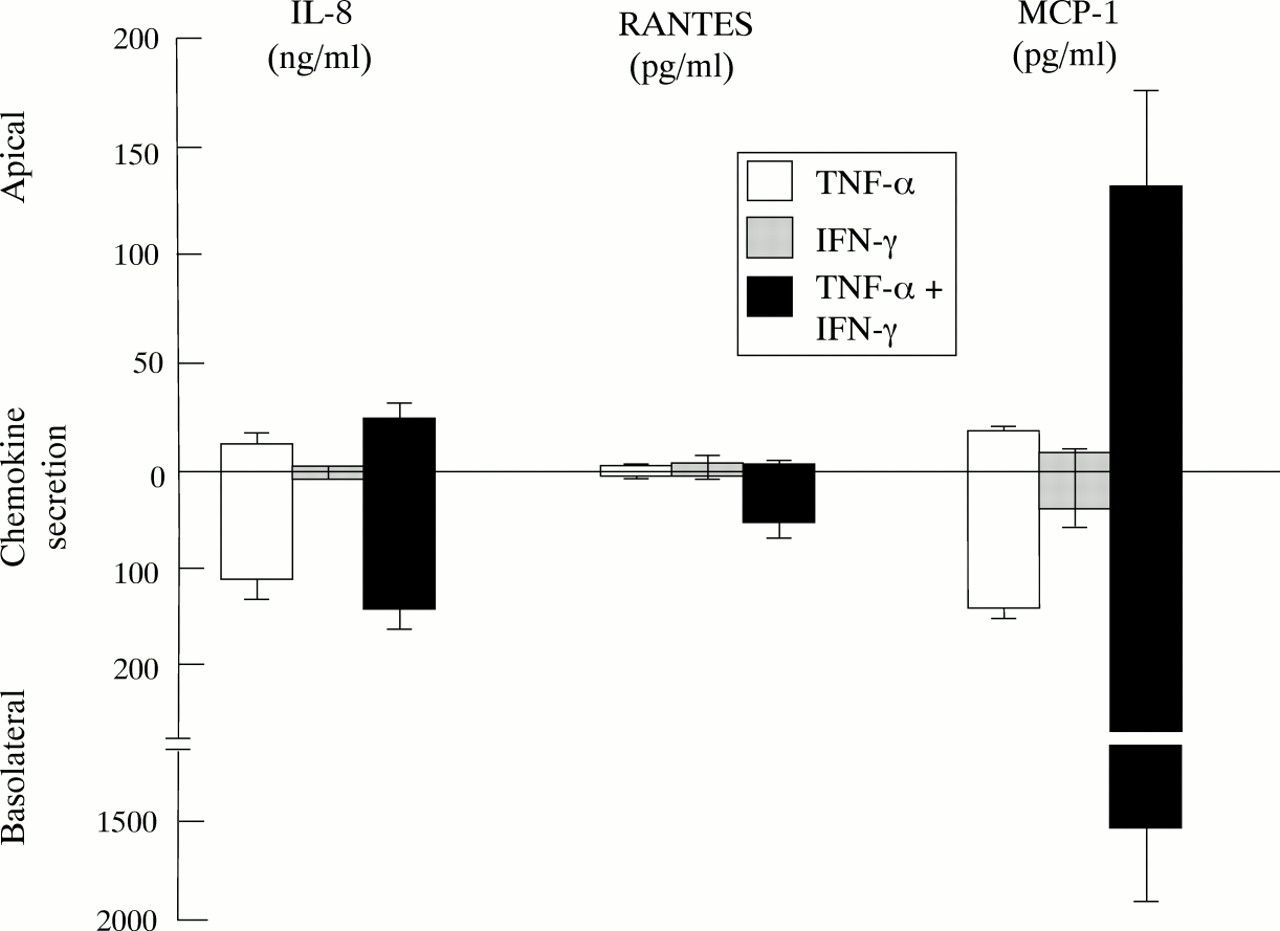
Effect of TNF-α and IFN-γ on vectorial secretion of chemokines from polarised HT29-19A monolayers. Cells were grown to confluency on Transwell filter supports and the integrity of the resulting monolayer checked by monitoring development of transepithelial resistance. TNF-α (50 ng/ml), IFN-γ (50 IU/ml), or TNF-α + IFN-γ was added to the basolateral reservoir. The concentration of IL-8, MCP-1, and RANTES in the medium was measured in both apical and basolateral reservoirs after 24 hours. Values are mean of four monolayers in each group except IFN-γ alone on MCP-1 secretion where n=2.
VECTORIAL SECRETION OF CHEMOKINES
Recent evidence has shown that combinations of TNF-α and IFN-γ modulate tight junctions in colonic cell lines, causing a decrease in the electrical resistance of cell monolayers.17 To investigate whether there is a correlation between this phenomenon and the synergistic stimulation of β chemokine secretion we examined the effects of TNF-α and IFN-γ on transepithelial electrical resistance (Rt) in HT29-19A monolayers. Basolateral incubation with IFN-γ (50 U/ml) or TNF-α (50 ng/ml) alone had no significant effect on HT29-19A Rt over a 24 hour incubation period (control: 214 (68); with IFN-γ: 270 (45); with TNF-α: 260 (96) ohms/cm2, n=4). When IFN-γ and TNF-α were added together, monolayer Rt significantly increased over 24 hours (IFN-γ + TNF-α: 393 (58) ohms/cm2, n=4, p<0.05). However, continued exposure to this combination of IFN-γ and TNF-α elicited a marked decrease in Rt over the following 48 hours (control: 285 (95); IFN-γ + TNF-α: 39 (11) ohms/cm2 at 72 hours, n=4, p<0.01). Significant stimulation of RANTES and MCP-1 secretion occurred after incubation for 8–12 hours with IFN-γ and TNF-α and was maximal at 24 hours (data not shown). This indicates that the synergistic stimulation of β chemokine expression in response to TNF-α and IFN-γ is occurring independently of the loss of tight junctional integrity, which is mediated over a longer time course. In marked contrast to HT29-19A, Caco-2 cells showed no change in secretion of either α or β chemokines in response to IFN-γ alone or in combination with TNF-α (data not shown). Similarly, IFN-γ had a relatively minor effect on the Rt of Caco-2 monolayers in the presence of TNF-α, probably indicating that this cell line expresses a much lower level of functional IFN-γ receptors.
Discussion
Increased mucosal infiltration by leucocytes is a characteristic feature of inflammatory bowel disease and chemotactic factors play a key role in the recruitment of inflammatory cells. Intestinal epithelial cells act as the interface between the contents of the gut lumen and the internal milieu, which places them in an ideal position to detect and respond to proinflammatory stimuli.
In this study we have shown that human colonic cell lines, HT29-19A and Caco-2, express distinct patterns of chemokine secretion, both constitutively and after stimulation by inflammatory agonists. HT29-19A cells, which have a phenotype similar to that expected of Cl− secretory cells in the crypt epithelium,18 secreted high levels of both IL-8 and GROα, primarily in response to TNF-α and IL-1. These chemokines were secreted in parallel, probably via a common pathway as judged by their virtually identical time course for secretion and dose responsiveness to cytokine stimulation, suggesting that they may have complementary roles in the inflammatory process. In this context, although both IL-8 and GROα bind to the α chemokine receptor IL-8RB, only IL-8 binds to the IL-8RA receptor.19
In contrast, HT29-19A cells show little or no secretion of the β chemokines RANTES, MCP-1, and MIP-1α in response to the same agonists. Caco-2 cells, which have features more typical of the villus absorptive cell (brush border differentiation, expression of Na+ dependent coupled transport systems), exhibited a quite different pattern of chemokine secretion. These cells were poor secretors of α chemokines, with only IL-1β stimulating a modest secretion of IL-8 and GROα, but showed significant expression of the β chemokine MCP-1, which acts preferentially in the recruitment and stimulation of monocytes.20 At present it is unclear whether the different profiles of chemokine secretion by HT29-19A and Caco-2 cells reflect their different phenotypes or levels of differentiation. However, preliminary observations showing that butyrate, which has been shown to induce epithelial differentiation in some epithelial cell lines, can modulate the pattern of α and β chemokine expression in Caco-2 cells,21 lend support to the hypothesis that the profile of chemokine secretion could vary within different enterocyte populations.
One of the most interesting aspects of this study was the finding that IFN-γ stimulates a differential upregulation of α and β chemokine secretion in HT29-19A cells. Interferon γ is produced primarily by Th1 lymphocytes and has been shown to be elevated in patients with both active ulcerative colitis and active Crohn’s disease.22 ,23 Interferon γ is also produced by natural killer cells but these are reported to be present only in small numbers in normal bowel mucosa24 and are further reduced in IBD.25 Interferon γ has several effects on intestinal epithelial cells, including induction of major histocompatibility complex (MHC) class II molecule expression26 and stimulation of epithelial-neutrophil adhesion,27 leading to the suggestion that IFN-γ may transform the epithelial cell surface from its classic transport function to an immune accessory function. In the present study, IFN-γ was shown to be capable of transforming the chemokine profile of HT29-19A cells during stimulation with TNF-α, producing a marked upregulation of β chemokines, MCP-1, and RANTES while having relatively little effect on IL-8 and GROα secretion. The effect on RANTES expression was particularly interesting, since RANTES protein was virtually undetectable in HT29-19A cells stimulated by IFN-γ, TNF-α, or IL-1β alone. However, combinations of IFN-γ and TNF-α induced a synergistic production of RANTES at the level of both mRNA and secreted protein. Synergy between IFN-γ and TNF-α has also been reported for RANTES expression in fibroblasts,28 endothelial cells,10 and bronchial epithelial cells,6suggesting that the combination of these agonists may be an important determinant for RANTES secretion.
The mechanism of upregulation by IFN-γ remains to be defined. Rodriguez et al,17 in a study of the effects of IFN-γ/TNF-α combinations on epithelial barrier function, suggest that IFN-γ acts by inducing expression of TNF-α receptors in HT29-19A cells. However, this would appear an unlikely explanation for the induction of β chemokine secretion in these cells since the potent activation of IL-8 secretion by TNF-α alone suggests that TNF-α receptors are already expressed at high levels in the absence of IFN-γ. Interaction at postreceptor sites such as the activator protein 1 and nuclear factor κB29 may account for the synergistic effects of IFN-γ and TNF-α observed here, although further studies are needed to confirm this. The inability of IFN-γ to up regulate chemokine production in Caco-2 cells may indicate that different control mechanisms regulate transcription of RANTES in different cell types30 or more likely, given the relative lack of effect of IFN-γ on Caco-2 tight junction integrity of Caco-2 monolayers, that IFN-γ receptors are expressed at a much lower level by this cell type.
The production of β chemokines by the intestinal epithelium in response to simultaneous activation of IFN-γ producing Th1 cells and TNF-α producing monocytes could contribute to the development of chronic inflammatory infiltrates. It is interesting to speculate that failure to control β chemokine secretion as a result of inappropriate activation of specific T cell immunity underlies the pathology of inflammatory bowel disease. This is supported by the observation that T lymphocytes with a Th1 profile predominate in IBD.22 ,23However, considerable caution is required in extrapolating the responses of tumour derived cell lines to those of epithelial cells in vivo and confirmation of the ability of the gut epithelium to secrete this class of chemokines will be an important aim of future studies.
The present study provides evidence that epithelial cells derived from the intestine are a potential source for the IFN-γ induced secretion of β chemokines, although further studies will be necessary to establish their significance in the promotion of chronic inflammatory infiltrates in IBD.
Acknowledgments
We thank the North West Regional Health Authority for financial support.