Article Text

Abstract
Background and aims: Epithelium derived interleukin (IL)-15 signalling via IL-15Rα is critical for the development, activation, and survival of intraepithelial lymphocytes (IEL). We aimed to better understand the IL-15 driven effects on IEL underlying mucosal damage and lymphomagenesis in coeliac disease (CD).
Methods: Enterocytes, IEL, and lamina propria mononuclear cells (LPMC) were isolated from 46 patients with uncomplicated CD (25 untreated and 21 treated) and 22 controls. IL-15 and IL-15Rα expression were determined by immunoblotting. Secretion of IL-15, interferon γ (IFN-γ), tumour necrosis factor α (TNF-α), and granzyme B into cell culture supernatants was assessed by ELISA. The ability of IL-15 to regulate IEL proliferation, perforin/granzyme dependent cytotoxicity, and apoptosis was tested by adding different combinations of IL-15, IL-15 blocking antibody, or chloroquine to IEL cultured alone or with Caco-2 cells as target. IL-15 mucosal levels were also determined by ELISA in five patients with complicated CD (two ulcerative jejunoileites, one refractory sprue, and two enteropathy associated T cell lymphomas) tested for T cell receptor γ chain clonality.
Results: IL-15 was overexpressed in untreated CD enterocytes and LPMC, and in the mucosa of complicated CD patients and uncomplicated untreated CD patients, where its levels correlated with the degree of mucosal damage. Enterocytes from untreated, but not treated, CD patients and controls secreted IL-15. Untreated CD IEL, characterised by higher IL-15Rα expression, showed increased proliferation, production of IFN-γ and TNF-α, and perforin/granzyme dependent cytotoxicity, and a decreased propensity to apoptosis in response to IL-15.
Conclusions: Our findings suggest that IL-15 plays a crucial role in the generation of epithelial damage in active CD. Its promotion of IEL survival in CD may predispose to the emergence of T cell clonal proliferations. Blocking IL-15, by suppressing uncontrolled IEL activation and survival, has the potential to provide new therapeutic tools to prevent tissue damage and lymphomagenesis in CD.
- CD, coeliac disease
- EDTA, ethylenediamine-tetraacetic acid
- FITC, fluorescein isothiocyanate
- IFN, interferon
- IL, interleukin
- IEL, intraepithelial lymphocyte
- LPMC, lamina propria mononuclear cell
- LPS, lipopolysaccharide
- PI, propidium iodide
- TCR-γ, T cell receptor γ chain
- TNF, tumour necrosis factor
- apoptosis
- coeliac disease
- cytotoxicity
- enterocyte
- interleukin 15
- intraepithelial lymphocyte
- T cell lymphoma
Statistics from Altmetric.com
- CD, coeliac disease
- EDTA, ethylenediamine-tetraacetic acid
- FITC, fluorescein isothiocyanate
- IFN, interferon
- IL, interleukin
- IEL, intraepithelial lymphocyte
- LPMC, lamina propria mononuclear cell
- LPS, lipopolysaccharide
- PI, propidium iodide
- TCR-γ, T cell receptor γ chain
- TNF, tumour necrosis factor
- apoptosis
- coeliac disease
- cytotoxicity
- enterocyte
- interleukin 15
- intraepithelial lymphocyte
- T cell lymphoma
The discovery that interleukin (IL)-15 may be produced by intestinal epithelial cells1 and is a potent stimulant of intraepithelial lymphocytes (IEL)2 focused the attention in mucosal immunity on the role of IL-15 in the interplay between enterocytes and IEL.3,4 IL-15, a 14 kDa glycoprotein mainly produced by macrophages as well as non-lymphoid cells, is a key regulatory cytokine which supports the homeostasis between innate and adaptive immunity.5 The occurrence of lymphocytopenia and specific lack of natural killer cells, natural killer T cells, CD8+ T cells, and γ/δ IEL in IL-15 deficient (IL-15−/−) and IL-15Rα deficient (IL-15Rα−/−) transgenic mice6,7 suggest that IL-15 signals transmitted via IL-15Rα are critical for the development, activation, and/or survival of these cells.8
As IL-15 expression is strictly regulated at multiple distinct levels, including transcription, translation, and intracellular trafficking,5 removal of these negative control mechanisms results in increased IL-15 production, which may predispose to the risk of excessive autoreactive T cell survival and abnormal lymphocyte activation, thus leading to the development of autoimmune or chronic inflammatory diseases.9,10 Indeed, IL-15 overexpression is associated with an array of immune mediated intestinal disorders, such as inflammatory bowel disease11,12 and coeliac disease (CD).13–16 In this latter condition, IL-15 has been shown to be involved in the generation of villous atrophy,13,14 in favouring the selective expansion of CD94+ IEL,15 and in promoting the emergence of T cell clonal proliferations in refractory sprue.16
In the present study, we aimed to clarify the role of IL-15 in modulating the interactions between enterocytes and IEL in CD.
PATIENTS AND METHODS
Patients and tissues
Size appropriate and well oriented endoscopic biopsy specimens were obtained from the second part of the duodenum in 25 uncomplicated untreated CD patients (mean age 39.2 years; range 20–65). The histopathological diagnosis was based on typical mucosal lesions with crypt cell hyperplasia, villous atrophy, and increased number of IEL. All untreated CD patients tested positive for antiendomysial antibodies at the time of diagnosis. Biopsies from 18 of them showed a Marsh IIIc lesion (total villous atrophy), while biopsies from the remaining seven patients showed a Marsh IIIb lesion (subtotal villous atrophy). In all of them there was histological improvement of duodenal mucosa following gluten withdrawal. Biopsies were also collected from 21 uncomplicated CD patients on a gluten free diet for at least 12 months (mean age 41.5 years; range 21–63) who were in clinical and histological remission, and negative for antiendomysial antibodies. Twenty two subjects undergoing upper gastrointestinal endoscopy for functional dyspepsia (mean age 43.2 years; range 21–68), who tested negative for antiendomysial antibody and had normal IgA levels and normal histology, were also studied. Some of the biopsy samples were processed according to standard methods for routine histology while others were homogenised for determination of IL-15 or used to obtain suspensions of purified enterocytes, IEL, and lamina propria mononuclear cells (LPMC). IL-15 was also determined in tissue samples from the small intestine of five patients with complicated CD (mean age 49.0 years; range 34–66). Two of these patients were affected by ulcerative jejunoileitis, one by refractory sprue, and two by an enteropathy associated T cell lymphoma. In the mucosal samples of complicated CD patients, the clonality of the T cell receptor γ chain (TCR-γ) gene was investigated by polymerase chain reaction amplification of TCR-γ using an adapted protocol with five different primers in two reactions, as described by McCarthy and colleagues.17 Clinical and pathological features of complicated CD patients are reported in table 1. Each subject gave informed consent to the study.
Clinical and pathological features of complicated coeliac patients
Cell isolation
Enterocytes, IEL, and LPMC were isolated as previously described.18 Briefly, the epithelial layer was removed with 1 mM ethylenediamine-tetraacetic acid (EDTA; Sigma, St Louis, Missouri, USA) and 1 mM dithiothreitol (Sigma). After stirring for one hour at 37°C, the single cell suspension was pelleted from the supernatant and IEL and enterocytes were then separated on a Percoll density gradient (Pharmacia, Uppsala, Sweden). A discontinuous density gradient (25%, 40%, and 75%) was used. Cells obtained between the 40% and 75% fractions were collected as IEL whereas cells that were found between the 40% and 25% interface were collected as enterocytes, respectively. To release LPMC, the remaining tissue was treated with type 1A collagenase 128 U/ml (Sigma) for two hours with stirring at 37°C. The crude cell suspension was allowed to stand for five minutes to permit sedimentation of debris. Cells from the supernatant were washed twice with RPMI-1640 medium containing 10% fetal calf serum, 100 U/ml penicillin, and 100 μg/ml streptomycin. Single cell suspensions were resuspended in 1 ml of phosphate buffered saline and kept on ice until used. Cells were not used if viability did not exceed 90%. The purity of enterocyte and IEL populations, determined by staining cytocentrifuge preparations with anticytokeratin pan antibody (Boehringer Mannheim, Indianapolis, Indiana, USA) at a dilution of 1:400, was found to exceed 93% and 90%, respectively. The purity of the LPMC population was 90%, as assessed by flow cytometric analysis with fluorescein isothiocyanate (FITC) conjugated mouse antihuman CD45 monoclonal antibody and its isotype control (FITC conjugated IgG1) (Becton Dickinson, San Jose, California, USA).
Cell culture
IL-15 was measured by ELISA in the supernatants of freshly isolated enterocytes (1×105/ml) cultured for 24 hours, and in the supernatants of freshly isolated LPMC (1×105/ml) cultured for 24 hours in the presence or absence of either 5 μg/ml lipopolysaccharide (LPS) or 1000 U/ml interferon (IFN)-γ (R&D Systems, Minneapolis, Minnesota, USA). IFN-γ and tumour necrosis factor (TNF)-α were measured by ELISA in the supernatants of IEL (1×105/ml) cultured for 24 hours in the presence or absence of recombinant human IL-15 (Chemicon, Temecula, California, USA) at a concentration of 50 ng/ml, which has been shown to be optimal for lymphocyte activation.3 Flow cytometric FITC-annexin V and propidium iodide (PI) staining was used to quantify apoptosis in IEL incubated for 18 hours in the presence or absence of IL-15 (1, 10, and 50 ng/ml), 10 μg/ml IL-15 blocking antibody (R&D Systems), or an isotype matched control antibody (mouse IgG1). Granzyme B concentrations in the supernatants of IEL cultured in the presence or absence of 50 ng/ml IL-15, 10 μg/ml IL-15 blocking antibody, or control IgG1 were determined by ELISA.
Proliferation assay
For the proliferation assay, 1×105 IEL were plated out in 96 well microtitre plates in a total volume of 0.2 ml per well in the presence of IL-15 (1, 10, or 100 ng/ml). After 72 hours of culture, 1 μCi [3H] thymidine was added to each well for the last eight hours to determine DNA synthesis. IEL lysates were collected on a glass filter and radioactivity was measured. Values were calculated as stimulation index of mean cpm in the presence of IL-15 divided by the mean cpm of unstimulated cultures.
Cytotoxicity assay
A 51Cr release assay was performed using cells of the human enterocyte-like cell line Caco-2 as target. Caco-2 cells were labelled with 30 μCi of 51Cr (Amersham, Arlington Heights, Illinois, USA) for one hour and then cocultured with IEL at different effector target ratios (0.6, 1.2, 2.5, and 5) for four hours in the presence or absence of 50 ng/ml IL-15, IL-15 blocking antibody (1, 5, and 10 μg/ml), or control IgG1. In parallel experiments, IEL preincubated for one hour with different concentrations (0, 10, and 50 μg/ml) of chloroquine (Sigma), an inhibitor of granzyme/perforin dependent cytotoxicity,19 were cocultured with Caco-2 cells at the effector target ratio of 5. After incubation, 100 μl of medium from each well of labelled cocultures were carefully removed and radioactivity measured in a gamma counter. The percentage of specific 51Cr release was calculated according to the following formula:
100 × (experimental release − spontaneous release) / (total content − spontaneous release),
where total content was determined by lysing target cells with 1 N HCl.
Evaluation of apoptosis by FITC-annexin V and PI binding
Apoptosis was quantified using FITC-annexin V and PI according to the manufacturer’s protocol (ApoDETECT FITC-Annexin V Kit; Zymed Laboratories, San Francisco, California, USA). Cells were stained with 5 μl of FITC-annexin V diluted 1:10 in buffer, and 2.5 μl of PI. After incubation for 15 minutes, cells were analysed by flow cytometry with standard FACScan equipment (Becton Dickinson). The following controls were used to set up compensation and quadrants: unstained cells, cells stained with FITC-annexin V alone, and cells stained with PI alone. Cells undergoing the early stage of apoptosis were stained with FITC-annexin V alone (% of apoptosis). Cells permeabilised with 0.1% saponin were used as the control for PI positive cells.
ELISA
IL-15, IFN-γ, TNF-α, and granzyme B levels were measured by ELISA using, respectively, the Quantikine human IL-15 immunoassay (R&D Systems), the human IFN-γ ELISA kit (Chemicon), the human TNF-α ELISA kit (Chemicon), and the Pelikine compact human granzyme B ELISA kit (CLB, Amsterdam, the Netherlands) according to the manufacturers’ instructions. For determination of IL-15 concentrations in the mucosa, tissue samples were added to 0.3 ml of Tris HCl 20 mmol/l, pH 7.4, containing 1 μmol/l phenylmethylsulfonyl fluoride, 10 μmol/l leupeptin, 10 μmol/l pepstatin, and 1 μmol/l aprotinin, and homogenised using a handheld Potter-Elvehjem tissue grinder and by sonication (5 W, 80% output, 1 minute and 50 seconds, alternating 10 seconds sonication and 10 seconds pause) with a sonicator (Vibracell; Sonic and Materials, Danbury, Connecticut, USA).
Immunoblotting
Cell lysates were prepared by lysing cells in ice cold lysing buffer containing 20 mM Tris HCl, pH 7.6, 150 mM sodium chloride, 1 mM phenylmethylsulfonyl fluoride, 1% Triton-X, 1 mM EDTA, 2 mM sodium orthovanadate, 10 μg/ml pepstatin, 10 μg/ml leupeptin, and 10 μg/ml aprotinin (Sigma). Protein concentrations in cell lysates were determined using the Pierce protein assay (Pierce, Rockford, Illinois, USA). Equivalent amounts of protein (12 μg) were separated on 10–15% sodium dodecyl sulphate-polyacrylamide gels and transferred to nitrocellulose membranes (Bio-Rad Laboratories, Hercules, California, USA). Membranes were blocked with 5% non-fat milk, followed by incubation for 90 minutes at room temperature with mouse anti-IL-15 or anti-IL-15Rα antibodies (both from R&D Systems). Membranes were washed several times with TTBS and then incubated for 90 minutes at room temperature with appropriate horseradish peroxidase conjugated secondary antibodies (Santa Cruz Biotechnology, Santa Cruz, California, USA). After extensive washing, the reaction was developed by enhanced chemiluminescent staining (Super Signal; Pierce). Each blot was stripped and analysed for β-actin, as an internal loading control, using an antihuman β-actin antibody (Santa Cruz Biotechnology). Bands were quantified by scanning densitometry using an LKB Ultrascan XL Laser Densitometer (Kodak Ltd, Hemel Hempstead, UK).
Mucosal morphometry
In order to quantify the degree of mucosal flattening in CD, the surface area to volume ratio was estimated in biopsy specimens using a Weibel graticule (Graticules Ltd, Tunbridge Wells, UK) inserted into the microscope eyepiece, according to the method of Dunnil and Whitehead,20 as standardised by us.21 A record was made of the number of times the lines cut the mucosal surface (c) and the number of hits (h)—that is, end points of the lines falling on the lamina propria. At a constant magnification (×125), the length (l) of each line of the graticule was 1.7×10−2 cm. The ratio c/lh was then calculated and regarded as a measure of surface area to volume ratio.
Statistical analysis
Data were analysed using the GraphPad Prism statistical PC program (GraphPad Software, San Diego, California, USA) by means of the non-parametric Mann-Whitney U test. One way analysis of variance (ANOVA test) was used to define differences in the percentage of cytotoxic or apoptotic cells obtained in cultures treated with various concentrations of stimuli. A level of p<0.05 was considered statistically significant.
RESULTS
IL-15 production by enterocytes
As shown in fig 1A, IL-15 protein was significantly (p<0.0001) higher in enterocytes from untreated CD patients in comparison with treated CD patients and controls. No significant difference was found between treated CD patients and controls. Figure 1B shows that IL-15 was undetectable in the supernatants of enterocytes from treated CD patients and controls, but it was secreted by enterocytes from untreated CD patients (mean 19.3 (4.6) pg/ml).

Interleukin 15 (IL-15) production by enterocytes. (A) Expression of IL-15 and β-actin in enterocytes isolated from an untreated patient with coeliac disease (CD), a treated CD patient, and a control subject. The example is representative of experiments performed in 10 subjects for each group. Bottom panel indicates densitometry of western blots. Horizontal bars represent median values (*p<0.0001 v treated CD patients and controls). (B) IL-15 release by enterocytes from 10 untreated CD patients, nine treated CD patients, and 10 controls. Freshly isolated enterocytes were cultured for 24 hours and IL-15 was measured by ELISA in the enterocyte supernatants. Results are mean (SD).
IL-15Rα expression by IEL
We investigated by immunoblotting whether IEL expressed IL-15Rα required for IL-15 triggered signal transduction.22 Figure 2A shows significantly (p<0.001) lower IL-15Rα expression in IEL isolated from controls in comparison with untreated and treated CD patients, without any significant difference between untreated and treated CD patients.

Interleukin 15 receptor α (IL-15Rα) expression by intraepithelial lymphocytes (IEL) and IL-15 driven Th1 cytokine production and proliferation of IEL. (A) Expression of IL-15Rα and β-actin in IEL isolated from an untreated patient with coeliac disease (CD), a treated CD patient, and a control subject. The example is representative of experiments performed in 10 subjects for each group. Lower panel shows densitometry of western blots. Horizontal bars represent median values (*p<0.0001 v untreated and treated CD patients). (B) Interferon (IFN)-γ and tumour necrosis factor (TNF)-α secretion by IEL from nine untreated CD patients, eight treated CD patients, and eight controls after IL-15 stimulation. Freshly isolated IEL were cultured for 24 hours in the absence or presence of 50 ng/ml IL-15. IFN-γ and TNF-α levels were measured by ELISA in the cell culture supernatants. Results are mean (SD). (C) Proliferation of IEL in response to IL-15. IEL from eight untreated CD patients, seven treated CD patients, and seven controls were cultured in the presence of different concentrations of IL-15 (1, 10, and 100 ng/ml) for 72 hours. Cultures were then pulsed with 1 μCi [3H] thymidine for the last eight hours and [3H] thymidine incorporation was measured. Data are expressed as stimulation index. Results are mean (SD).
Effect of IL-15 on secretion of IFN-γ and TNF-α by IEL
To examine the role of IL-15 in regulating Th1 cytokine production by IEL, IFN-γ and TNF-α were measured in the supernatants of both unstimulated and IL-15-stimulated IEL. As shown in fig 2B, IL-15 stimulation significantly (p<0.001) enhanced IFN-γ and TNF-α secretion by IEL in untreated CD patients (from mean 88.4 (10.7) to 758.1 (122.7) and from 12.8 (5.6) to 121.7 (34.2) pg/ml, respectively), in treated CD patients (from mean 10.4 (1.6) to 98.3 (36.2) and from 0 to 22.4 (7.3) pg/ml, respectively), and in controls (from mean 0 to 79.4 (22.8) and from 0 to 18.6 (6.9) pg/ml, respectively). In the supernatants of IL-15 stimulated IEL, IFN-γ and TNF-α levels were significantly (p<0.001) higher in untreated CD patients in comparison with treated CD patients and controls while no significant difference was found between treated CD patients and controls.
Proliferative responses of IEL to IL-15
As IL-15 induces IEL proliferation,2 the proliferation inducing effects of IL-15 on IEL from untreated and treated CD patients and controls were compared. As shown in fig 2C, in untreated CD patients the proliferative stimulation index was raised significantly (p<0.0001) by incubation with increasing concentrations (1, 10, and 100 ng/ml) of IL-15. As for treated CD patients and controls, the proliferative stimulation index was increased by IL-15, but only after 100 ng/ml IL-15 did this difference reach a statistically significant level (p<0.001). At each IL-15 concentration, the proliferative stimulation index of IEL from untreated CD patients was significantly (p<0.001) higher than that of IEL from treated CD patients and controls.
Effect of IL-15 on the cytotoxicity of IEL against epithelial cells
To test whether exogenous IL-15 influenced the killing activity of IEL against epithelial cells, IEL isolated from untreated CD patients and controls were cocultured with Caco-2 cells in the presence or absence of 50 ng/ml IL-15, and a cytotoxicity assay was performed to determine the extent of target cell death (fig 3A, B). Figure 3A shows that control IEL, which had no killing activity in the absence of exogenous IL-15, manifested strong cytotoxicity against Caco-2 cells when cultured with IL-15. Additionally, the cytotoxicity of CD IEL was significantly (p<0.001) greater than that of control IEL, and was increased further by IL-15. At each effector target ratio, the killing activity of IL-15 stimulated IEL from both untreated CD patients and controls was significantly higher (p<0.001) than that observed when IEL were cultured in unstimulated conditions. Figure 3B shows that the cytotoxicity of IL-15 stimulated CD and control IEL and of unstimulated CD IEL was abrogated when cells were preincubated with chloroquine, an inhibitor of granzyme/perforin dependent cytotoxicity.19
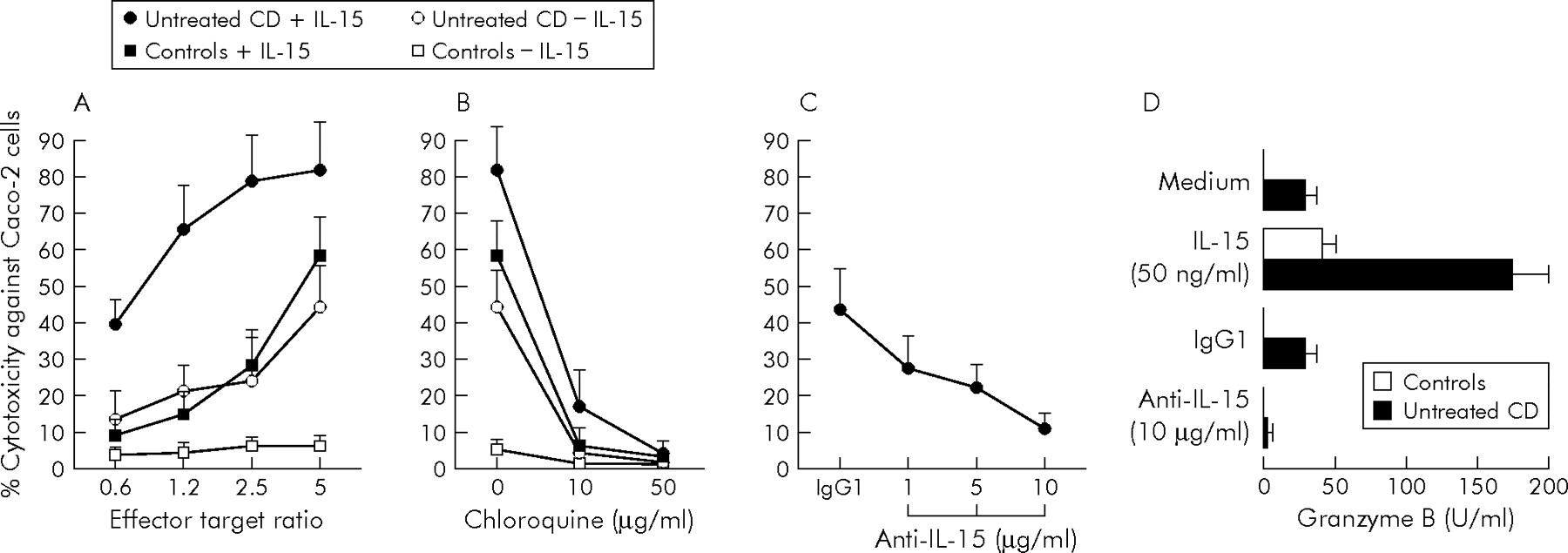
Effects of interleukin 15 (IL-15) on perforin/granzyme dependent cytotoxicity of intraepithelial lymphocytes (IEL). (A) Caco-2 target cells labelled with 51Cr were cocultured with IEL from eight untreated coeliac disease (CD) patients and eight control subjects in the presence or absence of 50 ng/ml IL-15 at various effector target ratios. (B) IEL from seven untreated CD patients and seven controls were pretreated with different concentrations of chloroquine, and then cocultured at an effector target ratio of 5 with Caco-2 target cells in the presence or absence of 50 ng/ml IL-15. (C) IEL from seven untreated CD patients were cocultured at an effector target ratio of 5 with Caco-2 target cells in the presence of increasing concentrations of IL-15 blocking antibody or control IgG1. Cytotoxic activity against Caco-2 target cells was tested by 51Cr release assay. Results are mean (SD). (D) Granzyme B release by IEL from eight untreated CD patients and eight controls cultured for 24 hours in RPMI 1640 medium only, or in the presence of 50 ng/ml IL-15, 10 μg/ml IL-15 blocking antibody, or control IgG1. Granzyme B was measured by ELISA in the cell culture supernatants. Results are mean (SD).
To investigate the role of endogenous IL-15 in modulating IEL cytotoxicity in CD, IEL isolated from untreated CD patients were cocultured with Caco-2 cells after treatment with IL-15 blocking antibody. As shown in fig 3C, cytotoxicity against Caco-2 cells was reduced in a concentration dependent manner when IEL were treated with increasing concentrations of IL-15 blocking antibody.
To further explore the role of perforin/granzyme molecules in the cytotoxic activity of IEL in CD, granzyme B was measured in the supernatants of IEL cultured with IL-15 or IL-15 blocking antibody. Figure 3D shows that control IEL secreted detectable amounts of granzyme B only when stimulated with IL-15 (43.4 (8.8) U/ml) but not when cultured in the presence of RPMI 1640 medium only. In untreated CD patients, IL-15 stimulation significantly (p<0.001) enhanced granzyme B release by IEL (from 32.1 (6.5) to 174.2 (24.1) U/ml). Additionally, neutralisation of IL-15 by culturing IEL from untreated CD patients with IL-15 blocking antibody significantly reduced (p<0.001) release of granzyme B from 30.1 (5.9) to 7.4 (2.1) U/ml.
Effect of IL-15 on apoptosis of IEL
To investigate the role of IL-15 in regulating IEL apoptosis, control IEL were cultured with IL-15, while IEL from untreated CD patients were cultured with IL-15 blocking antibody or control IgG1. Apoptosis was analysed by flow cytometry with FITC-annexin V and PI staining. In fig 4, representative results of control subjects (A, B) showed that, in comparison with unstimulated IEL (fig 4A), IEL cultured in the presence of 50 ng/ml IL-15 (fig 4B) showed a lower percentage of apoptotic cells (FITC-annexin V positive/PI negative). Additionally, representative results of untreated CD patients (fig 4C, D) showed that in comparison with IEL cultured with IgG1 (fig 4C), IEL treated with 10 μg/ml IL-15 blocking antibody (fig 4D) showed a higher percentage of apoptosis. As shown in fig 4E, the percentage of apoptotic control IEL (mean 32.1 (6.7)%) was significantly reduced by incubation with increasing concentrations of 1, 10, and 50 ng/ml IL-15 to 19.3 (4.9)% (p<0.01), 12.8 (3.2)% (p<0.001), and 8.8 (1.9)% (p<0.001), respectively. Consistent with this, blocking IL-15 significantly (p<0.001) enhanced the percentage of apoptosis of CD IEL from 4.2 (1.3)% to 17.7 (3.9)%.

Effect of interleukin 15 (IL-15) on intraepithelial lymphocyte (IEL) apoptosis. IEL isolated from a control subject were cultured in the absence (A) or presence (B) of 50 ng/ml IL-15, while IEL isolated from an untreated coeliac disease (CD) patient were cultured with control IgG1 (C) or 10 μg/ml IL-15 blocking antibody (D). The percentage of apoptotic cells was assayed by flow cytometric analysis of fluorescein isothiocyanate (FITC)-annexin V and propidium iodide (PI) binding. The lower right quadrant represents apoptotic cells (FITC-annexin V positive/PI negative). Numbers within the dot plots represent the percentages of apoptotic cells. Data are representative of experiments performed in nine untreated CD patients and eight controls. (E) Percentage of apoptotic IEL from controls subjects cultured with increasing concentrations of IL-15 and blocking effect of 10 μg/ml IL-15 neutralising antibody on CD IEL apoptosis. Results are mean (SD).
IL-15 production by LPMC
As shown in fig 5A, IL-15 protein was significantly (p<0.001) higher in LPMC from untreated CD patients in comparison with treated CD patients and controls, while no significant difference was found between treated CD patients and controls. Figure 5B shows that only LPMC from untreated CD patients released IL-15 in the absence of stimulation (20.8 (4.7) pg/ml). When LPMC were stimulated with LPS or IFN-γ, significantly (p<0.001) higher IL-15 levels were detected in the supernatants of untreated CD patients (mean 82.4 (15.1) and 68.3 (12.7) pg/ml, respectively) in comparison with treated CD patients (mean 17.8 (4.6) and 13.6 (3.9) pg/ml, respectively) and controls (mean 15.9 (3.7) and 14.1 (2.4) pg/ml, respectively). No significant difference was found between treated CD patients and controls.

Interleukin 15 (IL-15) production by lamina propria mononuclear cells (LPMC). (A) Expression of IL-15 and β-actin in LPMC isolated from an untreated coeliac disease (CD) patient, a treated CD patient, and a control subject. The example is representative of experiments performed in 10 subjects for each group. Bottom panel shows densitometry of western blots. Horizontal bars represent median value (*p<0.001 v treated CD patients and controls). (B) LPMC isolated from 11 untreated CD patients, 10 treated CD patients, and nine controls were cultured for 24 hours in the absence or presence of 5 μg/ml lipopolysaccharide (LPS) or 1000 U/ml interferon γ (IFN-γ). IL-15 was measured by ELISA in cell culture supernatants. Results are mean (SD).
IL-15 expression in the small intestinal mucosa
Figure 6 shows that mucosal IL-15 concentration was significantly higher both in patients with uncomplicated untreated CD (median 11.6 pg/ml (range 11.0–14.1)) and in patients with complicated CD (median 12.6 pg/ml (range 11.3–16.1)) than in patients with uncomplicated treated CD (median 7.7 pg/ml (range 4.0–11.5); p<0.001) and controls (median 6.5 pg/ml (range 3.9–10.6); p<0.001). There was no significant difference between uncomplicated untreated CD patients and complicated CD patients, or between treated CD patients and controls. In the complicated CD group, two patients with enteropathy associated T cell lymphoma showed the highest IL-15 levels. Figure 7 shows that in uncomplicated untreated CD, a significant inverse correlation (rs = −0.85; p<0.005) between IL-15 mucosal levels and surface area to volume ratio was found.

Interleukin 15 (IL-15) levels in the small intestinal mucosa of five patients with complicated coeliac disease (CD), 10 uncomplicated untreated CD patients, 10 uncomplicated treated CD patients, and 10 controls. Among the five patients with complicated CD, two were affected by ulcerative jejunoileitis (triangle), one by refractory sprue (square), and two by an enteropathy associated T cell lymphoma (open circle). IL-15 mucosal concentration was measured by ELISA. Horizontal bars represent median values.

Correlation between the degree of mucosal damage, morphometrically evaluated and expressed as surface area to volume ratio, and interleukin 15 (IL-15) mucosal levels in uncomplicated untreated coeliac disease patients.
DISCUSSION
The results presented in this study suggest the involvement of IL-15 in the pathogenesis of CD. Firstly, we confirmed that IL-15 is overexpressed in active CD mucosa where its level correlated with the degree of mucosal damage according to morphometric analysis, and showed that enterocytes and LPMC represent the major source of this cytokine. Furthermore, we found that IL-15 induces IEL to proliferate and release proinflammatory cytokines, promotes IEL perforin/granzyme dependent cytotoxicity against epithelial cells, and inhibits IEL apoptosis. In addition, we have shown that the concentration of IL-15 in the mucosa is significantly higher in patients with complicated CD and in untreated CD patients in comparison with patients with treated CD and controls.
It has been suggested that macrophages and dendritic cells are the main producers of IL-15.5 However, recently, attention has been focused on intestinal epithelial cells as a source of this cytokine. Immunohistochemical studies have shown conflicting results concerning IL-15 expression by CD epithelium, which has been reported as either devoid of staining or intensely stained from the surface to the bottom of the crypts.13,16 Our immunoblotting analysis on isolated enterocytes showed an increase in IL-15 protein in untreated CD in comparison with controls. In keeping with previous flow cytometric findings,16 enterocyte expression of IL-15 returned to normal levels in treated CD. Interestingly, although IL-15 is reported to be a poorly secreted cytokine, hardly detectable by ELISA,23 we were able to measure it in the supernatants of enterocytes from untreated CD patients cultured for 24 hours. These findings contrast with those of Mention and colleagues,16 who reported that IL-15, rather than being released, is retained in functional form on the surface of epithelial cells in active CD. However, in that study, IL-15 secretion was not tested directly on enterocytes isolated from untreated CD patients but on the T84 intestinal epithelial cell line and on organ culture of intestinal biopsy specimens from treated CD patients incubated with gliadin.16 The very high affinity binding between IL-15 and IL-15Rα,22 the IL-15 receptor α chain that we found upregulated on CD IEL, might explain the apparent lack of secreted IL-15 in the organ culture supernatants of CD biopsies challenged with gliadin. Our data, together with the demonstration that Th1 cytokines, which are known to be overexpressed in active CD mucosa,24,25 are capable of inducing activation of a specific protease that cleaves cell surface IL-15,26 support the hypothesis that IL-15 may be secreted by enterocytes in untreated CD. On this basis, and in agreement with findings on human peripheral blood monocytes,23 we suggest that, under physiological conditions, IL-15 exerts its biological effects mainly as a membrane bound form at the epithelial level. In contrast, in active CD, IL-15 may be released by enterocytes, either as a consequence of Th1 cytokine induced proteolytic shedding of the membrane anchored form or on cell death.
IL-15 is a potent activator of normal IEL, stimulating both their proliferation and IFN-γ secretion.2 Our findings provide evidence that IEL from untreated CD patients are more susceptible to stimulation by IL-15 than those from treated CD patients and controls, showing in response to IL-15 treatment increased proliferation and greater release of IFN-γ and TNF-α. In particular, of these two cytokines, IFN-γ seems to be much more involved in the generation of gluten driven mucosal damage in CD, as indicated by the efficacy of anti-IFN-γ antibodies in preventing villous atrophy.27
In the normal intestine, where enterocytes are exposed to a wide array of pathogens, IL-15 has a role in enhancing IEL cytotoxicity and in modulating perforin/granzyme and NFG2D signalling pathways in order to maintain a healthy epithelium by eliminating infected cells.2,28,29 Additionally, IL-15 is known to induce the lymphokine activated killer activity of human peripheral blood lymphocytes and the killing of murine intestinal epithelial cells by a perforin dependent pathway.30,31 In a pathological condition such as CD, characterised by overexpression of IL-15 by enterocytes associated with upregulation of IL-15Rα on IEL, promotion of perforin/granzyme mediated killing might lead to epithelial cell damage and, then, villous atrophy. As we previously reported higher infiltration of perforin+ IEL in active CD, which significantly correlated with the degree of enterocyte apoptosis,32 we investigated in vitro whether IL-15 was capable of activating the perforin/granzyme dependent cytotoxicity provided by IEL against epithelial cells. Our results, in accordance with those reported in refractory sprue,16 showed that in untreated CD, the IL-15 induced killing action of IEL against epithelial cells was higher than in treated CD and controls. The fact that blocking the perforin pathway with chloroquine, an inhibitor of granzyme/perforin dependent cytotoxicity,19 completely suppressed IEL cytotoxic activity against Caco-2 cells clearly indicates that this cytolytic mechanism is perforin dependent. Consistent with this, blocking IL-15 reduced in a concentration dependent manner the cytotoxic activity of untreated CD IEL against Caco-2 cells and significantly decreased granzyme B release in IEL supernatants. Taken together, these findings are in keeping with those of Meresse and colleagues33 who showed that IL-15 can convert cytotoxic T lymphocytes into lymphokine activated killer cells through induction of an NKG2D signalling pathway both in vitro and in vivo in CD.
To our knowledge, this is the first study to explore LPMC secretion of IL-15 in vitro in CD. Previous immunohistochemical studies of IL-15 expression in CD lamina propria, both affected by the subjective quantification of immunostaining, produced discrepant results, showing either positivity of some scattered mononuclear cells or massive upregulation of LPMC expression.13,16 We observed higher LPMC production of IL-15 in active CD, as demonstrated by immunoblotting and by culture of LPMC stimulated with LPS or IFN-γ. IL-15 protein, not measurable in the supernatants of unstimulated control and treated CD LPMC, was detectable in unstimulated LPMC from untreated CD patients, allowing us to infer that this cytokine is secreted in the lamina propria of active CD patients. As IL-15 plays a crucial role in the maturation and activation of dendritic cells and macrophages,5 it is conceivable that in CD, in addition to its role in activating the innate response by promoting TCR independent activation and cytotoxicity of IEL, it might amplify the adaptive response to gluten by enhancing the ability of dendritic cells to present gliadin peptides to CD4+ T cells (fig 8).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Possible model of the pathogenic role of interleukin 15 (IL-15) in modulating both innate (shaded panel) and adaptive arms of the immune response in coeliac disease. APC, antigen presenting cell; DC, dendritic cell; IEL, intraepithelial lymphocyte; LPMC, lamina propria mononuclear cell; TCR, T cell receptor.
Activated T cells undergo apoptosis on repeated stimulation. This activation induced cell death is an important mechanism underlying T cell homeostasis after an immune response,34 particularly in the intestine where IEL activation through TCR by foreign and self antigens is likely to occur constantly.35 In the present study, we demonstrated that exogenous IL-15 prevented apoptosis of normal IEL, while the decreased propensity of IEL to undergo apoptosis in active CD, shown in a previous study of ours,18 was reverted by blocking endogenous IL-15 with a neutralising antibody. The antiapoptotic effect of IL-15 reported in this study is consistent with previous observations in which IL-15 promoted the survival of human peripheral blood B and T cells on cross linking of Fas or antigenic receptors,36 and of activated intestinal murine γ/δ IEL.37 IL-15 induced resistance to apoptosis that we found in CD IEL is particular intriguing when one considers that in refractory sprue, an established complication of CD characterised by an abnormal IEL population with clonal rearrangement of the TCR-γ, IL-15 promotes the in vitro survival and expansion of clonal CD103+sCD3− IEL, which may ultimately transform a refractory form into an aggressive T cell lymphoma.16 Accordingly, we found high IL-15 content in the intestinal mucosa of patients with complicated CD, four of them characterised by clonal rearrangement of the TCR-γ.
In conclusion, our findings suggest that IL-15 plays a pivotal role in the development of CD. Overexpression of IL-15 in active CD may increase the number of inflammatory cells involved in the generation of epithelial damage by stimulating proliferation, Th1 cytokine production, and cytotoxicity of IEL, and by protecting these cells from apoptosis. These findings together with the recent demonstration by Maiuri and colleagues13 that the non-immunodominant peptide p31–43, but not the immunodominant epitopes, is able to induce rapid expression of IL-15 in tissues from patients with CD, strongly suggest that this cytokine is involved in modulating the innate immune response to gliadin in CD. Moreover, the pro-survival effect of IL-15 on IEL may predispose to the emergence of T cell clonal proliferations in CD, thus promoting the transformation of low grade/“cryptic” enteropathy associated T cell lymphomas-refractory sprue and ulcerative jejunoileitis38 into high grade lymphomas sharing the same clonality.
Dissecting the complex machinery of the IL-15/IL-15R system will provide interesting answers to the numerous questions raised by IL-15 involvement in the pathogenesis of CD and will open up new possibilities for the development of therapeutic strategies aimed at suppressing uncontrolled IEL activation and survival by blocking IL-15 or its receptor, thus preventing both tissue damage and lymphomagenesis that occur in CD.
Acknowledgments
The authors thank Dr Federico Biagi for providing blinded tissue samples from some of the patients with uncomplicated and complicated coeliac disease.
REFERENCES
Footnotes
-
Published online first 16 August 2005
-
Conflict of interest: None declared.