Article Text

Abstract
Background—The cytokines interleukin 1β (IL-1β) and tumour necrosis factor α (TNF-α) are inhibitors of gastric acid secretion when administered systemically.
Aims—To investigate the inhibitory effect of IL-1β and TNF-α on cultured, acid secreting parietal cells in order to determine the mechanism of this inhibition.
Methods—Rabbit parietal cells were prepared by collagenase-EDTA digestion and counter flow elutriation. Acid secretory activity was assessed by aminopyrine accumulation.
Results—IL-1β and TNF-α inhibited basal and stimulated acid secretion in a dose dependent manner; near maximal effects were seen with both at 10 ng/ml. Inhibition was maximal with 15 minutes pretreatment but seen with up to 18 hours of preincubation. Both cytokines inhibited histamine, carbachol, gastrin, forskolin, and A23187 stimulated acid secretion but had no effect on stimulation by dibutyryl-cAMP. Inhibition of acid secretion was not accompanied by a change in radioligand binding to histamine H2 or gastrin/CCKB receptors. Pertussis toxin abolished the inhibitory effects on histamine and forskolin stimulation. The tyrosine kinase inhibitor herbimycin reduced the inhibitory effects of TNF-α against all stimuli but only reduced the effects of IL-1β against histamine and forskolin stimulation.
Conclusions—IL-1β and TNF-α seem to inhibit parietal cell acid secretion by multiple pathways; the inhibition occurs at postreceptor level and involves pertussis toxin and tyrosine kinase dependent and independent pathways. Mucosal production of cytokines may be important in the regulation of gastric acid secretion.
- acid secretion
- aminopyrine
- cytokines
- interleukin 1β
- tumour necrosis factor α
- parietal cell
Statistics from Altmetric.com
Interleukin 1β (IL-1β) and tumour necrosis factor α (TNF-α) are multifunctional cytokines.1 In addition to important roles in regulating the host’s inflammatory response and playing a part in the tissue injury seen in many inflammatory states,2 they have been shown to mediate a wide variety of biological effects in other cell types.3 ,4 Peripheral and central administration of IL-1 to experimental animals potently reduces gastric acid secretion5 ,6 and parenteral TNF-α7 has also been shown to inhibit acid secretion. Similarly, IL-1 has been reported to reduce the gastric injury produced by a wide variety of noxious stimuli, including ethanol, non-steroidal anti-inflammatory agents, cysteamine, and water immersion stress.8-10 Inhibition of acid secretion contributes to this gastric protection.6 In addition both cytokines may play a part in healing of tissue injury as TNF-α can promote angiogenesis and vascular remodelling11 and IL-1 can promote cartilage and bone remodelling.3 Therefore, locally released cytokines might play an important part in the healing of gastric mucosal lesions and ulcers. The mechanism and site of action for the acid suppressive effects of IL-1 and TNF-α have not been fully defined. The acid secreting parietal cell is the final common pathway of acid secretion and it is likely that locally produced cytokines may inhibit acid secretion by acting directly on the parietal cell. In addition to the regulatory peptides and mediators, such as gastrin, histamine, muscarinic agonists, prostaglandins, and somatostatin, which have been known for some time to regulate parietal cell function,12 recent studies have also suggested that growth factors such as epidermal growth factor (EGF) and transforming growth factor α (TGF-α)13 ,14 and inflammatory mediators such as platelet activating factor15 are also capable of directly regulating acid secretion by the parietal cell. By contrast with the many in vivo studies showing inhibition of acid secretion by the cytokines, the mechanism of the inhibition by IL-1β and TNF-α has received relatively little attention by in vitro studies. One study in acutely isolated canine parietal cells16 showed that TNF-α and IL-1α inhibited receptor dependent secretagogue stimulated aminopyrine accumulation but did not provide much information on the underlying mechanisms for this action. Therefore, we have studied the effect of IL-1β and TNF-α on cultured rabbit parietal cells, using receptor dependent and independent stimuli to define the site(s) of action and pertussis toxin and the tyrosine kinase inhibitor herbimycin A as tools to determine the mechanism of action. This should provide further insights into the molecular events involved in the regulation of acid secretion and the control of gastric secretion in disease states.
Materials and methods
CHEMICALS
Histamine, carbachol (carbamylcholine chloride), recombinant human TNF-α, recombinant human IL-1β, gastrin-17, Earle’s balanced salt solution (EBSS), A23187, bovine serum albumin (BSA), pertussis toxin, collagenase type I, collagenase type H, dithiothreitol, penicillin G, crystalline bovine insulin, hydrocortisone, streptomycin, gentamicin, dinitrophenol, and EDTA were purchased from Sigma, Poole, UK. Forskolin, herbimycin A, N6,O2-dibutyryladenosine 3, 5-cyclic monophosphate (dibutyryl cyclic AMP: dbcAMP) were obtained from LC laboratories, Nottingham, UK; Triton X-100 and N-2-hydroxyethylpiperazine-N′-ethansulphonic acid (HEPES) were from BDH-Merck (Poole, UK); 14C-dimethylamine-aminopyrine (103 mCi/mmol) was obtained from Amersham International, Amersham, UK; and 125I-CCK-8 (2200 Ci/mmol) and 3H-histamine (50 Ci/mmol) from Dupont-NEN, Stevenage, UK. Ham’s F12/Dulbecco’s modified Eagle’s culture media (F12/DMEM) (50:50 vol/vol), glutamine, Hank’s balanced salt solution (HBSS), basal medium Eagle’s (BME), and fetal calf serum were obtained from Gibco, Paisley, UK; basement membrane Matrigel was from Universal Biologicals, London, UK; and ranitidine was from Glaxo, Uxbridge, UK.
PARIETAL CELL PREPARATION
Rabbit parietal cells were isolated and enriched by a modification of previously described methods.17-19 New Zealand white rabbits (body weight 2–2.5 kg) were killed by overdose of sodium pentobarbitone and the stomach immediately excised. Gastric fundic mucosa was digested with sequential exposure to collagenase (type I 0.175 g/l with type H 0.175 g/l) and EDTA. Parietal cells were enriched from the crude suspension with a Beckman JE 5.0 elutriator rotor using the standard elutriation chamber.17 For selected experiments further purification of parietal cells was performed using density gradient centrifugation with 50% percoll as previously described.20
CELL CULTURE
Cells from the parietal cell enriched fractions were collected by brief centrifugation and pooled and resuspended in complete culture medium (Ham’s F12/DMEM 50/50 nutrient mix, containing 10% heat inactivated fetal calf serum, 10 mM HEPES pH 7.4, 100 mg/l gentamicin, 100 mg/l streptomycin, 100 mg/l penicillin, 2 mM glutamine, 8 μg/ml hydrocortisone, and 1 μg/ml insulin) and then cultured with a modification of previously described methods.19 Parietal cells were then plated on to Matrigel coated 12 and 24 well tissue culture plates (Corning) at 0.5–1.0 million cells/well. The coated plates had been prepared by diluting Matrigel 1:7 with sterile water and then uniformly coating the wells. After setting with overnight incubation at 37°C, the remaining water was aspirated and the wells were allowed to dry in a culture hood for 60 minutes. The plates were then equilibrated with 1 ml culture medium before plating the cells. The parietal cell enriched fraction was then cultured at 37°C in an atmosphere of 5% CO2/95% air for 40 hours.
MEASUREMENT OF ACID SECRETION
Intracellular accumulation of 14C-aminopyrine was used as a measure of functional acid secretory activity.18-21Cultured cells in 24 well plates were washed once with 2 ml EBSS containing 0.1 % BSA, 10 mM HEPES pH 7.4, 2 mM glutamine, and 0.22% NaHCO3 to remove dead and non-adherent cells and then 1 ml of the above medium was added and 0.1 μCi aminopyrine and the various stimulant substances were added simultaneously to each well. Cells were then incubated for 30 minutes at 37°C in an atmosphere of 5% CO2/95% air. Incubations were terminated by removing the medium from each well using a vacuum pump and then washing twice with 1 ml EBSS solution. Cells were then lysed with 1 ml 1% Triton X-100. Aliquots of cell lysates and incubation media were counted in Optiphase Safe, Wallac, Milton Keynes, UK, using a Beckman LS 1801 liquid scintillation counter with DPM correction. Dinitrophenol (0.1 mM) was added to separate wells to assess non-specific incorporation and values were subtracted from test values.13 For most of the studies with recombinant cytokines, the TNF-α and IL-1β were added to the wells 15 minutes before the initial washing step and before adding the aminopyrine and stimulants. Pilot studies showed that this method gave results corresponding to those obtained if the cells were washed first and then cytokines added and preincubated for 15 minutes before being stimulated. For time course experiments the cytokines were added to the culture medium at intervals up to 18 hours before aminopyrine accumulation was measured simultaneously in all test wells. When appropriate, pertussis toxin (200 ng/ml) was added two hours before and herbimycin was added one hour before the cytokines.
RECEPTOR BINDING STUDIES
Binding of 3H-histamine to H2 receptors and 125I-CCK-8 to gastrin/CCKB receptors on parietal cells was assessed as previously described (labelled CCK-8 has been shown previously to bind with equal efficacy as125I-[Leu15]gastrin to parietal cell CCKB receptors and was used as tracer because of its ready availability).22 ,23 Briefly, cells on Matrigel coated 12 well plates were washed three times with HBSS containing 0.1% BSA to remove non-adherent cells and incubated at 37°C with 10 fmol125I-Bolton-Hunter CCK-8 for 60 minutes or 1 pmol3H-histamine for 30 minutes in the presence or absence of unlabelled gastrin or the H2-receptor antagonist ranitidine as appropriate. Binding reactions were terminated by aspirating the medium containing the unbound label and then the cells were washed twice with ice chilled HBSS. The adherent cells were solubilised by incubating in 1% Triton X-100 and the bound and unbound radioactivity was counted in a gamma counter (Wallac 1260, Wallac, Milton Keynes, UK) or scintillation counter as necessary. Binding of 125I-CCK in the presence of 10-6 M gastrin and3H-histamine binding in the presence of 10-4 M ranitidine was regarded as non-specific binding and these values were subtracted from the total binding to obtain the values for specific binding.
STATISTICAL ANALYSIS
During each set of experiments, each condition was assessed in duplicate wells and compared with basal (unstimulated) and the appropriate stimulated aminopyrine accumulation from wells on the same 24 well plate. The mean of the data from one animal preparation was regarded as n=1. For time course experiments the effects of cytokines were compared with parietal cells on the same 24 well plate treated identically but without addition of cytokine. Data are presented as the mean (SEM) of 3–12 different animal preparations. Graphical data are presented as aminopyrine accumulation compared with basal values to enable comparison of the relative stimulation seen with the different agonists. Student’s paired t test was used to compare the effect of cytokines with the appropriate controls. A p value <0.05 was regarded as significant.
Results
EFFECT OF IL-1β AND TNF-α ON UNSTIMULATED AMINOPYRINE ACCUMULATION
Both cytokines produced a reduction in basal acid secretion, when cells were exposed to cytokines 15 minutes before uptake was measured. This was evident over the dose ranges tested (0.1–100 ng/ml). Basal activity was an aminopyrine ratio of 441 (39) and this was reduced by TNF-α by 15(4)% (p<0.05; fig 1) and IL-1β by 25 (3)% (p<0.05; fig 2) (both at 100 ng/ml).

Effect of increasing concentrations of TNF-α on aminopyrine (AP) uptake in cultured rabbit parietal cells. Cells were incubated with cytokine for 15 minutes and then aminopyrine accumulation was measured over 30 minutes. Uptake of aminopyrine was measured in the basal state and when stimulated by histamine (10−4 M) and carbachol (10−4 M). Results are expressed as percentage (mean (SEM)) of basal control (n=6). *p<0.05; **p<0.01.

Effect of increasing concentrations of IL-1β on aminopyrine (AP) uptake in cultured rabbit parietal cells. Cells were incubated with cytokine for 15 minutes and then aminopyrine accumulation was measured over 30 minutes. Uptake of aminopyrine was measured in the basal state and when stimulated by histamine (10−4 M) and carbachol (10−4 M). Results are expressed as percentage (mean (SEM)) of basal control (n=6). *p<0.05; **p<0.01.
EFFECT OF IL-1β AND TNF-α ON HISTAMINE AND CARBACHOL STIMULATED AMINOPYRINE ACCUMULATION
Both IL-1β and TNF-α reduced secretagogue stimulated acid secretion in a dose dependent manner. Over the range tested (0.1–100 ng/ml) both cytokines inhibited histamine and carbachol stimulated acid secretion. TNF-αproduced significant reductions in histamine and carbachol stimulated aminopyrine uptake at 10 and 100 ng/ml (maximal reduction 23 (2)% for histamine (p<0.05) and 63 (12)% for carbachol (p<0.01; fig 1)) and IL-1β significantly impaired the actions of both secretagogues at doses from 1–100 ng/ml (maximal inhibition 23 (4)% for histamine (p<0.01 ) and 56 (7)% for carbachol (p<0.01; fig 2). The degree of inhibition by both cytokines was greater for carbachol stimulated acid secretion. Because of the reliable near maximal inhibition produced by both cytokines at 10 ng/ml, this concentration was used for further characterisation.
EFFECT OF TIME OF PRETREATMENT ON CYTOKINE MEDIATED INHIBITION OF ACID SECRETION
Pretreatment of the cells with cytokines (10 ng/ml) for 15 minutes seemed to give maximal degrees of inhibition although significant and similar degrees of inhibition of basal, histamine, and carbachol stimulated acid secretion were still evident when pretreatment was prolonged up to 18 hours before stimulation. Therefore, the 15 minute time point was used for the studies on the mechanisms of inhibition.
EFFECT OF IL-1β AND TNF-α ON GASTRIN MEDIATED ACID SECRETION
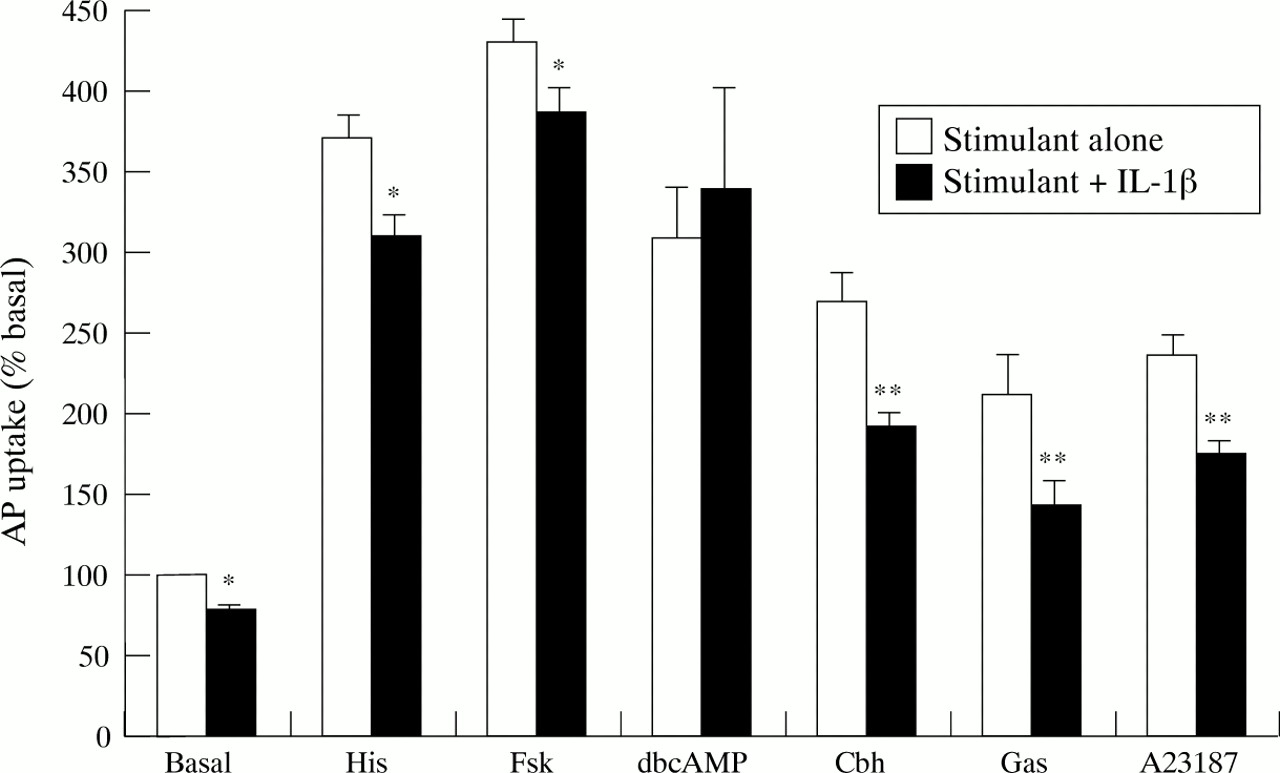
The effect of the cytokines on gastrin mediated secretion was assessed to determine whether the inhibitory effects were seen against other secretagogues. Parietal cells stimulated with gastrin 10−7 M showed a modest acid secretory response (AP ratio of 929 (63), compared with 10−4 M histamine 1675 (102) and 10−4 M carbachol 1203 (89)) and this response was unaffected by 10−5 M ranitidine showing that it was due to direct stimulation of the parietal cell and not via the intermediary of a histamine secreting cell. This acid response to gastrin was significantly impaired by both 10 ng/ml TNF-α (by 57 (6) %: p<0.01; fig 3) and IL-1β (by 57 (12)%: p<0.01; fig 4). The results obtained with the parietal cells cultured after the percoll gradient gave identical degrees of inhibition with both cytokines against histamine, carbachol, and gastrin stimulation to that seen with the cells from the elutriator only enriched fraction (data not shown). Therefore the elutriated but not percoll purified cells were used for further study.

Effect of TNF-α (10 ng/ml) on aminopyrine accumulation by parietal cells in the basal state and when stimulated by histamine (His; 10−4 M), forskolin (Fsk; 10−5 M), dibutyryl-cAMP (dbcAMP; 10−4 M), carbachol (Cbh; 10−4 M), gastrin-17 (Gas; 10−7 M), or A23187 (10−6 M). Cells were exposed to TNF-α for 15 minutes before stimulation. Results are expressed as percentage of basal aminopyrine uptake in the absence of cytokine (mean (SEM)), n=5–6. *p<0.05; **p<0.01.

Effect of IL-1β (10 ng/ml) on aminopyrine (AP) accumulation by parietal cell in the basal state or stimulated by histamine (His; 10−4 M), forskolin (Fsk; 10−5 M), dibutyryl-cAMP (dbcAMP; 10−4 M), carbachol (Cbh; 10−4 M), gastrin-17 (Gas; 10−7 M), or A23187 (10−6 M). Cells were exposed to IL-1β for 15 minutes before stimulation. Results are expressed as percentage of basal aminopyrine uptake in the absence of cytokine (mean (SEM)), n=5–6. *p<0.05; **p<0.01.
EFFECT OF IL-1β AND TNF-α ON SECRETAGOGUE BINDING TO PARIETAL CELLS
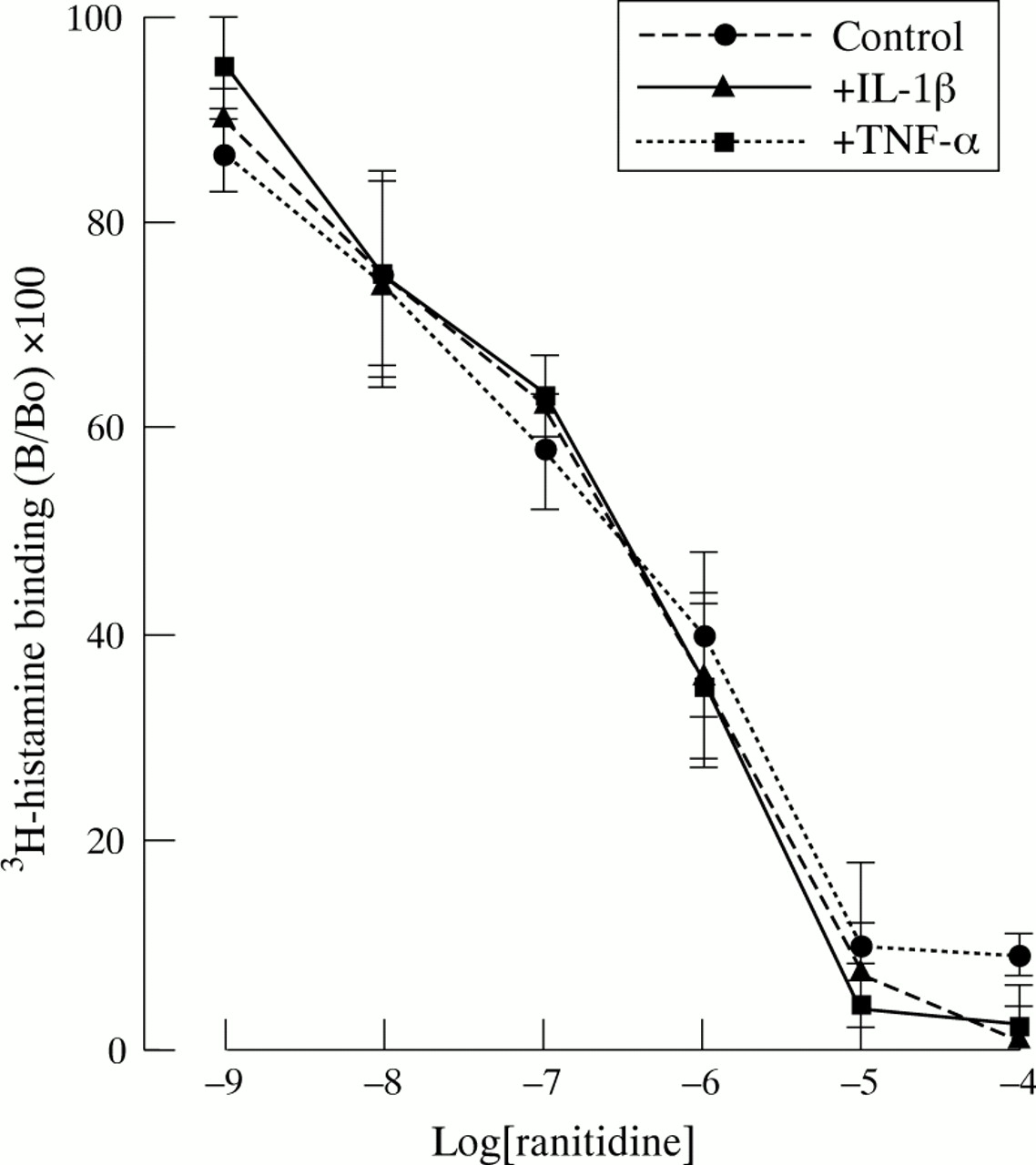
To test whether the inhibitory effect of TNF-α and IL-1β on secretagogue stimulated aminopyrine uptake was mediated in part through down regulation of the respective receptors, we examined the effect of these cytokines on binding of the appropriate ligands. Neither IL-1β nor TNF-α at concentrations which were effective in inhibiting the stimulatory actions of histamine (fig 5) and gastrin (fig 6) had any significant effect on the specific or non-specific binding or displacement of radioligands.

Effect of TNF-α and IL-1β (both 10 ng/ml) on binding of 3H-histamine to rabbit parietal cells. Cells were incubated in appropriate cytokine for 15 minutes before performing the binding experiments. Data are expressed as percentage of binding in the absence of unlabelled histamine (B/Bo). Results are the mean (SEM) of three separate animal preparations.

Effect of TNF-α and IL-1β (both 10 ng/ml) on binding of 125I-CCK to rabbit parietal cells. Cells were incubated in appropriate cytokine for 15 minutes before performing the binding experiments. Data are expressed as percentage of binding in the absence of unlabelled gastrin (B/Bo). Results are the mean (SEM) of three separate animal preparations.
EFFECT OF IL-1β AND TNF-α ON RECEPTOR INDEPENDENT STIMULI OF ACID SECRETION
To investigate whether the cytokines could inhibit parietal cell acid secretion at sites distal to the interactions of secretagogues with their cell surface receptors, we used forskolin (10−5 M) to activate adenylate cyclase directly, dbcAMP (10−4 M), as a direct analogue of intracellular cAMP and the calcium ionophore A23187 (10−6 M) to raise intracellular calcium concentrations directly and thus mimic activation of the muscarinic receptor. All three agents were effective in stimulating acid secretion (AP ratios: forskolin 1938 (107), dbcAMP 1384 (134), A23187 1062 (71)). Control experiments confirmed that the H2 receptor antagonist ranitidine (10−5 M) and the muscarinic antagonist atropine (10−5 M) completely abolished the stimulatory actions of histamine and carbachol, respectively. However neither antagonist had any inhibitory effects against basal aminopyrine accumulation or when cells were stimulated by any of the receptor independent stimuli (data not shown). Both cytokines significantly reduced forskolin and A23187 stimulated acid secretion, but had no inhibitory action against the stimulatory effect of dbcAMP. Interleukin 1β 10 ng/ml reduced forskolin stimulated acid secretion by 14 (2)% (p<0.05) and A23187 by 47 (3)% (p<0.01; fig 4) and TNF-α inhibited these stimulatory effects by 16 (3) (p<0.05) and 65 (6)% (p<0.01), respectively (fig3).
EFFECT OF PERTUSSIS TOXIN PRETREATMENT ON CYTOKINE INHIBITION OF ACID SECRETION
Pretreatment of cultured parietal cells with pertussis toxin (200 ng/ml for two hours) did not alter the aminopyrine response in unstimulated or histamine, carbachol, forskolin, gastrin, or A23187 stimulated cells. However, pertussis toxin pretreatment abolished the inhibitory effects of both 10 ng/ml TNF-α (fig 7) and 10 ng/ml IL-1β (fig 8) on both histamine and forskolin stimulated aminopyrine accumulation. Such pertussis toxin pretreatment had no effect on the inhibitory actions seen against carbachol, gastrin or A23187.

Effect of pertussis toxin (PTX; 200 ng/ml) on TNF-α (10 ng/ml) inhibition of aminopyrine (AP) accumulation in the basal state or stimulated by histamine (His; 10−4 M), forskolin (Fsk; 10−5 M), carbachol (Cbh; 10−4 M), gastrin-17 (Gas; 10−7 M), or A23187 (10−6M). Cells were incubated with PTX for two hours before addition of cytokine. Data are expressed as mean (SEM), n=6. *p<0.05 compared with the effect of TNF-α in the absence of PTX.
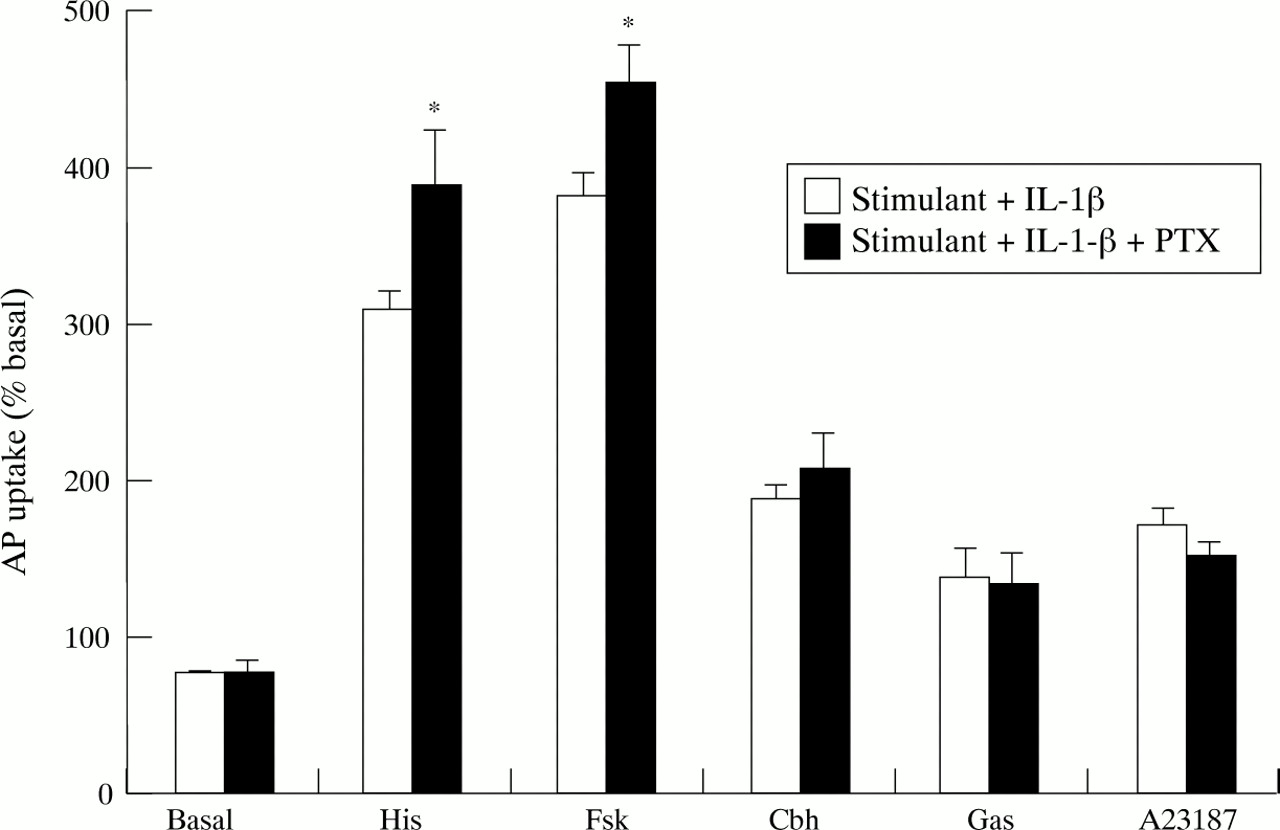
Effect of pertussis toxin (PTX; 200 ng/ml) on IL-1β (10 ng/ml) inhibition of aminopyrine (AP) accumulation in the basal state or stimulated by histamine (His; 10−4 M), forskolin (Fsk; 10−5 M), carbachol (Cbh; 10−4 M), gastrin-17 (Gas; 10−7 M), or A23187 (10−6M). Cells were incubated with PTX for two hours before addition of cytokine. Data are expressed as mean (SEM), n=6. *p<0.05 compared with the effect of IL-1β in the absence of PTX.
EFFECT OF THE TYROSINE KINASE INHIBITOR HERBIMYCIN ON THE INHIBITORY EFFECT OF CYTOKINES ON ACID SECRETION
The role of tyrosine kinases in mediating the inhibitory effects of the cytokines was assessed by pretreating the cells with herbimycin (10−6 M) for 60 minutes before adding the cytokines. Preliminary experiments confirmed that this concentration had no effects on basal or stimulated aminopyrine uptake and was not associated with toxicity and loss of viability as assessed by MTT assay.24 Higher doses tended to reduce both viability and acid secretion and were therefore not used in the studies into the mechanism of cytokine action. Herbimycin at 10−6 M was effective in abolishing the inhibitory effect of both cytokines on basal and histamine and forskolin stimulated aminopyrine uptake (figs 9and 10). Herbimycin significantly reduced the inhibitory effect of TNF-α against carbachol and A23187 (fig 9) but the inhibitory effects of IL-1β in this situation persisted even with herbimycin pretreatment (fig 10).
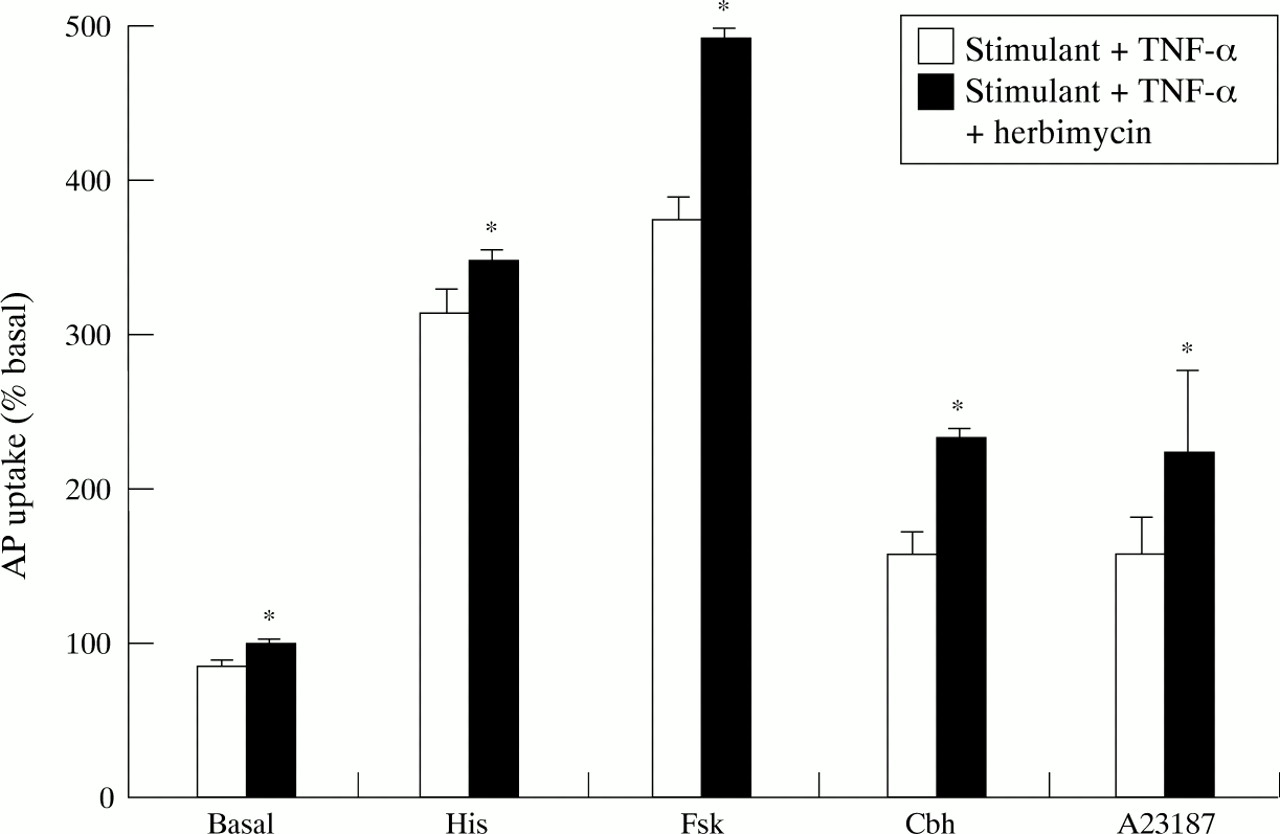
Effect of the tyrosine kinase inhibitor herbimycin (10−6 M) on TNF-α (10 ng/ml) induced inhibition of aminopyrine (AP) uptake in rabbit parietal cells in the basal state or stimulated with histamine (His; 10−4 M), forskolin (Fsk; 10−5 M), carbachol (Cbh; 10−4 M), or A23187 (10−6 M). Herbimycin was added 60 minutes before TNF-α. Data are expressed as mean (SEM), n=4. *p<0.05 compared with the effect of TNF-α in the absence of herbimycin.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Effect of herbimycin (10−6 M) on IL-1β (10 μg/ml) inhibition of aminopyrine (AP) accumulation in rabbit parietal cells in the basal state or stimulated with histamine (His; 10−4 M), forskolin (Fsk; 10−5 M), carbachol (Cbh; 10−4 M), or A23187 (10−6M). Herbimycin was added 60 minutes before IL-1β. Data are expressed as mean (SEM), n=5. *p<0.05 compared with effect of IL-1β in the absence of herbimycin.
Discussion
This study shows that both recombinant cytokines IL-1β and TNF-α were effective in inhibiting acid secretion by cultured rabbit parietal cells. It also provides some new insights into the pathways involved in this inhibition at the cellular level. Both cytokines were effective in reducing unstimulated, basal aminopyrine uptake as well as that stimulated by histamine, gastrin, or carbachol. Thus the inhibitory action was not secretagogue specific, although the degree of inhibition was always characteristically greater for the agents acting through the phospholipase C/Ca2+/inositol phosphate pathway (gastrin and carbachol) than for histamine, which predominantly activates adenylate cyclase.25 This difference in effectiveness is interesting in the light of some in vivo studies showing that systemic IL-1β is considerably more effective in inhibiting pentagastrin as opposed to histamine stimulated acid secretion.26 Previous studies with the cytokines in vitro did not show a reduction in basal secretion in acutely isolated canine parietal cells16; the difference seen in our study probably stems from the relatively greater basal activity characteristic of cultured as opposed to acutely isolated cells as well as possible differences between the species studied.19However, IL-1β has been reported to decrease basal as well as stimulated acid secretion in rats in vivo.27 To examine whether the inhibitory effects of the cytokines are due to down regulation of the stimulatory cell surface receptors, the effect of acid inhibitory concentrations of cytokines on radioligand binding were investigated. Despite being effective in reducing acid secretion at 10 ng/ml neither TNF-α nor IL-1β had any effect on the binding of H2 or gastrin receptors. As binding of 125I-CCK was unaffected, further characterisation with125I-[Leu15]gastrin was not attempted. Thus although the cytokines were effective inhibitors of aminopyrine uptake, this was not associated with alterations in receptor binding. Some inhibitory actions of TNF-α on cells have been reported to be associated with reduction of receptor density.28 Parietal cell gastrin and muscarinic receptor density is known to be regulated by protein kinase C activity29 and in some systems TNF-α can lead to protein kinase C activation.30 However, the data show that under these experimental conditions the inhibitory effects of IL-1β and TNF-α on histamine or gastrin stimulation of parietal cells does not involve such changes in receptors. These data are consistent with those experiments showing that both cytokines were effective at inhibiting acid secretion stimulated by receptor independent agonists. Both IL-1β and TNF-α were effective in inhibiting the stimulatory effects of forskolin and A23187. Interestingly, the degree of inhibition seen with these approximated to the scale of effect seen with the cytokines against the respective receptor ligands—that is, the inhibition was greater for A23187 (about 45–60%) than forskolin (20%), which compares well with the effects on carbachol and gastrin (45–60%) and histamine (16–23%). Therefore it seems that both TNF-α and IL-1β inhibit parietal cell acid secretion at points distal to receptor activation. Despite being effective in inhibiting forskolin stimulated aminopyrine uptake neither cytokine was effective in reducing the stimulation mediated by dbcAMP. This suggests that the inhibition was occurring at the level of cAMP generation, probably at the level of adenylate cyclase. This pattern of inhibition is similar to that reported with TGF-α and EGF which have been shown to inhibit parietal cell acid secretion stimulated mine and forskolin, but not by dbcAMP.13 This pattern of inhibition is different from that produced by somatostatin, which also inhibits parietal cell function via multiple pathways but it seems that somatostatin acts to inhibit both cAMP generation and the actions of cAMP.31 It is also interesting to compare the degrees of inhibition produced by IL-1β and TNF-α with somatostatin, which is thought to be the main endogenous inhibitor of acid secretion, and TGF-α and EGF.
Both somatostatin31 and TGF-α/EGF inhibited agonist stimulated acid secretion by about 30–40%13 ,32; this is rather less than the inhibitory effects of IL-1β and TNF-α against carbachol stimulated acid secretion but slightly greater than the effects against histamine. Our preliminary findings have shown that submaximal inhibitory combinations of somatostatin and IL-β have roughly additive effects on acid secretion but that the maximal overall effect is unaltered. Characteristically, the maximal in vitro acid inhibitory effects of all of these bioactive peptides are significantly less than the maximal effect of potent receptor antagonists such as ranitidine or atropine18 ,33 ; however, the endogenous peptides typically inhibit acid secretion stimulated by more than one agonist. The inhibitory effects of several agents on parietal cell acid secretion are mediated by pertussis sensitive guanine nucleotide binding (G) proteins.13 ,31 In this study pertussis toxin abolished the inhibitory effects of both IL-1β and TNF-α on histamine and forskolin stimulated acid secretion. However, it had no effect on the inhibition of carbachol or A23187 stimulation. This suggests that the cytokines are inhibiting acid secretion via at least two separate pathways. Inhibition of adenylate cyclase by the inhibitory G protein Gi is well characterised34 ,35 and the removal of the inhibitory effects against histamine and forskolin strongly suggest that IL-1β and TNF-α are acting via Gi to inhibit adenylate cyclase. By contrast, pertussis toxin had no effect on the stimulation produced by receptor dependent or independent activation of the phospholipase/Ca2+ pathway, showing that this action was independent of pertussis sensitive G proteins. In some systems the activation of tyrosine kinases seems to be an important step in mediating the effects of TNF-α and IL-1β.36 ,37 In the presence of herbimycin, a difference in the effects of TNF-α and IL-1β was evident. Herbimycin reduced the inhibitory effects of TNF-α against all stimulants but had no effect on IL-1β inhibition of carbachol and A23187 stimulated acid secretion. Thus the inhibitory effects of both cytokines mediated via the pertussis toxin sensitive pathway also seem to depend on tyrosine kinase activity but there is a difference in the herbimycin sensitivity of the inhibitory actions of the two cytokines against carbachol stimulated acid secretion. The inhibitory effect of IL-1β against stimulation by carbachol and A23187 seems to be insensitive to herbimycin. As the effect of EGF on parietal cells is biphasic,14 acute exposure causing inhibition of acid secretion and more prolonged treatment actually enhancing acid secretion, we tested the effect of the cytokines for any such biphasic responses. Despite exposure of the cells for up to 18 hours before measuring acid secretion only inhibitory actions of TNF-α and IL-1β were detected. Although TNF-α and particularly IL-1β have been shown to be effective inhibitors of acid secretion in vivo exact site(s) of action have been disputed. The results of our study are in agreement with that of Nompleggi et al, in that there is a direct inhibitory action on the parietal cells and this action is not secretagogue specific.16 This does not of course rule out any additional modes and sites of action—for example, IL-1β is an extremely effective acid suppressant when given directly intracisternally38 and it also inhibits release of histamine from gastric enterochromaffin-like cells.39These multiple actions may all contribute to IL-1β being such a profound inhibitor of acid secretion. IL-1β and TNF-α are produced in inflamed gastric mucosa,40 ,41 the consequences of this may vary but local production of these cytokines would be expected from the current experimental data to reduce acid secretion. Both cytokines exhibit increased expression during healing of gastric ulceration.42 In addition to important roles in inflammation, angiogenisis, and mucosal healing, this local production of TNF-α and IL-1β may be important in paracrine inhibition of gastric acid secretion and so enhance ulcer healing. Such an effect could also contribute to the mucosal protection seen in various experimental models of gastric damage. Helicobacter pyloriinfection is the major acquired cause of duodenal and gastric ulceration43 ,44 and it is possible that the cytokines acting in a local paracrine manner to inhibit acid secretion are involved in the pathogenesis. For example, acute H pyloriinfection is associated with achlorhydria45-47 and although the pathology is variable, some patients exhibit reduced acid secretion without major parietal cell damage48 ,49 and locally produced mediators such as the cytokines are likely to be responsible. In chronic H pylori infection, increased acid secretion is characteristic of duodenal ulcer disease50but other patients may show reduced acid secretion.44 In patients with duodenal ulcer bacterial colonisation and inflammation tend to be greater in the gastric antrum51 with relative sparing of the oxyntic mucosa and it may be that increased bacterial load in the vicinity of parietal cells liberates the acid inhibitory cytokines and gives relative protection against duodenal ulcer to those non-ulcer patients with inflammation in the gastric body.
In summary, we have shown that TNF-α and IL-1β directly inhibit acid secretion by parietal cells. Multiple intracellular pathways are involved and production of these cytokines in inflammed mucosa may have an important role in the regulation of acid secretion.
Acknowledgments
Some of this work was presented in abstract form at the meetings of the British Society of Gastroenterology (Spring 1996, Brighton) and Digestive Diseases Week (San Francisco 1996). ILPB was funded by a Medical Research Council research training fellowship.