Article Text

Abstract
Background—Although inflammation in Crohn’s disease is believed to be mediated by activated T cells, genotyping of all MHC class II alleles in white people with this disease has not been reported.
Aims—To perform a detailed molecular analysis of HLA DPB, DQB, and DRB genes in white patients with Crohn’s disease and controls in order to determine if the inheritance of any class II genes confers susceptibility or resistance to this disease.
Methods—Complete molecular typing of HLA class II DPB, DQB, and DRB alleles was performed in 58 white patients with Crohn’s disease and 93 healthy controls using a polymerase chain reaction-sequence specific oligonucleotide based approach.
Results—No significant association with any DPB or DQB alleles was noted in patients with Crohn’s disease. Since our previous studies had shown a strong association of an HLA DRB3*0301/DRB1*1302 haplotype with Crohn’s disease, we re-examined this association using more stringent genotyping criteria. This haplotype was present in 20.7% of patients and 5.4% of controls (p = 0.0066; relative risk = 4.59).
Conclusions—The DRB3*0301/DRB1* 1302 haplotype is the only significant MHC class II association noted in white people with Crohn’s disease and represents the strongest association of any MHC or non-MHC locus with this disease.
- Crohn’s disease
- inflammatory bowel disease
- genetic susceptibility
- major histocompatibility complex
- HLA class II genes
Statistics from Altmetric.com
- Crohn’s disease
- inflammatory bowel disease
- genetic susceptibility
- major histocompatibility complex
- HLA class II genes
A combination of environmental and genetic factors presumably plays a role in the pathogenesis of Crohn’s disease. A susceptibility locus on chromosome 16 has been identified using a sib-pair analysis approach,1 ,2 and other non-MHC linked susceptibility loci are likely to be of importance as well.3 Given that activated inflammatory T helper cells are a prominent feature in Crohn’s disease, it is likely that HLA class II molecules play an important role in the disease process. Genetic susceptibility to Crohn’s disease linked to HLA class II genes has been most striking in studies from Japan. In Japanese patients with Crohn’s disease a strong positive association of the disease with DQB1*0401 and DQB1*0402 has been observed, and a negative association of Crohn’s disease with DQB1*0102 was noted.4
It remains unclear whether the inheritance of particular MHC class II alleles confers susceptibility to Crohn’s disease in the white population. Most studies on MHC class II genes in white patients with Crohn’s disease have involved the analysis of a limited number of alleles.3 ,5-9 In a recent study in which all DRB1, DRB3, DRB4, and DRB5 alleles were analysed using a polymerase chain reaction-sequence specific oligonucleotide (PCR-SSO) approach, we noted a strong association of Crohn’s disease with DRB3*0301.10A significant association of Crohn’s disease was also noted with DRB1*1302. Every patient and control who inherited the DRB1*1302 allele was also positive for DRB3*0301. This was not surprising given that these two alleles are known to be tightly linked in the white population.11 In all the other studies referred to above,3 ,5-9 the DRB1*1302 and DRB3*0301 alleles were not typed. In one recent European study in which more extensive typing was employed, a single MHC class II allele, DRB1*1302, was noted to be associated with disease12 (the linked DRB3*0301 allele was not typed). This study provided independent corroboration of our previous analysis of DRB alleles and susceptibility to Crohn’s disease.10
In this study we report the complete molecular typing of HLA class II DPB and DQB alleles in patients with Crohn’s disease and ethnically matched healthy white controls. Apart from seeking to explore if there was any linkage of the disease to DPB or DQB alleles, we were interested in determining if our previously reported DRB3*0301 and DRB1*1302 associations in Crohn’s disease reflected linkage disequilibrium to any DQB allele. In addition, our previous study had only used generic primers for the typing of the DRB3 locus, and complete concordance was not seen between DRB3*0301 positivity and the inheritance of DRB1*1302, which was somewhat surprising. We were concerned that the approach to DRB3 typing used previously was not stringent enough and that typing with locus specific primers would provide a more accurate assessment of DRB3 allele frequencies. We therefore wished to test more stringently (using additional typing approaches not used in our previous study) the null hypothesis that the DRB1*1302/DRB3*0301 haplotype is linked to Crohn’s disease in the white population.
Methods
PATIENTS
Fifty eight consecutive white patients with Crohn’s disease seen at the Center for the Study of Inflammatory Bowel Disease at Massachusetts General Hospital between January 1995 and November 1996 were included in the study. The diagnosis of Crohn’s disease was made by conventional endoscopic, histological, and clinical criteria. Information on ethnicity was obtained from each patient. Retrospective clinical information was obtained for comparison from most of the patients. Information analysed included the age of onset, the existence of a family history of inflammatory bowel disease, anatomical sites of disease, the presence of fistulae or strictures, steroid dependence of the disease, and the presence of extraintestinal manifestations such as arthritis, uveitis, or erythema nodosum. Forty two of these patients had been included in our previous study.10
CONTROLS
The 93 healthy controls have been described previously and were all whites.10 Information on ethnicity was also obtained from controls.
MHC CLASS II TYPING
Genotyping of all DRB, DPB1, and DQB1 alleles was performed using a PCR-SSO approach according to the protocols described in the 11th International HLA Workshop.13 Details of DRB typing have been described previously.10 Briefly, PCRs were performed in a total volume of 200 μl and included 3 μl genomic DNA, 200 pM each primer, 0.2 mM each deoxynucleotide triphosphate, 2 mM MgCl2,, 10 mM Tris/HCl, pH 8.0, 50 mM KCl, 0.001% (w/v) gelatin and 2.5 U Taq DNA polymerase (Fisher). Samples were denatured at 96°C for six minutes. Forty cycles of amplification sequentially involved incubation at 96°C for one minute followed by an annealing step for one minute (the temperature of which varied when different alleles were being analysed), and extension at 72°C for two minutes. After amplification, samples were incubated at 72°C for 10 minutes. The annealing temperature for the generic DRB1 alleles was 55 and 60°C for DRB1 and DRB3 group specific amplifications, 60°C for DP and 55°C for DQ. A 10 μl sample of the reaction mixture was run out on a 1.5% agarose gel, and the PCR products were viewed under UV light. Controls included previously typed samples. Generic typing of DRB1, DRB3, DRB4, and DRB5 loci was performed using a single pair of generic primers for PCR amplification followed by hybridisation of filters with 29 different group identifying SSOs. For group specific DRB typing, genomic DNA was amplified with five different 5’ primers specific for DRB1-DR1, DRB1-DR2, DRB1-DR4, the DRB1-DR52 associated group and the DRB3-DR52 group. The 3’ primer in the above cases was the same as that used for generic DRB typing. A total of 50 different SSOs was used in the group specific typing studies. Five SSOs were used for DRB1-DR1 (DRB1*0101-03), 12 for DRB1-DR2 (DRB1*1501-03, DRB1*160102), nine for DRB1-DR4 (DRB1*0401-0411), 12 for the DRB1-DR52 associated group (DRB1* 0301/02, DRB1*0801-0804, DRB1*1101-04, DRB1*1201/02, DRB1*1301-05, DRB1* 1401-08), and 12 SSOs were used for the DRB3-DR52 group (DRB3*0101, DRB3* 0201/0202 and DRB3*0301).
Generic primers were used for DPB1 and DQB1 typing; 25 SSOs were used to type 36 DPB1 alleles, and 20 SSOs for 17 DQB1 alleles. Details of the hybridisation conditions have been published.10 In brief, filters were prehybridised overnight at 54°C in buffer containing 3 M tetramethylammonium chloride, 50 mM Tris/HCl, pH 8.0, 2 mM EDTA, 5 × Denhardt’s solution, 0.1% sodium dodecyl sulphate (SDS), and 100 mg/ml salmon sperm DNA. Hybridisation was carried out using [γ-32P]ATP 5’ labelled SSOs at 54°C for two hours. The filters were washed twice at room temperature for 15 minutes each in a solution containing 2 × SSPE and 0.1% SDS followed by three washes for 10 minutes at 58°C in buffer containing 3 M tetramethylammonium chloride, 50 mM Tris/HCl, 2 mM EDTA, and 0.1% SDS. Each filter was exposed twice for autoradiography, once for one to two hours and subsequently for 14–16 hours. Reactivity was graded visually, using a scale recommended by the 11th International HLA Workshop.13 The use of multiple oligonucleotide probes facilitated definitive identification of negative and positive alleles.
ANALYSIS OF DRB3*0301
As noted above, in these studies we examined DRB3 alleles not only by the generic PCR amplification approach described previously10 but also using a separate 5’ primer to amplify DRB3 alleles. A total of 12 SSOs was used for DRB3 typing using the specific PCR approach. Of these 12, five reacted with DRB3*0301 and two were specific for DRB3*0301.
STATISTICAL ANALYSIS
Two tailed uncorrected p values were reported using Fisher’s exact test for the analyses of MHC class II allele frequencies and for most comparisons of clinical information between DRB1*1302/DRB3*0301 haplotype positive and negative patients. The relative risk was calculated as an odds ratio using the approximation of Woolf.14 For comparing mean ages of patients with Crohn’s disease who were positive or negative for the DRB1*1302/DRB3*0301 haplotype, an unpaired Student’s t test was employed. p values were corrected for the number of comparisons essentially using a modified Bonferroni correction as suggested by Svejgaard and Ryder.15 The number of alleles assayed from a given specific PCR amplification was used as the basis for the number of comparisons made. Alleles for which the combined frequency in patients and controls was less than 1 were not included in the number of comparisons. In the case of the DPB1 locus, the number of comparisons made was 20. Thus the nominal level for comparison was p⩽0.0025. In the case of the DQB1 locus, the number of comparisons made was 13 and the nominal level for comparison was p⩽0.0038. In the case of the DRB1 locus, the number of comparisons was 26 and the nominal level for comparison was p⩽0.0019. In the case of the DRB3, 4, and 5 loci, the number of comparisons was six and the nominal level for comparison was p⩽0.0083.
Results
Detailed molecular typing failed to disclose any statistically significant associations of DPB1 alleles with Crohn’s disease (table1). However, weak positive associations were noted with DPB1*0201 (p = 0.0702) and DPB1*0501 (p = 0.0509), and a weak negative association was noted with DPB1*0402 (p = 0.0783). Some of these associations may attain significance in a study with more power. Analysis of DQB alleles (table 2) showed weak associations in Crohn’s disease with DQB1*0201 (p = 0.0168) and DQB1*0605 (p = 0.0226). We believe that these weak DQB associations reflect the known linkage of these DQB alleles to DRB1*1302 in the white population.16 Weak negative associations were also noted with DQB1*03032 (p = 0.0883) and DQB1*05031 (p = 0.0883); these might acquire more significance in a larger study.
DPB1 alleles in patients with Crohn’s disease and controls
DQB1 alleles in patients with Crohn’s disease and controls
On this extended series of patients, a weak negative association of DRB1*0401 (p = 0.0173) was noted in patients with Crohn’s disease (table 3). This association ceases to be statistically significant after corrections are made for the number of comparisons (adjusted nominal level is p⩽0.0019). This negative association may attain more significance in a larger study. One of the major goals of this study was to test the null hypothesis that Crohn’s disease in whites is linked to DRB3*0301 or to a DRB3*0301/DRB1*1302 haplotype. In our previous report we had analysed the DRB3*0301 allele only using generic primers in the amplification step. We had used one set of generic primers to analyse DRB1 alleles and the DRB3, 4, and 5 loci, before evaluating PCR products with a range of probes. Although subsequent specific PCRs were performed to assay groups of DRB1 alleles, specific amplification of the DRB3 locus had not been performed. While every subject in that study who tested positive for the DRB1*1302 allele was also positive for DRB3*0301, a few patients who were typed for DRB3*0301 apparently did not carry a DRB1*1302 allele. Given the known tight linkage of DRB1*1302 to DRB3*0301, this lack of concordance was puzzling. We sought to assay for DRB3*0301 more stringently in this extended group of patients with Crohn’s disease and in controls. Accordingly, we performed a specific PCR for DRB3 alleles and probed this PCR product with 12 separate SSOs as described under Methods. Using these criteria every patient or control inheriting a DRB1*1302 allele was also noted to have inherited a DRB3*0301 allele (tables 3and 4). Four patients who had previously been categorised as being DRB3*0301 positive in our previous study10 but who were negative for the DRB1*1302 allele were found to be negative for DRB3*0301 using the more stringent criteria described above. A strong association with the DRB1*1302/DRB3*0301 haplotype was noted in patients with Crohn’s disease (p = 0.0066; relative risk 4.59). Given that the DRB1*1302/DRB3*0301 haplotype was specifically targeted for analysis by our initial hypothesis, corrections for the number of comparisons made were not called for. Nonetheless, the above p value (p = 0.0066) is less than the corrected nominal value of p⩽0.0083 (for the DRB3 locus), and it should be considered that individuals with this haplotype are at a significantly increased risk for Crohn’s disease. Figure 1 shows representative assays for the molecular typing of HLA DP, DQ, and DR alleles.
DRB1 alleles in patients with Crohn’s disease and controls
DRB3 DRB4 and DRB5 alleles in patients with Crohn’s disease and controls
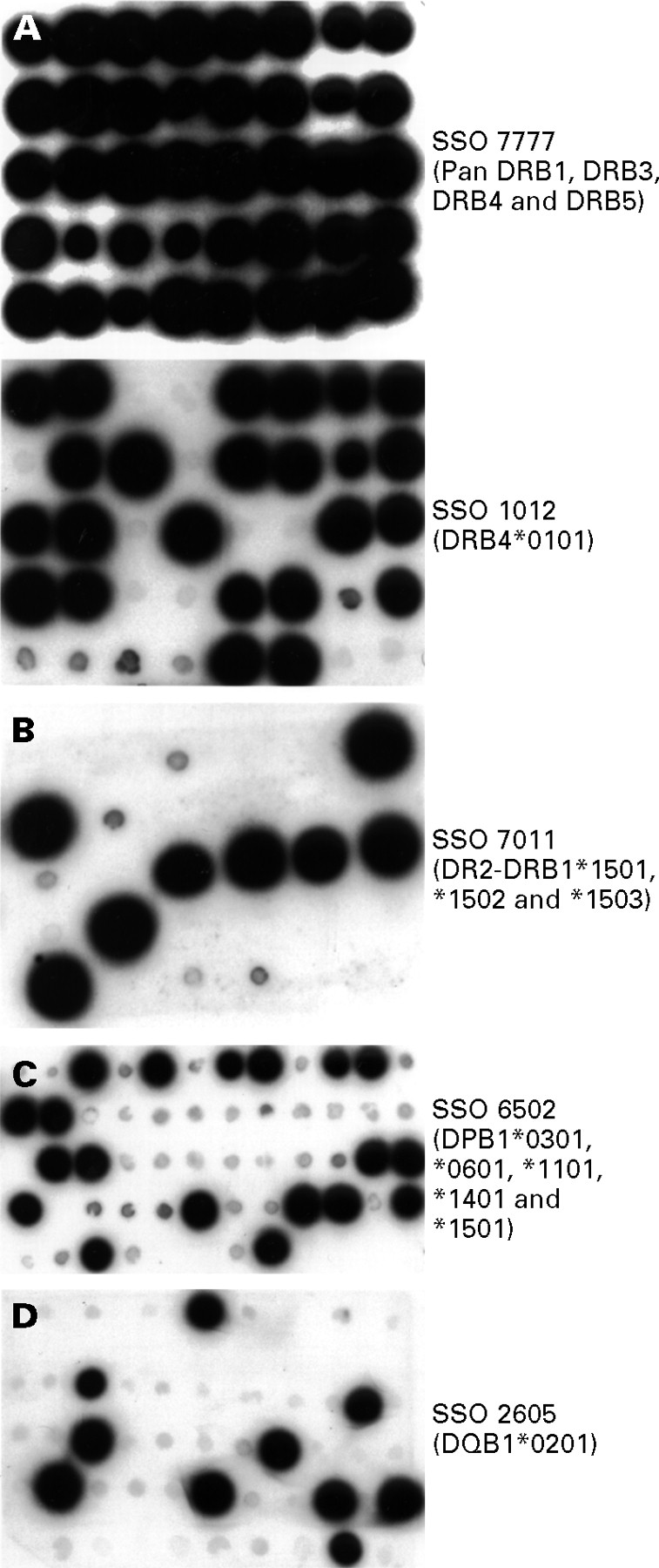
{kind=link}
Representative SSO-PCR analyses for HLA DP, DQ, and DR alleles. (A) A representative filter depicting generic DRB typing for groups of DRB1, DRB3, DRB4, and DRB5 alleles. Upper panel: dot blot hybridisation of a representative filter with a pan DRB probe to show the presence of amplified DNA in each spot on replicate filters. A total of 40 Crohn’s disease and control samples were spotted on this filter. Lower panel: a replicate of the filter depicted in the upper panel was probed with an SSO specific for DRB4*0101. Amplified DNA that hybridised or failed to hybridise with the SSO may be visually distinguished (24 of the 40 spots containing amplified DNA hybridised with this probe). (B) A representative filter depicting group specific DRB typing for the DRB1-DR2 group. A separate PCR amplification for the DRB1-DR2 group was performed and a representative filter was probed with an SSO that is specific for some members of this group (DRB1*1501, DRB1*1502, and DRB1*1503). Multiple SSO probes were used to type all the alleles in each group. Eight of the 27 samples were revealed by this probe. (C) A representative filter depicting generic DPB1 typing using an SSO that can reveal five DPB1 alleles; 19 of 54 samples (from normal controls) were hybridised with this specific probe. (D) A representative filter depicting generic DQ typing of normal controls. This filter was probed with an SSO specific for DQB1*0201; 10 of 50 samples were revealed by this probe.
It has been pointed out that susceptiblity alleles for inflammatory bowel disease may vary between ethnic groups.17 Compared with other North American studies, a larger proportion of our white patients with Crohn’s disease are non-Jewish. However, the frequency of the DRB1*1302/DRB3*0301 haplotype in Jewish patients with Crohn’s disease (13.33%, n = 15) and non-Jewish patients with Crohn’s disease (23.25%, n = 43) was not significantly different (p = 0.7123). We have attempted to correlate the inheritance of this haplotype in our patients with a number of clinical parameters listed in table 5. No significant correlation was seen with the anatomical extent of the disease, the presence of stricturing or fistulating disease, steroid dependency, or the presence of extraintestinal manifestations. A trend was noted towards an earlier age of onset (mean age 22.7 years) in patients who inherited this haplotype than in those who did not (mean age 29.4 years). The two tailed p value (derived by an unpaired Student’s t test) for this comparison was 0.18, which is not significant.
Comparison of clinical characteristics of DRB1*1302/DRB3*0301 positive and negative patients with Crohn’s disease
Table 6 summarises selected results of weak and strong HLA class II associations with Crohn’s disease.
Summary of HLA class II associations in white patients with Crohn’s disease
Discussion
The lesions in Crohn’s disease result from the activation of inflammatory T cells by peptides presented by MHC class II molecules which may or may not be of host origin. It is of some interest that persistent clonal expansions of peripheral CD4 T cells have been noted in patients with Crohn’s disease.18 ,19 Given the importance of T cell mediated inflammation in Crohn’s disease, there is little doubt that MHC class II molecules play a critical role in the disease process. Whether or not specific MHC class II alleles are linked to the disease is another issue.
Making confident assertions about the presence or absence of the association of MHC class II alleles with a disease depends not only on proper patient selection, but also on the accurate typing of all known alleles at each of the polymorphic HLA class II loci. The ideal study would involve the analysis of all known DP, DQ, and DR alleles in a very large number of carefully characterised patients and controls. The larger the number of subjects the more likely less frequent associations will be detected. Even in a study of a large number of patients, if an allele that is associated with disease in a fair proportion of patients is not typed, such an association will obviously be missed. This is an important limitation of a number of previous studies on MHC genes and susceptibility to Crohn’s disease. The present report represents the most comprehensive analysis of DP, DQ, and DR alleles in this disease.
Although we noted no significant association of Crohn’s disease with DPB or DQB alleles, it is possible that these alleles present peptides of pathogenic significance in a relatively small number of patients. In a substantial proportion of white patients with Crohn’s disease, the disease is clearly linked to inheritance of the DRB1*1302/DRB3*0301 haplotype. Our more stringent analyses in this study have confirmed the findings in our previous report.10 In that study, association of Crohn’s disease with the DRB1*1302/DRB3*0301 haplotype was actually less significant (p = 0.0234; relative risk 4.1412) than the association seen in the present study. The association seen with DRB3*0301 alone in that study appeared stronger (p = 0.0004; relative risk 7.04), but we recognise that these results were inaccurate and reflect the less stringent methods used at the time for the analysis of the DRB3*0301 allele.
It is of interest that in the only other study on white patients with Crohn’s disease in which the DRB1*1302 allele was actually typed,12 this proved to be the only allele associated with disease. In that study by Heresbach et al,12 additional susceptibility loci were identified by examining clinical subgroups of patients with Crohn’s disease. In our continuing studies we will seek to analyse MHC class II associations in similar clinical categories of subjects drawn from a larger set of patients with Crohn’s disease.
The detection of associations between the inheritance of specific alleles and disease was limited in this study by the number of subjects studied; we note that on the basis of 58 cases and a two sided 0.05 level test, we had about 80% power to detect a difference of 20% in the frequency of inheritance of a given haplotype in patients versus controls (for example 25% in patients versus 5% in controls). The power to detect differences of 15% was only about 60%. In our study therefore we were able to detect only strong associations. When the Bonferroni correction is made for the number of comparisons, the power is much lower, suggesting that the associations actually noted are likely to be seen as even more significant in a larger study. It is therefore possible that a number of additional significant associations of MHC class II alleles with Crohn’s disease (other than the strong DRB3*0301/DRB1*1302 association confirmed in this study) will be noted in a study with more power.
An association between an HLA class II gene and a disease syndrome does not necessarily imply that the linked class II heterodimeric product is directly involved in the disease process. In principle, such an association may reflect linkage to another HLA gene, linkage to a non-HLA gene that is relevant to immune function, or even to some other gene on chromosome 6 which has no direct bearing on the immune system. An example of the latter category is the linkage disequilibrium between a haplotype including HLA B47 and the 21-hydroxylase gene on chromosome 6 in patients with one form of congenital adrenal hyperplasia.20 In a number of autoimmune or chronic inflammatory diseases, inflammation is driven by helper T cells and susceptibility is linked to multiple loci, most of which are not on chromosome 6. In diseases that fall into this category, strong associations with a limited number of MHC class II alleles have generally been noted. In white patients with multiple sclerosis, susceptibility to the disease is often linked to DRB1*1501,21 ,22 and in type I diabetes suceptibility is associated primarily with the DRB1*04/DQB1*0302 and DRB1*0301/DQB1*0201 haplotypes.23 ,24 In these conditions, self antigen derived peptides are presented by these disease associated HLA class II molecules to helper T cells which are likely to be of pathogenic importance.25 ,26 There is growing evidence for the abrogation of T cell tolerance in such states by a process of molecular mimicry initiated by specific infectious agents which may trigger self reactive T cells.27 Although there is no convincing evidence to suggest that Crohn’s disease is an autoimmune disease, it is nevertheless likely that inflammation is driven by helper T cells responding to peptides of pathogenic significance presented by disease related HLA class II molecules. An alternative possibility is that disease linked HLA class II molecules may not directly be linked to the induction of inflammation in Crohn’s disease, but that the inheritance of a susceptibility allele may predispose individuals to infection by some unidentified microbial agent. This infection may, in turn, be responsible for the induction of disease related inflammatory T cells by molecular mimicry.
One of the indirect conclusions that we can draw from this study is that peptides that may be of pathogenic significance in Crohn’s disease in the white population may be presented by a large number of MHC class II alleles that are fairly widely distributed in the population. Certain peptides of disease significance may be preferentially presented by either DRB3*0301 or DRB1*1302. Linkage to this haplotype may reflect an actual role for one or both of these alleles, but may also reflect linkage disequilibrium to some other non-MHC gene. It clearly does not represent linkage disequilibrium to an MHC class II DQB allele. In any event, the association that we note with this particular haplotype represents a much stronger association (in terms of its frequency in affected white patients with Crohn’s disease) than any non-MHC linked locus currently identified. Given the association of disease with specific susceptibility alleles in a less heterogeneous Japanese Crohn’s disease population,4 it is quite likely that this result reflects a real MHC class II susceptibility haplotype for white patients with Crohn’s disease.
Acknowledgments
We thank all the patients who volunteered for this study and their referring clinicians. This work was supported by grants AI-33507 and P30DK43351 from the National Institutes of Health and by the core facilities of the Center for the Study of Inflammatory Bowel Disease, Massachusetts General Hospital.