Article Text

Abstract
Background and aims: A characteristic feature of Crohn’s disease (CD) is mesenteric adipose tissue hypertrophy. Mesenteric adipocytes or specific proteins secreted by them may play a role in the pathogenesis of CD. We recently identified adiponectin as an adipocyte specific protein with anti-inflammatory properties. Here we report on expression of adiponectin in mesenteric adipose tissue of CD patients.
Methods and results: Mesenteric adipose tissue specimens were obtained from patients with CD (n = 22), ulcerative colitis (UC) (n = 8) and, for controls, colon carcinoma patients (n = 28) who underwent intestinal resection. Adiponectin concentrations were determined by enzyme linked immunosorbent assay, and adiponectin mRNA levels were determined by real time quantitative reverse transcription-polymerase chain reaction. Tissue concentrations and release of adiponectin were significantly increased in hypertrophied mesenteric adipose tissue of CD patients compared with normal mesenteric adipose tissue of CD patients (p = 0.002, p = 0.040, respectively), UC patients (p = 0.002, p = 0.003), and controls (p<0.0001, p<0.0001). Adiponectin mRNA levels were significantly higher in hypertrophied mesenteric adipose tissue of CD patients than in paired normal mesenteric adipose tissue from the same subjects (p = 0.024). Adiponectin concentrations in hypertrophied mesenteric adipose tissue of CD patients with an internal fistula were significantly lower than those of CD patients without an internal fistula (p = 0.003).
Conclusions: Our results suggest that adipocytes in hypertrophied mesenteric adipose tissue produce and secrete significant amounts of adiponectin, which could be involved in the regulation of intestinal inflammation associated with CD.
- CD, Crohn’s disease
- UC, ulcerative colitis
- TNF-α, tumour necrosis factor-α
- IL, interleukin
- BMI, body mass index
- ELISA, enzyme linked immunosorbent assay
- RT-PCR, reverse transcription-polymerase chain reaction
- cDNA, complementary DNA
- CRP, C reactive protein
- PPARγ, peroxisome proliferator activated receptor γ
- Crohn’s disease
- fat wrapping
- mesenteric adipocyte
- adiponectin
Statistics from Altmetric.com
- CD, Crohn’s disease
- UC, ulcerative colitis
- TNF-α, tumour necrosis factor-α
- IL, interleukin
- BMI, body mass index
- ELISA, enzyme linked immunosorbent assay
- RT-PCR, reverse transcription-polymerase chain reaction
- cDNA, complementary DNA
- CRP, C reactive protein
- PPARγ, peroxisome proliferator activated receptor γ
Crohn’s disease (CD) is a chronic inflammatory bowel disease of the gastrointestinal tract. As its aetiology is still unknown, CD is identified and diagnosed by the appearance of a set of clinical, endoscopic, histological, and macroscopic characteristics. Transmural inflammation is the histological hallmark of CD. Occasionally, inflammation extends beyond the intestinal wall, and an internal fistula or an intra-abdominal abscess is formed. The macroscopic features of the mesentery adjacent to the intestinal segments involved in CD often include hypertrophied adipose tissue such as fat wrapping and mesenteric thickening.1–5 Fat wrapping, defined as hypertrophied adipose tissue extending from the mesenteric attachment and partially covering the intestinal circumference, is common in both the small and large intestine and is also considered a hallmark of CD.1 While considerable variation exists in the presentation of the disease in surgical specimens, CD is often recognised solely by the appearance of these macroscopic lesions of the mesentery.4 Their aetiology and significance, however, remain unknown.
While it is well known that adipose tissue serves as an energy reservoir, recent advances in cell biology have shown that adipocytes produce and secrete several bioactive molecules which are collectively referred to as adipocytokines. We previously identified an adipocyte specific secretory protein, adiponectin, the gene transcript of which was most abundant in the expression profiles of human adipose tissue.6 Adiponectin is composed of 244 amino acid residues containing a short non-collagenous N terminal segment followed by a collagen-like sequence. It belongs to the family of proteins that include C1q and the collectins,6,7 which play important roles in the innate humoral immune system.8–10 We and others have shown that adiponectin modulates a wide array of biological functions. Adiponectin was demonstrated to be related to improved insulin resistance and prevention of atherosclerosis, fatty liver, and liver fibrosis.11–19 In addition to the above actions, we reported that adiponectin has several anti-inflammatory effects. For example, adiponectin reduces monocyte attachment to cultured human aortic endothelial cells by inhibiting expression of vascular cell adhesion molecules, intercellular adhesion molecules, and E-selectin.20 Adiponectin inhibits the phagocytic activity and production of tumour necrosis factor (TNF)-α and interleukin (IL)-6 in cultured macrophages.7,21 These actions may be mediated by suppression of nuclear factor κB signalling and extracellular signal regulated kinase 1/2 activity.21,22 Adiponectin knockout mice display high levels of TNF-α mRNA in adipose tissue and high concentrations in plasma, which decrease by supplementation of plasma adiponectin.12 Furthermore, we recently reported that adiponectin enhances the production of IL-10 and tissue inhibitor of metalloproteinase 1 in human cultured macrophages.23 Thus adiponectin exhibits anti-inflammatory properties, especially in endothelial cells and macrophages.
To our knowledge, the role of adiponectin in the pathogenesis of CD has not been examined previously. The present study was designed to determine expression levels of adiponectin in hypertrophied mesenteric adipose tissue, and its relevance to the pathogenesis of CD.
MATERIALS AND METHODS
Patients and specimen preparation
The study protocol was approved by the Human Ethics Review Committee of Osaka University School of Medicine and a signed consent form was obtained from each subject. Mesenteric adipose tissue specimens were obtained from 22 patients with CD, eight with ulcerative colitis (UC), and 28 with colon cancer who underwent surgical resection. CD patients (16 men, six women; mean age 34.4 (1.8) years (range 24–56); mean body mass index (BMI) 18.8 (0.4) kg/m2) underwent ileal or colonic resection because of stenosis or an internal fistula. Three patients with CD were treated by steroids. The other patients were not on any specific medical treatment at the time of the study. Eleven patients with CD had pure ileal involvement, 10 had ileocolonic CD, and one had colonic CD. Ten patients with CD had an internal fistula. UC patients (five men, three women; mean age 49.4 (5.2) years (range 32–69); mean BMI 21.4 (0.8) kg/m2) underwent intestinal resection because of steroid dependency or lack of response to medication. Patients with colon cancer (19 men, nine women; mean age 67.3 (1.3) years (range 53–75); mean BMI 22.4 (0.6) kg/m2) were enrolled as the control group. All specimens were taken from mesenteric adipose tissue close to the intestinal wall. In 16 of 22 CD patients, paired samples were obtained from hypertrophied mesenteric adipose tissue adjacent to the involved intestine and apparently normal mesenteric adipose tissue contiguous with the healthy segment of the intestine. Resected specimens were immediately frozen in liquid nitrogen and stored −80°C until assayed. Some samples were fixed in 10% phosphate buffered formalin and embedded in paraffin for histopathological studies.
Histopathological analysis
Haematoxylin and eosin staining was routinely performed. For quantitation of the size and number of adipocytes, sectional areas of adipose tissue were analysed with an image analysis system (Macscope version 2.59; Mitani, Fukui, Japan). Tissue samples were immunohistologically stained for CD3, CD20, and CD68 using the labelled streptavidin-biotin system (Dako, Glostrup, Denmark). Mouse monoclonal antihuman CD68, CD3, and CD20 antibodies were purchased from Nichirei (Tokyo, Japan). For immunohistological staining of adiponectin, paraffin sections were deparaffinised, dehydrated, blocked with normal goat serum (Vector Laboratories, Burlingame, California, USA) for 10 minutes at room temperature, and then incubated at 4°C overnight with 5 μg/ml of a mouse monoclonal antihuman adiponectin antibody, ANOC 9104.22 Sections were then rinsed with 0.05 M Tris HCl buffer (pH 7.6), and incubated for 30 minutes with a 1:200 dilution of Alexa Fluor 594 goat antimouse IgG (H+L) (Molecular Probes, Eugene, Oregon, USA) at room temperature. As a negative control, the primary antibody was replaced by normal mouse serum (Dako). Nuclear counterstaining was carried out with a 1:5000 dilution of 4′, 6-diamidino-2-phenylindole (Molecular Probes) for five minutes at room temperature.
Determination of adiponectin and IL-6 concentrations in mesenteric adipose tissue
Mesenteric adipose tissue samples were homogenised in lysis buffer containing 20 mM Tris (pH 8.0), 137 mM NaCl, 10% glycerol, 1% Nonidet P-40, 10 mM ethylenediaminetetra acetic acid, 100 mM NaF, 1 mM phenylmethylsulfonylfluoride, 0.25 TIU/ml aprotinin, and 10 μg/ml leupeptin. Homogenates were incubated for 10 minutes at 4°C and then centrifuged at 16 000 g for 15 minutes at 4°C. The supernatants were collected and stored at −30°C until assayed. Adiponectin concentrations in homogenate supernatants of adipose tissues were determined by enzyme linked immunosorbent assay (ELISA), as described previously.24 IL-6 concentrations were measured using ELISA kits (BioSource International, Camarillo, California, USA). Total protein concentration was then determined using BCA Protein Assay Reagent (Pierce, Rockford, Illinois, USA) according to the manufacturer’s instructions. The final concentrations were expressed in ng of adiponectin and pg of IL-6 per mg of total protein.
Adiponectin release in short term cultures
Short term cultures were prepared with surgical specimens to examine release of adiponectin from the mesenteric adipose tissue. Adipose tissue samples (250 mg, wet weight) were diced finely and incubated in 2 ml of culture medium at 37°C in a 95% O2–5% CO2 humidified incubator for 24 hours, as described previously.25 Conditioned media were collected and stored at −30°C until used for assay. Adiponectin concentrations in conditioned media were then determined by ELISA.
Quantitation of adiponectin mRNA in mesenteric adipose tissue
Total RNA was extracted and reverse transcribed into complementary DNA (cDNA) as described previously.24 Real time quantitative polymerase chain reaction (PCR) was carried out on cDNA as described previously.14 The sequences of the primers and the probe for adiponectin were as follows: adiponectin sense primer, 5′-AGG TTG GAT GGC GGG C-3′; adiponectin antisense primer, 5′-TTT CAC CGA TGT CTC CCT TAG G-3′; adiponectin probe, 5′-(FAM)-TGG CAG AGA TGG CAC CCC TGG-(TAMRA)-3′. The primers and probe for β-actin were purchased from Applied Biosystems (Foster City, California, USA). The quantity of adiponectin was expressed as the ratio of adiponectin cDNA molecules to β-actin cDNA molecules.
Statistical analysis
Data are expressed as mean (SEM). Differences among the four groups were studied using the Kruskal-Wallis test for global comparisons and the Mann-Whitney test for paired comparisons. Paired and unpaired t tests were used to compare data between two groups when they showed normal distribution and the Mann-Whitney test when they were asymmetrical. Spearman’s test was applied to test for correlations. A p value <0.05 denoted the presence of a statistically significant difference.
RESULTS
Analysis of inflammatory cells and adipocytes in mesenteric adipose tissue
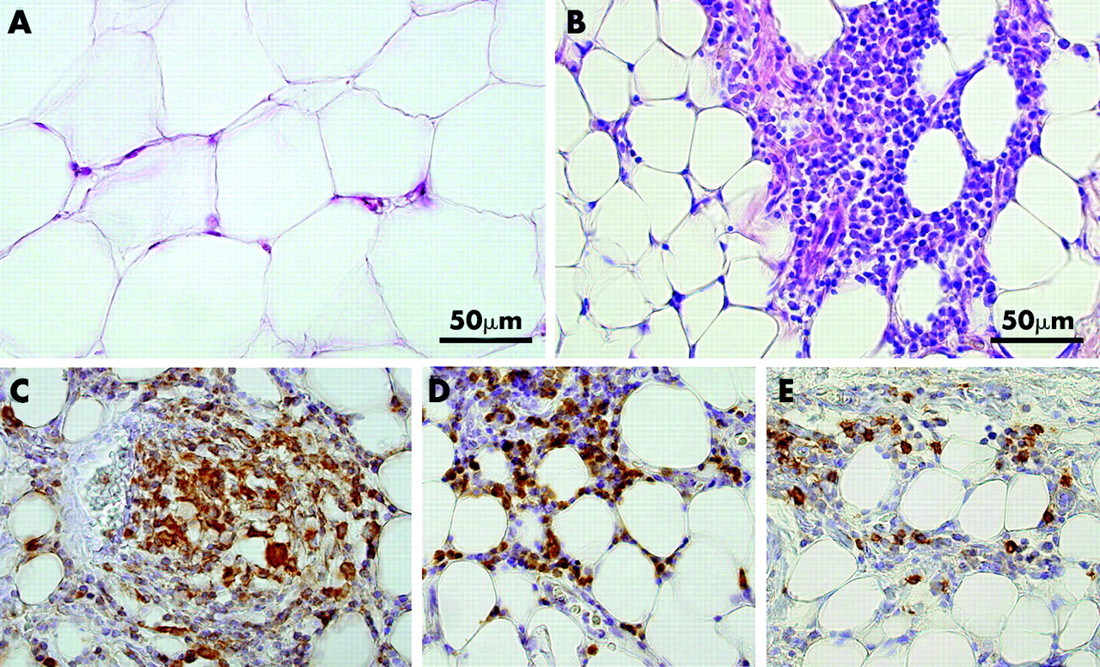
Histopathological examination showed a large number of inflammatory cells in hypertrophied mesenteric adipose tissue adjacent to the involved intestine of CD patients (fig 1B). In contrast, very few inflammatory cells were detected in normal mesenteric adipose tissue adjacent to the healthy intestine of CD patients, the intestines of UC patients, and the intestines of control subjects (fig 1A). Immunohistochemical staining using mouse monoclonal antibodies to CD68, CD3, and CD20 showed many CD68 positive (specific for monocytes/macrophages), CD3 positive (specific for T cells), and CD20 positive (specific for B cells) cells near the serosa in hypertrophied mesenteric adipose tissue of CD patients (fig 1C–E). The number of inflammatory cells decreased with increased distance from the serosa. The number of CD20 positive cells appeared to be less than other inflammatory cells. Quantitative analyses indicated that the size of adipocytes in hypertrophied mesenteric adipose tissue of CD patients (956 (21) μm2) was approximately one quarter the size of those in mesenteric adipose tissue from controls (4099 (174) μm2) (fig 1B, A). On the other hand, the number of adipocytes per unit area in hypertrophied mesenteric adipose tissue of CD patients was 3.5-fold that of adipocytes in the mesenteric adipose tissue from controls (603 v 173 per 900 000 μm2).

Haematoxylin-eosin (H&E) staining and immunohistochemical detection of CD68, CD3, and CD20 in hypertrophied mesenteric adipose tissue contiguous with involved intestinal segments of a representative patient with Crohn’s disease (CD). Mesenteric adipose tissue in a normal control subject (A) and hypertrophied mesenteric adipose tissue of a CD patient (B) were stained with H&E. Note that the size of adipocytes in hypertrophied mesenteric adipose tissue is approximately one quarter those of controls (A, B). CD68, CD3, and CD20 expressing cells were stained dark brown (C–E). Many CD68, CD3, and CD20 positive cells were present near the serosa in hypertrophied mesenteric adipose tissue. CD68 positive cells formed a non-caseous epithelioid cell granuloma (C). Nuclear counterstaining: Mayer’s haematoxylin. Original magnification ×200.
Immunostaining of adiponectin in mesenteric adipose tissue
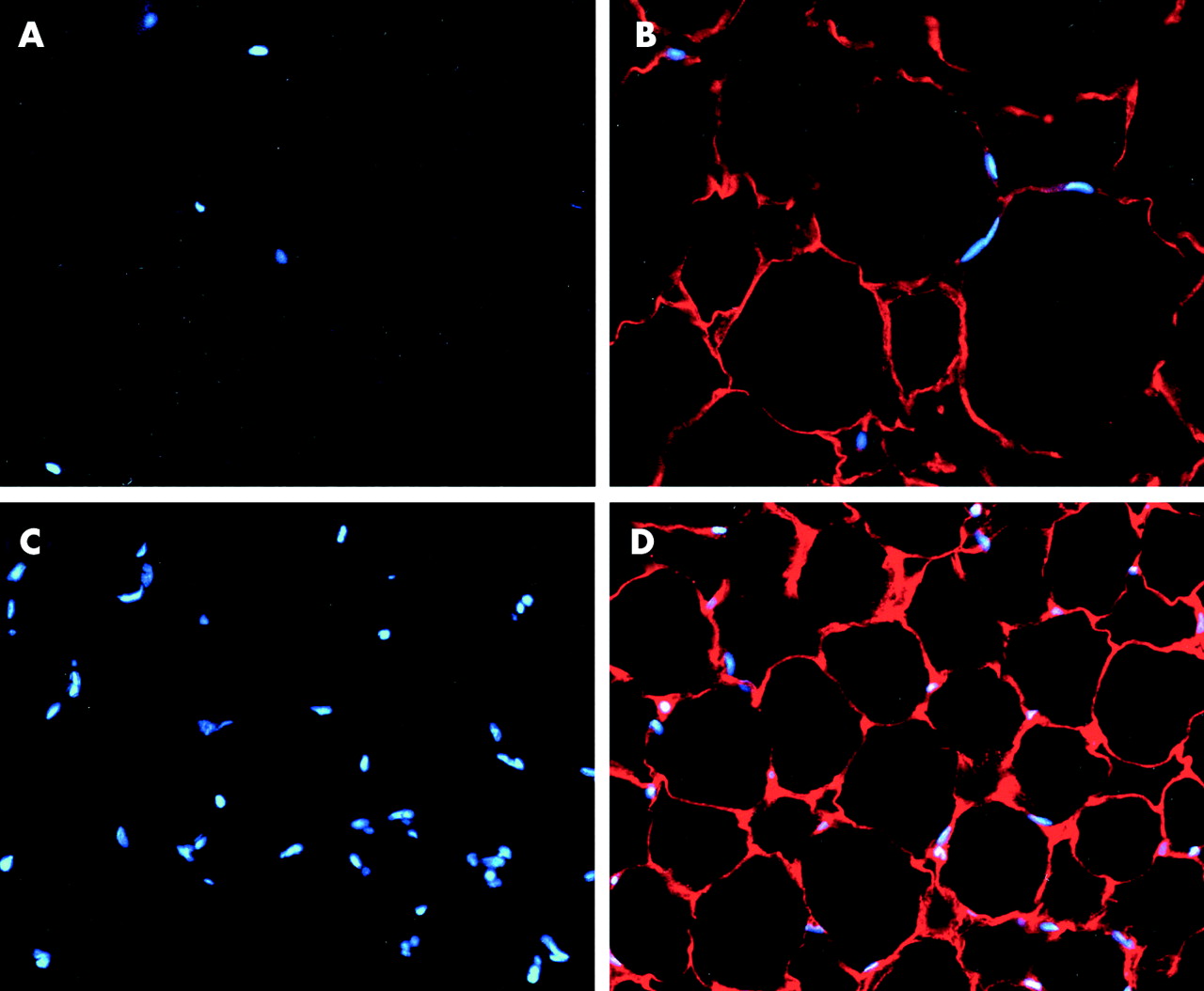
Adiponectin immunoreactivity appeared as a red fluorescence rim at the periphery of the cytoplasm of adipocytes in the mesenteric adipose tissue of controls and CD patients (fig 2B, D). In addition, sections incubated with normal mouse serum instead of mouse antihuman adiponectin antibody (fig 2A, C) or mouse antihuman adiponectin antibody preabsorbed with sufficient adiponectin showed no positive staining reaction (data not shown).

Immunohistochemical staining of adiponectin in the mesenteric adipose tissue of a representative control and Crohn’s disease (CD) patient. The mesenteric adipose tissue of the control (B) and the hypertrophied mesenteric adipose tissue of a CD patient (D) reacted with antiadiponectin antibody. No reaction product was detected against non-immune serum in the mesenteric adipose tissue of the control (A) and hypertrophied mesenteric adipose tissue of the CD patient (C). Adiponectin staining appeared as a red fluorescence rim in the periphery of the cytoplasm of adipocytes (B, D). Nuclear counterstaining: 4′, 6-diamidino-2-phenylindole. Original magnification: ×200.
Adiponectin concentration in mesenteric adipose tissue
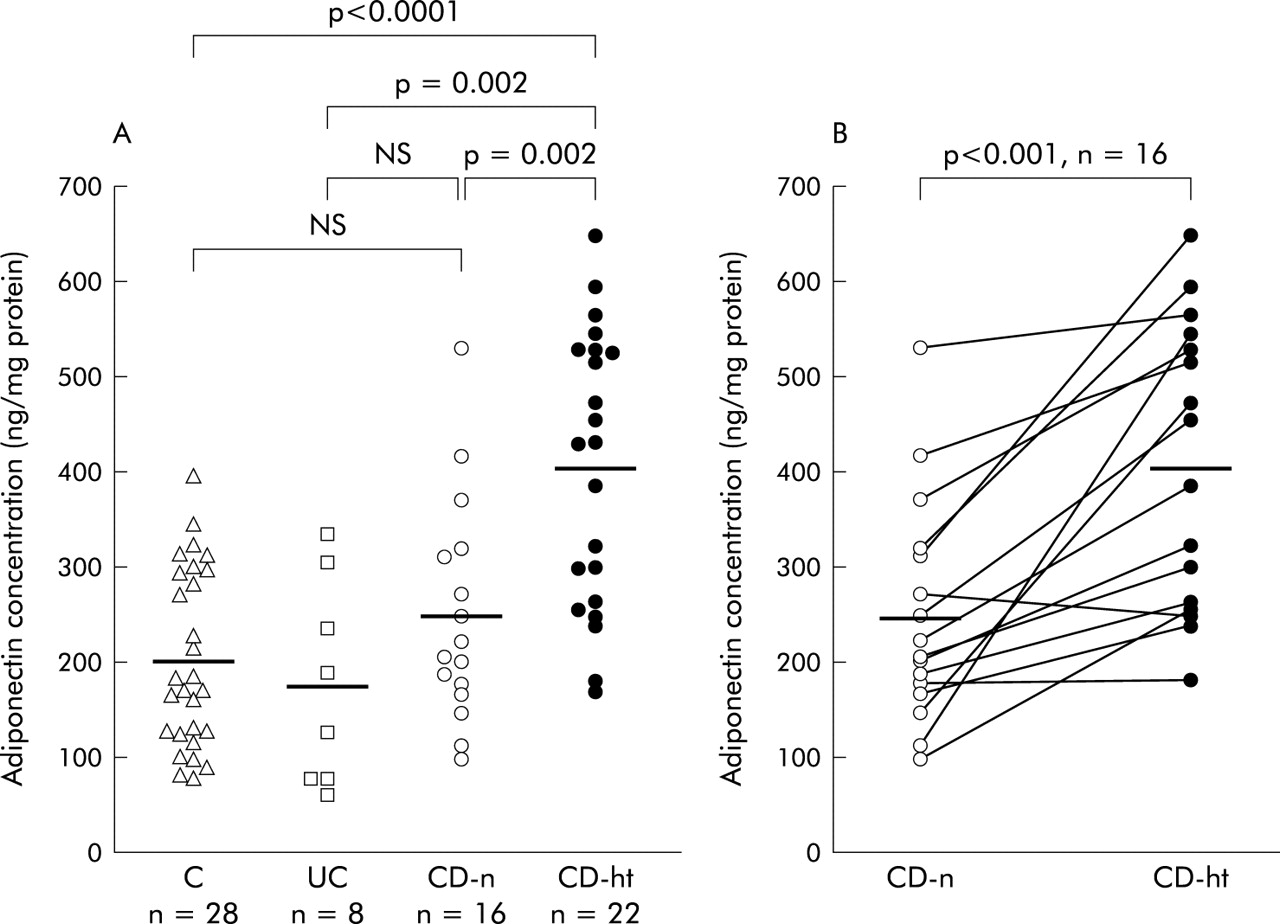
The mean adiponectin concentration in hypertrophied mesenteric adipose tissue of CD patients (404.7 (31.9) ng/mg) was significantly higher than that in normal mesenteric adipose tissue of controls (202.5 (17.7) ng/mg, p<0.0001), UC patients (176.1 (37.9) ng/mg, p = 0.002), or CD patients (249.3 (29.1) ng/mg, p = 0.002) (fig 3A). Moreover, adiponectin concentrations in hypertrophied mesenteric adipose tissues (407.4 (37.5) ng/mg) were also significantly higher than those in paired normal mesenteric adipose tissues (249.3 (29.1) ng/mg) from the same CD patients (p<0.001) (fig 3B).

Adiponectin concentrations in mesenteric adipose tissue. Adiponectin concentrations were determined by enzyme linked immunosorbent assay (ELISA) and values expressed in ng of adiponectin per mg of total homogenate protein. (A) Data are individual and mean values (horizontal bar) of adiponectin concentrations in normal mesenteric adipose tissues of controls (C), ulcerative colitis patients (UC), Crohn’s disease patients (CD-n), and hypertrophied mesenteric adipose tissues of CD patients (CD-ht). (B) Data are individual and mean values (horizontal bars) of adiponectin concentrations in normal mesenteric adipose tissues of CD patients (CD-n) and hypertrophied mesenteric adipose tissues of CD patients (CD-ht) from the same patients.
Adiponectin release from mesenteric adipose tissue
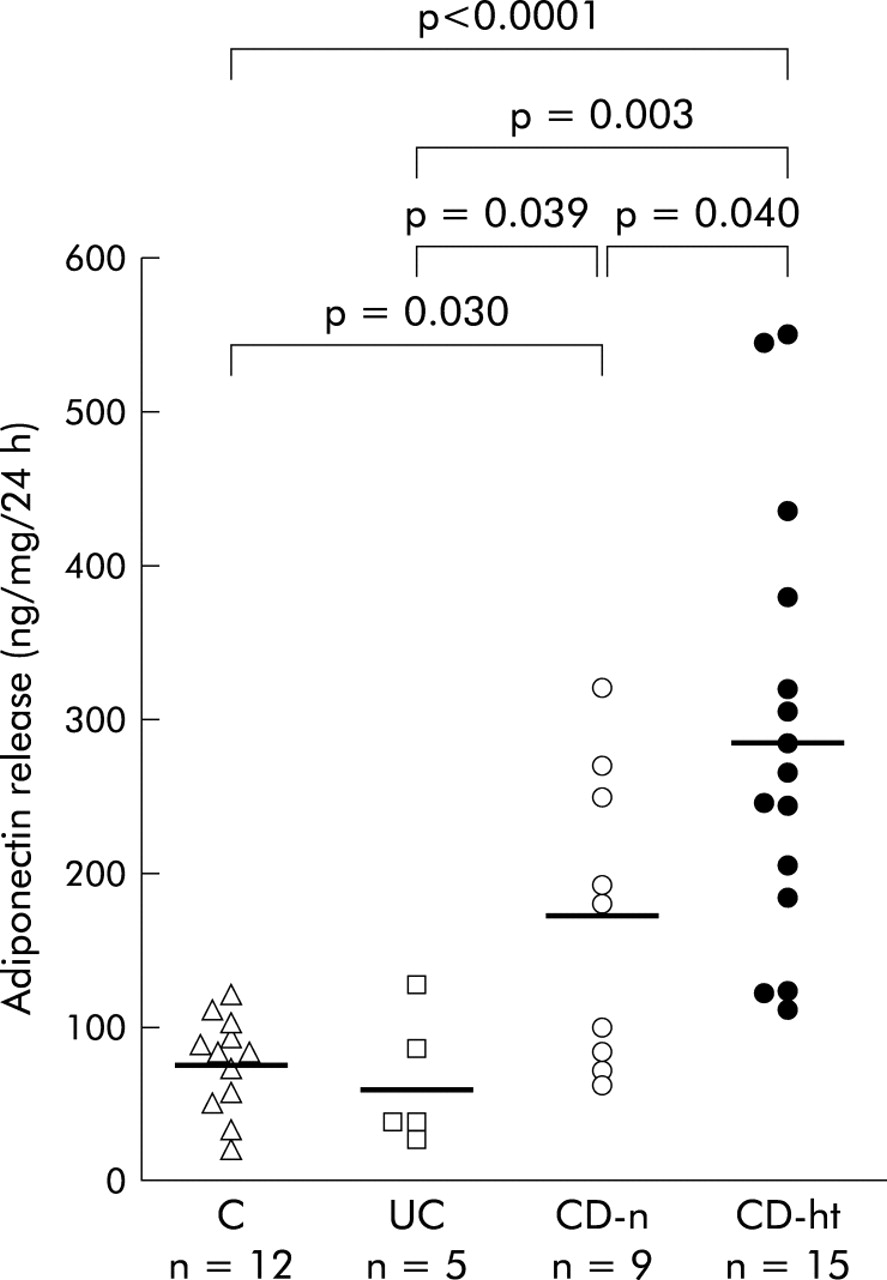
The amount of adiponectin released from hypertrophied mesenteric adipose tissue of CD patients (287.9 (36.0) ng/ml) was significantly higher than that from normal mesenteric adipose tissue of controls (76.0 (8.9) ng/ml, p<0.0001), UC patients (63.4 (19.0) ng/ml, p = 0.003), or CD patients (170.2 (31.9) ng/ml, p = 0.040) (fig 4). Furthermore, adiponectin release from normal mesenteric adipose tissue of CD patients was significantly higher than that from the mesenteric adipose tissue of control subjects (p = 0.030) or UC patients (p = 0.039) (fig 4).
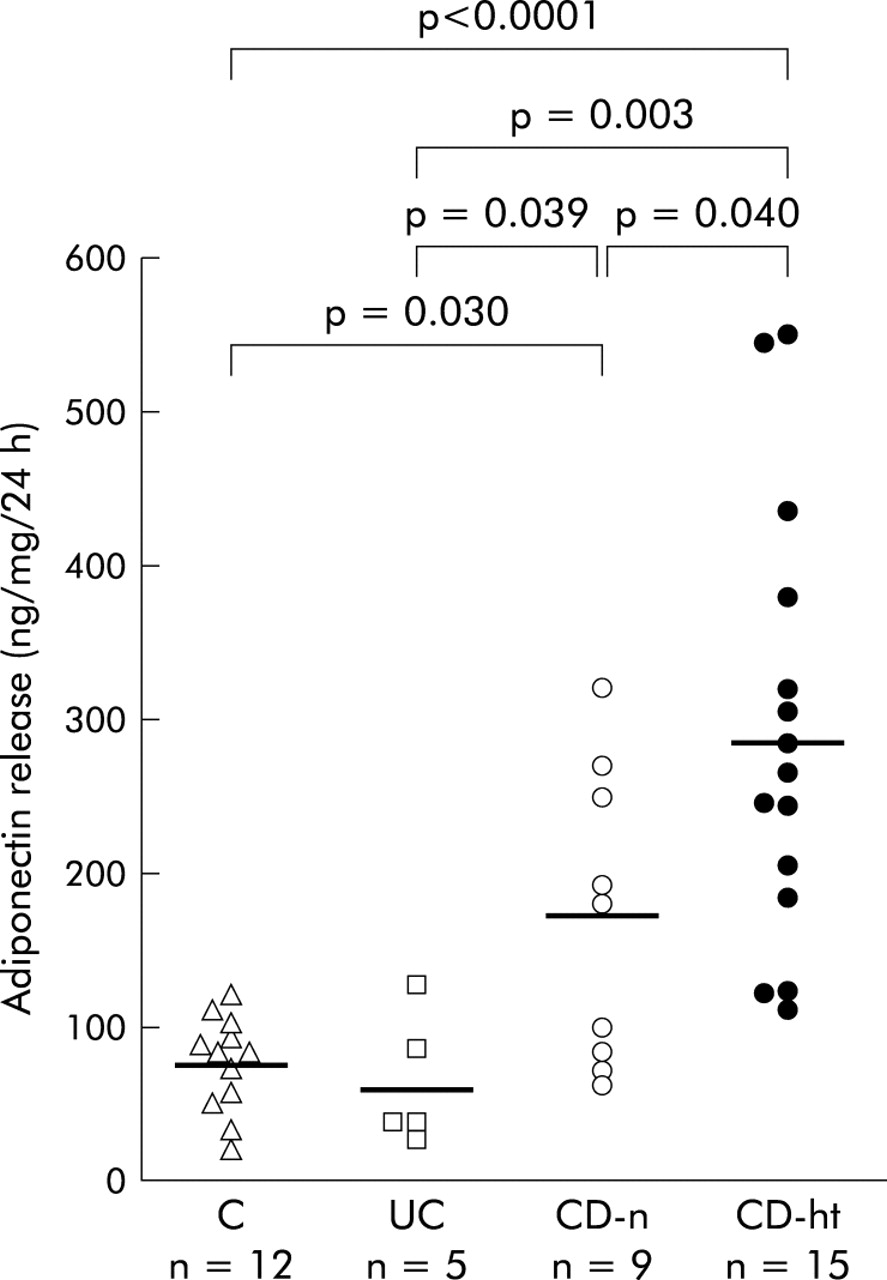
Adiponectin release from the mesenteric adipose tissue in short term culture. Release of adiponectin from adipose tissue was determined by enzyme linked immunosorbent assay and values expressed in ng of adiponectin per ml of conditioned media. Data are individual and mean values (horizontal bars) of adiponectin release from normal mesenteric adipose tissues of controls (C), ulcerative colitis patients (UC), Crohn’s disease patients (CD-n), and hypertrophied mesenteric adipose tissues of CD patients (CD-ht).
Adiponectin mRNA expression in the mesenteric adipose tissue of CD patients
Quantitative real time reverse transcription (RT)-PCR showed a significantly higher expression level of adiponectin mRNA in hypertrophied mesenteric adipose tissue of CD patients (15.1×10−2 (2.1×10−2) cDNA molecules) compared with that in paired normal mesenteric adipose tissue (8.9×10−2 (1.1×10−2) cDNA molecules) (p = 0.024) (fig 5).

Quantitative real time reverse transcription-polymerase chain reaction analysis of adiponectin mRNA expression in mesenteric adipose tissue. Data are individual and mean values (horizontal bars) of adiponectin cDNA molecules per molecule of β-actin cDNA in normal mesenteric adipose tissues of Crohn’s disease patients (CD-n) and hypertrophied mesenteric adipose tissues of CD patients (CD-ht) from the same patients.
Correlation between adiponectin concentrations in hypertrophied mesenteric adipose tissue and laboratory data of CD patients
We next evaluated the relationship between adiponectin concentrations in hypertrophied mesenteric adipose tissue and laboratory data measured just prior to surgery in CD patients. There was no significant correlation between adiponectin concentrations in hypertrophied mesenteric adipose tissues and leucocyte count, haemoglobin level, or platelet count in CD patients (r = −0.13, p = 0.558; r = −0.15, p = 0.507; r = −0.18, p = 0.434, respectively) (fig 6A–C). On the other hand, adiponectin concentrations in hypertrophied mesenteric adipose tissues of CD patients correlated inversely with serum C reactive protein (CRP) levels (r = −0.51, p = 0.015) (fig 6D). There was no significant correlation between tissue adiponectin concentrations and serum CRP levels in control subjects or UC patients (r = −0.32, p = 0.097 and r = −0.37, p = 0.369, respectively).

Correlation between adiponectin concentrations in hypertrophied mesenteric adipose tissue of Crohn’s disease patients and leucocyte count (A), haemoglobin level (B), platelet count (C), and serum C reactive protein (CRP) levels (D). These laboratory data were determined just prior to surgery.
Relationship between adiponectin and IL-6 concentrations in hypertrophied mesenteric adipose tissue of CD patients
Based on the above finding, we evaluated the relationship between adiponectin and the proinflammatory cytokine IL-6, a major inducer of CRP. The concentration of IL-6 in hypertrophied mesenteric adipose tissues of CD patients (90.3 (19.1) pg/mg) tended to be higher than that in the mesenteric adipose tissues of control subjects (54.4 (7.6) pg/mg) but the difference was not statistically significant (p = 0.063) (fig 7A). In hypertrophied mesenteric adipose tissue of CD patients, IL-6 concentrations correlated inversely with adiponectin concentrations (r = −0.43, p = 0.045) (fig 7B).

(A) Interleukin 6 (IL-6) concentrations in the mesenteric adipose tissue of controls (C) and hypertrophied mesenteric adipose tissue of Crohn’s disease patients (CD-ht). IL-6 concentrations were determined by enzyme linked immunosorbent assay and values expressed in pg of IL-6 per mg of total homogenate protein. (B) Correlation between adiponectin and IL-6 concentrations in hypertrophied mesenteric adipose tissue of CD patients.
Adiponectin concentrations in the mesenteric adipose tissue of CD patients with or without internal fistula
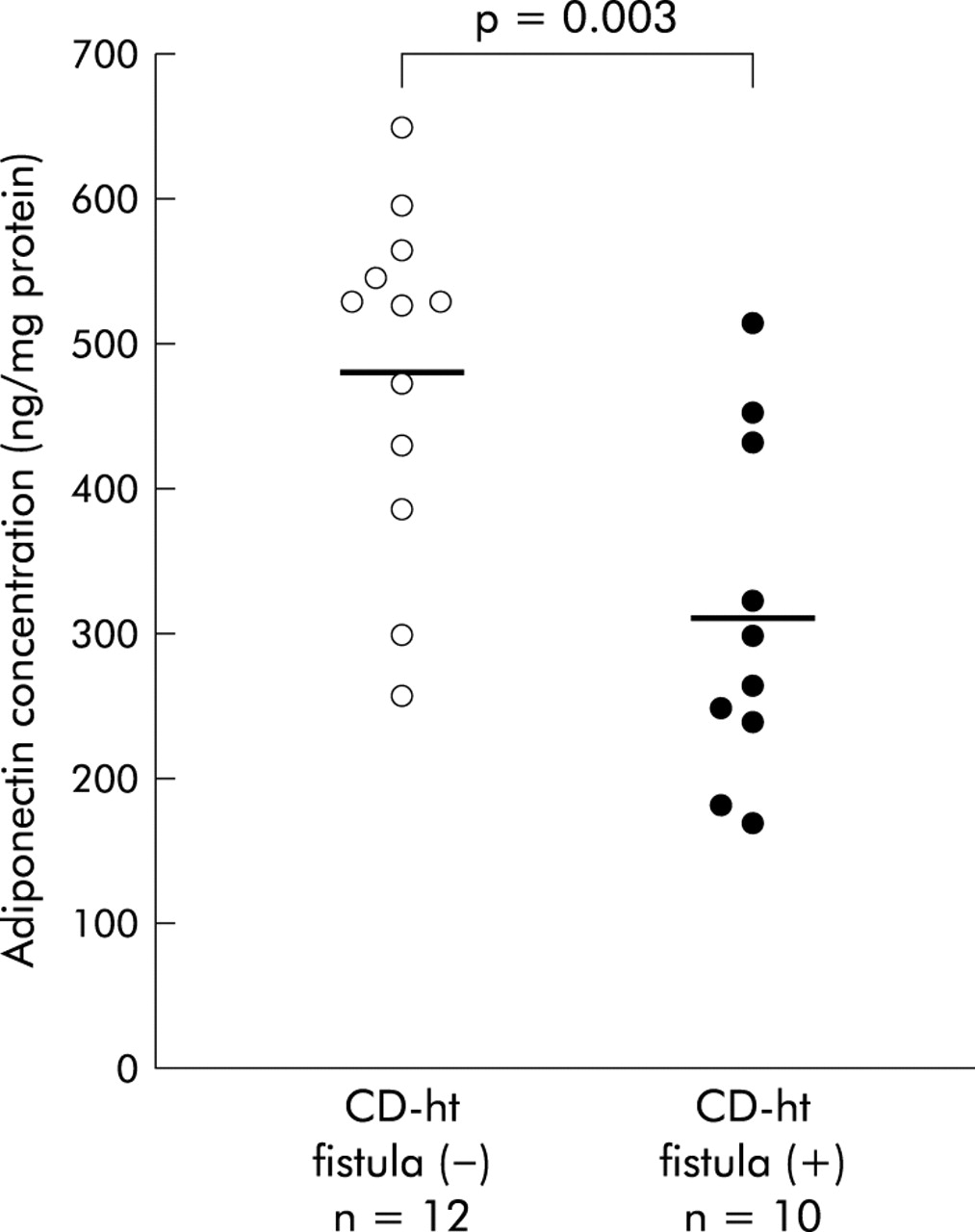
To investigate the association of adiponectin production in mesenteric adipose tissue with the development of an internal fistula, we examined adiponectin concentrations in the mesenteric adipose tissues of CD patients with an internal fistula (eight men, two women; mean age 32.1 (1.9) years (range 24–43)) and without an internal fistula (eight men, four women; mean age 36.3 (2.9) years (range 25–56)). These two groups were matched for age, BMI (18.4 (0.8) v 19.0 (0.4) kg/m2), and disease duration (5.8 (1.7) v 7.5 (1.4) years). Adiponectin concentrations in hypertrophied mesenteric adipose tissues of CD patients with a fistula (312.4 (37.3) ng/mg) were significantly lower than those without a fistula (481.6 (34.2) ng/mg, p = 0.003) (fig 8). Furthermore, adiponectin concentrations in normal mesenteric adipose tissues of CD patients with a fistula (234.5 (28.8) ng/mg) also tended to be lower than those without a fistula (264.0 (52.4) ng/mg) although the difference was not statistically significant (p = 0.629).

Adiponectin concentrations in hypertrophied mesenteric adipose tissue of Crohn’s disease (CD) patients with or without internal fistula. Data are individual and mean values (horizontal bars) of adiponectin concentrations in hypertrophied mesenteric adipose tissues of CD patients without an internal fistula (CD-ht fistula (−)) and those of CD patients with an internal fistula (CD-ht fistula (+)).
Correlation between BMI and adiponectin concentrations or its release
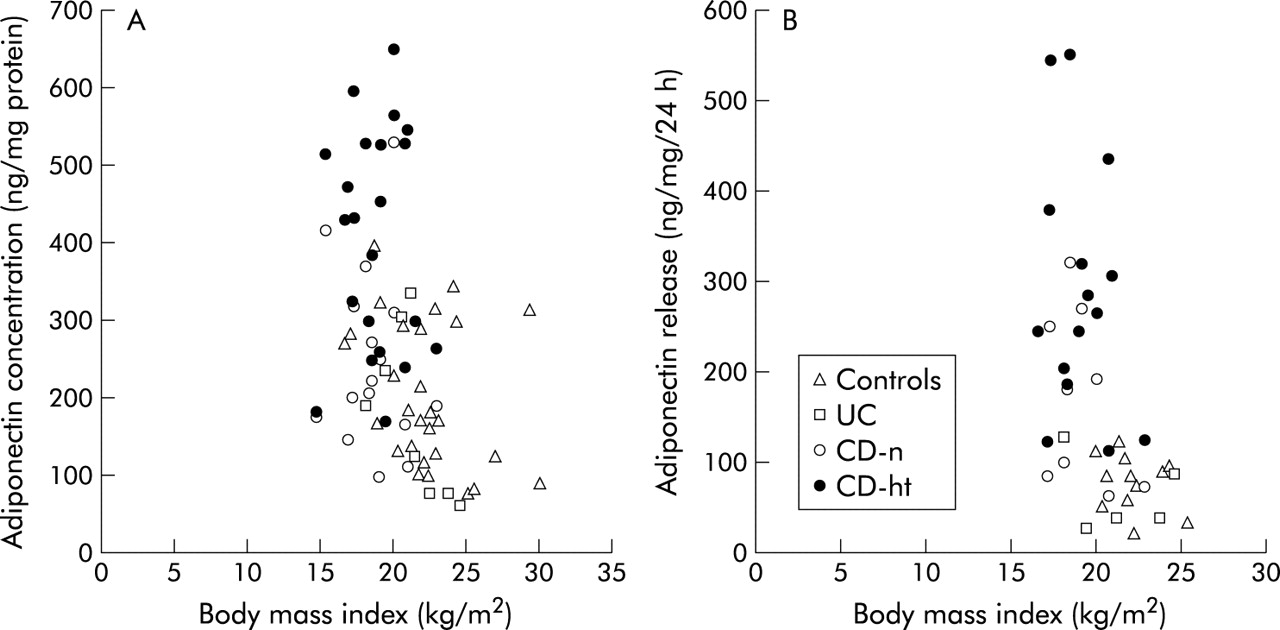
Finally, we evaluated the correlation between BMI and adiponectin concentrations or its release. Adiponectin concentrations and the amount released in all specimens correlated inversely with BMI (r = −0.40, p<0.001 and r = −0.52, p<0.001, respectively) (fig 9A, B). However, there was no significant correlation between BMI and adiponectin concentration or its release in each subgroup; normal mesenteric adipose tissue of controls (r = −0.32, p = 0.099 and r = −0.31, p = 0.328, respectively), UC patients (r = −0.62, p = 0.102 and r = −0.25, p = 0.682, respectively), and CD patients (r = −0.13, p = 0.619 and r = −0.38, p = 0.318, respectively) and hypertrophied mesenteric adipose tissue of CD patients (r = −0.02, p = 0.934 and r = −0.25, p = 0.378, respectively).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Relationship between body mass index and tissue concentrations (A) or release (B) of adiponectin in controls, ulcerative colitis patients (UC), Crohn’s disease patients (CD-n), and hypertrophied mesenteric adipose tissue of CD patients (CD-ht).
DISCUSSION
The presence of hypertrophied mesenteric adipose tissue is one of the characteristic features of CD.1 This change is important in terms of surgery for CD, and its recognition is crucial in determining the extent of resection and, from another point of view, in differentiating CD from other bowel diseases.4 Although the mesenteric fat hypertrophy in CD has long been recognised, its significance remains unknown. Adiponectin, an adipocyte specific secretory protein, was recently shown to play a role in the modulation of a wide array of biological functions. Recent studies have shown that adiponectin has anti-inflammatory effects, especially in endothelial cells and macrophages.7,20–24 Thus its secretion by mesenteric adipocytes could negatively regulate some of the inflammatory reactions observed in CD. The present study demonstrated for the first time that adiponectin production is enhanced in hypertrophied mesenteric adipose tissue contiguous with the involved intestine of CD patients.
Only a few histopathological studies of the mesenteric adipose tissue in CD have been reported. Fibrosis, perivascular inflammation, and intimal and medial thickening of vessels have been observed in hypertrophied mesenteric adipose tissue adjacent to the involved intestine of CD patients.26,27 In the present study, two additional histopathological features of hypertrophied mesenteric adipose tissue of CD patients were defined: significant infiltration of inflammatory cells and reduced size of adipocytes. Infiltrated cells were mainly CD68 positive and CD3 positive T lymphocytes. Massive infiltration of CD68 positive and CD3 positive cells suggests that the inflamed mucosa and its adjacent mesentery share a common inflammation in CD. These cells may disturb mesenteric functions, including blood and lymphatic flow, and hence contribute to mucosal damage. They may also interact with mesenteric adipocytes and induce hyperplasia. The morphological change in adipocytes is more interesting. As small adipocytes are capable of producing larger amounts of adiponectin compared with large adipocytes,28 the reduced size of adipocytes in hypertrophied mesenteric adipose tissue of CD patients is consistent with the increase in adiponectin production reported herein. Desreumaux and colleagues29 found significantly high expression levels of peroxisome proliferator activated receptor γ (PPARγ) mRNA in hypertrophied mesenteric adipose tissue of CD patients compared with normal mesenteric adipose tissue of CD patients and control subjects. PPARγ is a member of the nuclear hormone receptor superfamily, which is predominantly expressed in adipocytes.30,31 PPARγ is a key regulator of adipogenesis. As activation of PPARγ leads to an increase in small adipocytes and upregulates their expression of adiponectin,28,32–34 our results may be relevant to overexpression of PPARγ. Infiltrated cells are a possible source of PPARγ ligands such as 15-deoxy-Δ12–14-prostaglandin J2.35–37
A negative relationship between BMI and plasma adiponectin levels has been reported.38 In the present study, adiponectin concentrations and its release in all specimens of mesenteric adipose tissues correlated inversely with BMI. The high adiponectin concentration and release in CD patients may be partly explained by the lower BMI compared with controls and UC patients. However, in the paired samples of CD patients, adiponectin concentrations and mRNA levels in hypertrophied mesenteric adipose tissues were significantly higher than in normal mesenteric adipose tissues. Furthermore, adiponectin concentration and release in hypertrophied mesenteric adipose tissue of CD patients were shifted upwards in the scattered plot diagrams with BMI compared with the other subgroups (fig 9A, B). These results suggest that increased adiponectin production in hypertrophied mesenteric adipose tissue of CD patients is not only attributed to the lower BMI but also to other factors, including PPARγ ligands, as discussed above.
In the present study, adiponectin concentrations in hypertrophied mesenteric adipose tissue of CD patients correlated negatively with serum CRP levels. However, this inverse correlation was not significant in UC patients and controls. While the exact link between tissue adiponectin concentration and serum CRP level is not clear at present, this difference may be relevant to the characteristic involvement of mesenteric adipose tissue in the pathogenesis of CD. On the other hand, our study also demonstrated a negative correlation between adiponectin and IL-6 production in hypertrophied mesenteric adipose tissue of CD patients. This finding suggests that adiponectin inhibits inflammatory cytokine production in mesenteric adipose tissue. As IL-6 is a major inducer of CRP,39,40 it may be the missing link between adiponectin and CRP.
In CD, internal fistulas develop almost exclusively at the mesenteric border. Our results showed significantly low adiponectin concentrations in hypertrophied mesenteric adipose tissues of CD patients with an internal fistula compared with those free of this complication. Such a decrease in adiponectin production may worsen the inflammation and increase the risk of internal fistula formation. As anti-TNF-α therapy (infliximab) is effective in the treatment of internal fistulas in CD, this result may have relevance to the anti-TNF-α effects of adiponectin.7,12,21 The exact mechanisms that cause reduction of adiponectin concentrations in hypertrophied mesenteric adipose tissue of CD patients with a fistula remain elusive and further studies are needed to identify these mechanisms.
Adipocytes produce several adipocytokines, including proinflammatory cytokines. It has been reported that TNF-α mRNA levels in hypertrophied mesenteric adipose tissue of CD patients tended to be higher than in normal mesenteric adipose tissue of CD patients but the difference between these locations was not statistically significant.29 While mesenteric adipocytes may produce both TNF-α and adiponectin, our results suggest that the latter is a significant adipocytokine derived from mesenteric adipocytes stimulated by transmural inflammation as a result of CD. Barbier and colleagues41 recently reported high mRNA levels of leptin in the mesenteric adipose tissue of patients with CD and UC. However, leptin levels in hypertrophied mesenteric adipose tissue of CD patients were lower than those in normal mesenteric adipose tissue of CD patients, and similar to those in the mesenteric adipose tissue of UC patients.41 Considered together, it is conceivable that the mechanisms that regulate leptin production are different to those of adiponectin and TNF-α.
In conclusion, we have demonstrated in the present study that adipocytes in hypertrophied mesenteric adipose tissue of CD patients produce and secrete significant amounts of adiponectin. Mesenteric adipocytes may act, not only as energy storage cells, but also as immunoregulating cells in the case of intestinal inflammation via adiponectin production. The hypertrophied mesenteric adipose tissue of CD patients could serve as a barrier that prevents the spread of inflammation into the intra-abdominal space. Dysregulation of adiponectin production in mesenteric adipose tissue may play a role in the pathogenesis of intestinal disorders, including CD.
Acknowledgments
We gratefully acknowledge the technical assistance of Sachiyo Tanaka and Fumie Katsube.
REFERENCES
Footnotes
-
Conflict of interest: None declared.