Article Text

Abstract
Objectives: Diarrhoea-predominant irritable bowel syndrome (IBS-D) is characterised by elevated colonic lumenal serine protease activity. The aims of this study were (1) to investigate the origin of this elevated serine protease activity, (2) to evaluate if it may be sufficient to trigger alterations in colonic paracellular permeability (CPP) and sensitivity, and (3) to examine the role of the proteinase-activated receptor-2 (PAR-2) activation and signalling cascade in this process.
Patients and methods: Faecal enzymatic activities were assayed in healthy subjects and patients with IBS, ulcerative colitis and acute infectious diarrhoea. Following mucosal exposure to supernatants from control subjects and IBS-D patients, electromyographic response to colorectal balloon distension was recorded in wild-type and PAR-2–/– mice, and CPP was evaluated on colonic strips in Ussing chambers. Zonula occludens-1 (ZO-1) and phosphorylated myosin light chain were detected by immunohistochemistry.
Results: The threefold increase in faecal serine protease activity seen in IBS-D patients compared with constipation-predominant IBS (IBS-C) or infectious diarrhoea is of neither epithelial nor inflammatory cell origin, nor is it coupled with antiprotease activity of endogenous origin. Mucosal application of faecal supernatants from IBS-D patients in mice evoked allodynia and increased CPP by 92%, both of which effects were prevented by serine protease inhibitors and dependent on PAR-2 expression. In mice, colonic exposure to supernatants from IBS-D patients resulted in a rapid increase in the phosphorylation of myosin light chain and delayed redistribution of ZO-1 in colonocytes.
Conclusions: Elevated colonic lumenal serine protease activity of IBS-D patients evokes a PAR-2-mediated colonic epithelial barrier dysfunction and subsequent allodynia in mice, suggesting a novel organic background in the pathogenesis of IBS.
Statistics from Altmetric.com
Irritable bowel syndrome (IBS) is a functional gastrointestinal disorder characterised by abdominal pain or discomfort associated with altered bowel habits, and a “low-grade” mucosal inflammation which may be linked to enhanced exposure to antigens due to increased gut permeability. It has been reported to affect 5–20% of the population worldwide,1 thus increasing interest has recently been shown towards the poorly understood pathophysiology of and possible therapeutic approaches to the disease.
In a preliminary pilot study, we have shown that faecal supernatants of diarrhoea-predominant IBS (IBS-D) patients have a substantially higher serine protease activity, similar to that of ulcerative colitis (UC) patients, when compared with healthy subjects or other subgroups of IBS patients.2 Certain serine proteases are signalling molecules that activate protease-activated receptors (PARs). Four members of the PAR family have been identified in human tissues, of which PAR-2 is highly expressed on intestinal epithelial cells, particularly at the apical side.3 ,4 The primary activating protease of PAR-2 is trypsin; however, serine proteases of other endogenous and/or bacterial origins are also able to cleave the receptor.5 Activation of PAR-2 modulates several gastrointestinal functions, such as motility, ionic exchange, paracellular permeability, sensory functions and inflammation.6 Intracolonic but not intraperitoneal administration of the synthetic selective PAR-2 agonist, SLIGRL (H-serine-leucine-isoleucine-glycine-arginine-leucine-OH), produces delayed visceral hyperalgesia in rats associated with increased paracellular permeability.7–9 Both visceral hypersensitivity10–12 and impaired intestinal permeability13 ,14 are commonly observed features of IBS, particularly in the IBS-D subgroup,15 the same subpopulation where we detected increased faecal serine protease activity.2
The integrity of the epithelial barrier is dependent upon the contraction of the perijunctional actomyosin ring and subsequent physical tension on the tight junction, an event that involves the phosphorylation of myosin light chain (MLC), initiated by MLC kinase (MLCK).16 ,17
Therefore, we conducted this study (1) to investigate the origin of elevated faecal serine protease activity in IBS-D patients, (2) to evaluate if this elevated colonic lumenal serine protease activity may be sufficient to trigger alterations in colonic permeability and sensitivity in mice and to analyse its effects on MLC phosphorylation and the tight junction protein zonula occludens-1 (ZO-1), and (3) to examine the possible involvement of PAR-2 activation in this process.
METHODS
Study participants and faecal samples collection
Demographic data of the patients enrolled in the study are summarised in table 1. Patients fulfilling the Rome II criteria for IBS participated in the study.18 All participants provided medical history and underwent physical examination. Organic gastrointestinal disorders were excluded by detailed blood and stool analyses, serological assays for coeliac disease, lactose–hydrogen breath test and colonoscopy. Active UC was assessed clinically and endoscopically. The study protocol was approved by the Ethical Committee of the University of Szeged. All subjects provided written and informed consent to participate. This cohort was an extension from a previous published study by our group.2 Patients of this second series were enrolled from 7 to 9 months after patients from the first series already published, and all of them were different from those in the first series. The overlap was 61%, as described in table 1.
Only faeces collected in situ, or collected at home and transported within 1 h after defecation to the First Department of Internal Medicine in Szeged, Hungary, were used. Samples were stored at −80°C until transportation on dry ice to INRA Toulouse, France.
Sample preparation
Upon arrival, 1 g of faecal sample was thawed, dissolved, and homogenised in 7 ml of Tris buffer, centrifuged (4500 rpm, 10 min, +4°C) and filtered (0.2 μm, Nalgene, (Adventure 16, San Diego, California, USA)). The acquired supernatants were used for measuring total protease activity, serine protease activity, polymorphonuclear neutrophil (PMN) and pancreatic elastase, myeloperoxidase (MPO), calprotectin and human secretory leucocyte protease inhibitor (SLPI) activity.
Measurement of faecal enzymatic activities
To measure total faecal serine protease activity, supernatants of faecal homogenates (25 μl) were incubated with 1 ml of reaction buffer (0.15 M NaCl and 20 mM Tris-HCl, pH 8.3) and 1 ml of 0.5% (w/v) azocasein (Sigma, St Quentin Fallavier, France) at 40°C. The reaction was stopped after 20 min with 1 ml of 10% (v/v) trichloracetic acid (TCA, Sigma). Following centrifugation, absorption of the clear supernatant was measured at 366 nm. Enzymatic activities of the supernatants were normalised to protein content. To determine whether protease activity was dependent upon serine proteases, measurements were done after preincubation for 30 min with selective serine protease inhibitors, AEBSF (4-(2-aminoethyl) benzenesulphonyl fluoride hydrochloride) and a mixture of soybean trypsin inhibitor (SBTI) and aprotinin (Sigma). Protease activity was expressed as units per milligram of protein, standardised against activity elicited by 1 U of standard trypsin.
Pancreatic elastase-1, SLPI, PMN elastase and calprotectin concentrations were assayed by ELISA (Schebo-Tech, Giessen, Germany; R& D Systems, Lille, France; Immundiagnostik AG, Bensheim, Germany; HyCult Biotechnology, Uden, The Netherlands, respectively). Faecal MPO activity was measured as described earlier.19
Animals
Congenic 6–9-week old male C57BL/6J wild type (Janvier, Le Genest St-Isle, France) and PAR-2-deficient mice (The Jackson Laboratory, Bar Harbor, Maine, USA) were used in this study. The genetic status of the PAR-2-deficient mice was confirmed by PCR (data not shown). Mice were housed in polypropylene cages in a light- and temperature-controlled room (12 h/12 h cycles; 20±2°C), were fed standard pellets (Harlan Teklad, Bicester, Oxon, UK), and water was provided ad libitum. The experimental protocols described in the study were approved by the local Institutional Animal Care and Use Committee.
Visceral sensitivity
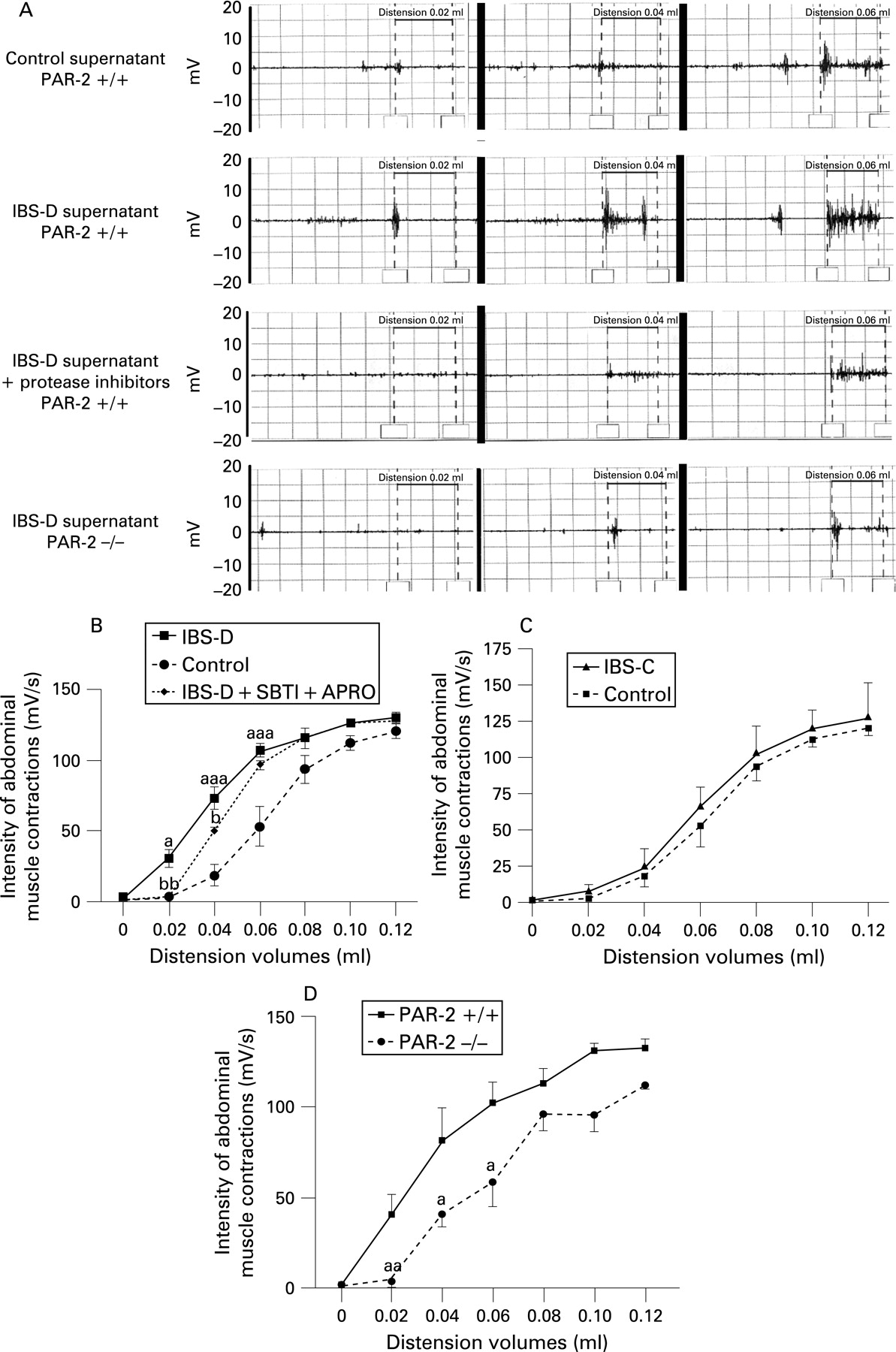
Under xylazine/ketamine anaesthesia (both 1.2 mg, subcutaneously), two nickel–chrome electrodes were implanted into the abdominal external oblique muscle and a third into the abdominal skin, and were exteriorised on the back of the neck. On the fifth to seventh postoperative day, colorectal distensions were used as noxious stimuli to evaluate visceral hyperalgesia by electromyographic (EMG) recording. Under sodium pentobarbital anaesthesia (10 mg, intraperitoneally), polyethylene perfusion and distension catheters (Fogarty catheter for arterial embolectomy, 4F, Edwards Lifesciences, Nijmegen, The Netherlands) were inserted into the colon. Animals received 0.3 ml of faecal supernatants of IBS-D or IBS-C (constipation-predominant IBS) patients, of healthy subjects or of IBS-D patients previously incubated with serine protease inhibitors, SBTI and aprotinin. The colorectal distension procedure started 60 min after the infusion of the supernatants had finished with volumes progressively increasing in 0.02 ml steps, from 0 to 0.12 ml, each step lasting 10 s with 5 min non-distension periods in between. During the distension periods, the striated muscle’s EMG activity was recorded and analysed according to Larsson et al.20 Basal EMG activity was subtracted from the EMG activity registered during the periods of distension.
In vitro permeability
Mice were sacrificed by cervical dislocation, and the distal part of the colon was removed. Colonic strips were mounted with a flux area of 0.3 cm2 in Easymount Ussing-type chambers (Physiologic Instruments, San Diego, California, USA), bathed in Krebs solution and oxygenated at a maintained temperature of 37°C. After allowing 15 min for equilibrium, one-fifth of the initial volume of the buffer solution of the apical compartment (mucosal side) was replaced with supernatants and fluorescein isothiocyanate (FITC)-labelled dextran (4000 Da, 0.022 g/ml, Sigma). One hour later, fluorescent intensity was measured on the serosal side of the chamber.
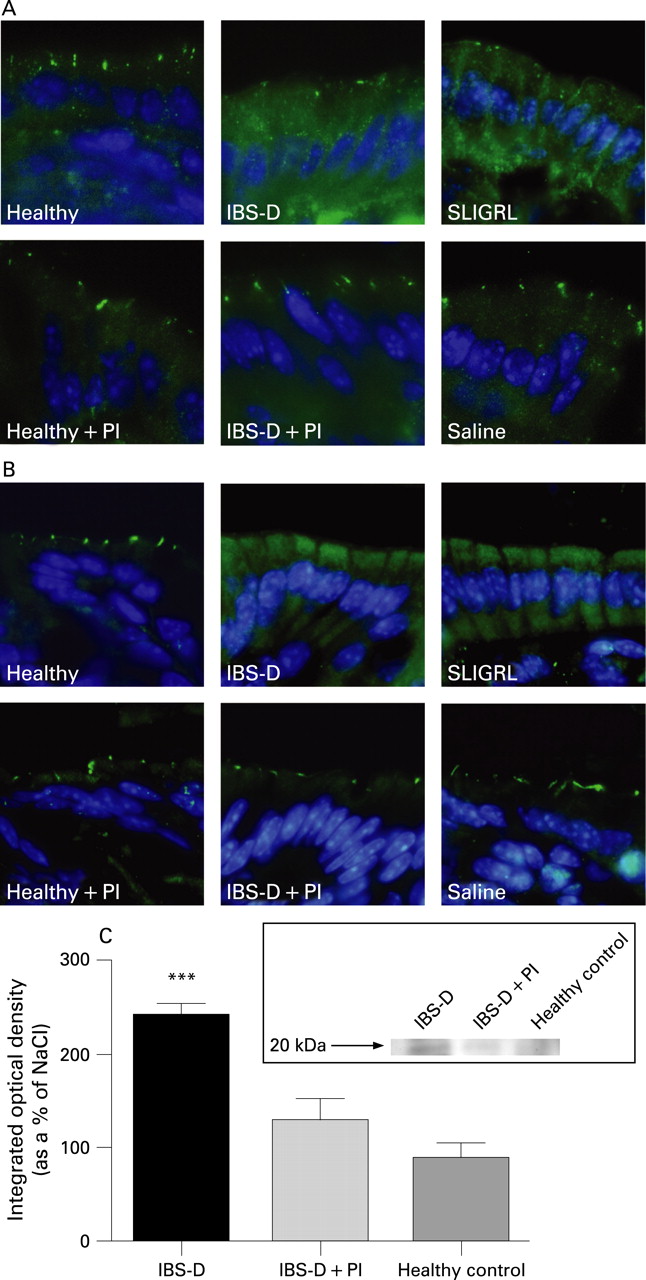
Immunohistochemistry of pMLC and ZO-1
At 1 and 4 h after the intracolonic infusion of supernatants of IBS-D patients or healthy controls, saline, SLIGRL (5 μg) or IBS-D supernatants previously incubated with serine protease inhibitors, the mice were sacrificed and the distal colon was removed.
For both phosphorylated MLC (pMLC) and ZO-1 immunolabelling, samples were fixed in buffered paraformaldehyde (4%), incubated in 30% sucrose (24 h, +4°C), embedded (Tissue Tek medium) and frozen in isopentane at −45°C. Cryostat sections (7 μm) were fixed with acetone (10 min, −20°C), hydrated in phosphate-buffered saline (PBS) and treated with 4 mg/ml sodium borohydride (45 min, +4°C). Sections were permeabilised with PBS–0.5% Triton X-100 and incubated in blocking solution (PBS containing 1% bovine serum albumin). For pMLC staining, samples were then incubated with goat anti-pMLC antibodies (1/100, SantaCruz, Santa Cruz, California, USA) followed by incubation with biotin-conjugated IgG donkey anti-goat antibody (1/1000, Interchim, Montlucon, France). Sections were rinsed in NaHCO3 (0.1 M, pH 8.2) and incubated with FITC-conjugated avidin (1/500) diluted in the same solution.
For ZO-1 labelling, sections were incubated with rabbit anti-ZO-1 antibodies (1/500, Zymed, San Francisco, California, USA) followed by incubation with Alexa fluor 488-conjugated IgG donkey anti-rabbit antibodies (1/2000, Molecular Probes, Cergy-Pontoise, France). All sections were mounted in Vectashield HardSet Mounting Medium with 4′,6-diamidino-2-phenylindole (DAPI; Vector Laboratories, Burlingame, California, USA) and examined under a Nikon 90i fluorescent microscope.
Western blot for pMLC
Colonic mucosa was collected from mice 1 h after intracolonic infusion with faecal supernatants of healthy subjects or IBS-D patients, or saline. Proteins were extracted with RIPA buffer and quantified. Following the Laemmli method,21 equal amounts of protein extracts were electrophoresed by 12% SDS–PAGE (sodium dodecyl sulphate–polyacrylamide gel electrophoresis) and then electrotransferred onto Hybond-P membrane (GE Healthcare, Bordeaux, France). After saturation, the membrane was incubated with anti-pMLC primary antibody (1/1000, Biosource, Worcester, Massachusetts, USA) and peroxidase-conjugated goat anti-rabbit secondary antibody (1/1000, Millipore, St Quentin en Yvelines, France). The membrane was developed with SuperSignal Reagent (Pierce, Prebières, France). Integrated density values were assessed by ImageJ 1.37 software (NIH, Bethesda, Maryland, USA).
Statistical analysis
All data are presented as means (SEM). For statistical analysis, Prism 4.0 (GraphPad, San Diego, California, USA) was used. Multiple comparisons for faecal enzymatic activities of different patient groups and integrated optical density values of pMLC western blots were analysed by repeated measures of one-way analysis of variance (ANOVA), followed by Tukey test or Kruskal–Wallis post-test (for MPO activity). Statistical significance for visceral hypersensitivity results was established by using two-way ANOVA, followed by Bonferroni post-test. Permeability results were analysed with one-way ANOVA, followed by Tukey post-test or two-tailed unpaired t test.
RESULTS
Faecal serine protease activity
In healthy subjects the total faecal serine protease activity was 698 (128) U/mg of protein. This serine protease activity was significantly greater in IBS-D patients (2079 (334) U/mg, p<0.001) and in UC patients (2193 (430), p<0.01) compared with healthy controls; a similar increase was not present regarding the IBS-C, IBS-A subgroups or patients with infectious acute diarrhoea (INF) (fig 1A). In addition, these differences between IBS-D patients and healthy subjects were found when studying only our second series of patients (data not shown). Adding the serine protease inhibitor AEBSF abolished this increased protease activity in IBS-D and UC supernatants. A similar inhibition was obtained with preincubation with SBTI and aprotinin, two common serine protease inhibitors validated for human use (fig 1B). Regarding the potential origin of the elevated serine protease activity, no significant difference was observed in faecal pancreatic elastase-1 concentration in UC or in any subgroups of IBS patients when compared with controls (fig 1C). However, both the faecal trypsin and pancreatic elastase-1 concentration was significantly decreased in INF patients (p<0.05), which might be attributed to the diluted faecal content (ie, profuse watery diarrhoea) and restricted diet. PMN-derived elastase had a significantly elevated faecal concentration in UC and INF patients (p<0.05), but not in any subgroup of IBS patients (fig 1D). Human faecal SLPI activity showed no significant difference in any of the studied groups of patients when comparing them with control subjects (fig 1E). Faecal inflammatory markers, such as human calprotectin and MPO, showed a significant increase in UC (p<0.001 and p<0.05, respectively) and INF patients (p<0.05 and p<0.001, respectively), but we found no increase in any of the IBS subgroups (fig 1F,G)

Visceral hypersensitivity is triggered by elevated serine protease activity of IBS-D supernatants and is dependent on mucosal PAR-2 expression
Intracolonic infusion of faecal supernatants from IBS-D patients administered prior to rectosigmoidal distensions in mice significantly increased the abdominal muscle EMG response (a valid criterion representing nociception, fig 2A) at low distension volumes, namely at 0.02, 0.04 and 0.06 ml, compared with the intensity of muscle contractions in animals treated with faecal supernatants of control subjects (0.02 ml, 30.6 (5.9) mV/s vs 3.5 (1.7) mV/s; 0.04 ml, 73.4 (7.9) mV/s vs 18.9 (7.5) mV/s; 0.06 ml, 107.4 (4.4) mV/s vs 53.2 (14.3) mV/s; p<0.05, p<0.001, p<0.001, respectively, fig. 2B). Conversely, colonic instillation of IBS-C supernatants did not evoke any modification of visceral sensitivity to colorectal distension (fig 2C). Incubation of faecal supernatants from IBS-D patients with serine protease inhibitors prior to colonic infusion prevented the increased EMG response to low volumes of distension (0.02 ml, 2.8 (1.4)mV/s, 0.04 ml, 49.8 (2.9) mV/s; p<0.01, p<0.05, respectively), while protease inhibitors per se had no effect (data not shown). Furthermore, IBS-D supernatants failed to induce allodynia or visceral hypersensitivity in PAR-2-deficient mice (0.02 ml, 4.6 (3.5) mV/s; 0.04 ml, 41.4 (7.0) mV/s; 0.06 m, 59.4 (14.2) mV/s, fig 2D).

Increase in colonic paracellular permeability (CPP) is evoked by elevated serine protease activity of IBS-D supernatants and is PAR-2 dependent
Administration of faecal supernatants from healthy subjects to the mucosal side of mouse colon mounted in Ussing chambers did not significantly alter CPP compared with saline (128% (16%) vs 100% (6%); NS). In contrast, addition of faecal supernatants from IBS-D patients significantly increased the FITC–dextran flux compared with the administration of supernatants from healthy subjects or saline (192% (13%), p<0.05). Comparatively, supernatants from IBS-C patients failed to induce such an increase in colonic permeability (137% (19%); NS, fig 3A). Previous incubation of the supernatants from IBS-D patients with SBTI and aprotinin prior to administration significantly decreased the elevated CPP evoked by supernatants from IBS-D patients (111% (17%) vs 202% (16%), p<0.01, fig 3B). No increase in CPP was observed in response to faecal supernatants from IBS-D patients on colonic strips of PAR-2-deficient mice in contrast to their wild-type strain (127% (28%) vs 248% (42%), p<0.05, fig 3C).

Increased rapid phosphorylation of MLC and delayed redistribution of ZO-1 in epithelial cells after mucosal exposure to IBS-D supernatants
One hour after intracolonic infusion with supernatants from IBS-D patients there was a pronounced and diffuse labelling of pMLC in epithelial cells, similar to that observed after the infusion of SLIGRL used as a positive control for PAR-2 activation, and inhibited by preincubation with serine protease inhibitors. Administration of supernatants from healthy subjects or saline failed to evoke an increase in pMLC immuno-staining, which remained restricted to the tight junction area (fig 4A).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Four hours after intracolonic infusion of supernatants from healthy subjects or saline, selective immunostaining of the tight junction protein ZO-1 showed labelling that was restricted to the surface of epithelial cells. In contrast, intracolonic infusion of supernatants from IBS-D patients resulted in a marked labelling of the intracellular compartment, similarly to that experienced after the infusion of SLIGRL, suggesting intensive internalisation of the protein. Previous incubation of faecal supernatants from IBS-D patients with serine protease inhibitors prevented the occurrence of a marked intracellular labelling of colonocytes (fig 4B).
Western blotting showed an increased level of pMLC in the colonic mucosa of mice 1 h after the intracolonic infusion of supernatants from IBS-D patients compared with those infused with supernatants from healthy controls (p<0.001). There was no significant difference in the level of pMLC in the colonic mucosa of mice that were infused with supernatants from healthy subjects or IBS-D supernatants previously incubated with a mixture of serine protease inhibitors (fig 4C).
DISCUSSION
In the present study we have shown that serine protease activity can be used as a marker to distinguish between IBS and acute infectious diarrhoea, and the absence of increased faecal inflammatory markers permits the differentiation of IBS-D patients from UC patients. Our present investigations provide evidence that faecal supernatants from IBS-D patients are able to evoke visceral hypersensitivity in mice when applied intracolonically, and to increase CPP in vitro. Both of these effects are prevented by the administration of serine protease inhibitors and are dependent on PAR-2 expression, since they are absent in PAR-2-deficient mice. We also showed that intracolonic infusion of supernatants from IBS-D patients increases the phosphorylation of MLC, a process that is known to be involved in disrupting the integrity of epithelial tight junctions.16 Furthermore, the tight junction protein ZO-1 showed internalisation in vivo in colonocytes in response to mucosal exposure to IBS-D faecal supernatants.
In agreement with our previous pilot study, the increase in faecal serine protease activity is similar in patients with IBS-D and those with active UC.2 Interestingly, patients with acute transient infectious diarrhoea are characterised by a low level of serine protease activity, suggesting that in IBS-D patients elevated enzymatic activity is not due to an accelerated intestinal transit. Of interest, IBS-A patients share with IBS-C patients a low level of faecal serine protease activity, and this raises the question of the significance of IBS-A status. Indeed, their pattern of faecal enzymatic activity was very close to that of IBS-C patients, suggesting a common pathophysiological origin. In agreement with previous studies,22–24 we found elevated levels of inflammatory markers in the faecal samples of UC and INF patients, but none of these markers were increased in any of the IBS subgroups. The high lumenal serine protease activity in the faecal samples of IBS-D patients is neither associated with elevated enzymatic activity of pancreatic or inflammatory cell origin nor coupled with decreased antiprotease activity (SLPI), which could disturb the lumenal balance between proteases and antiproteases. This is in contrast to recently published results on elevated mast cell tryptase activity found in biopsy samples from IBS patients.25 However, the authors noted an increased proteolytic activity in colonic washes, but did not characterise its origin, and we cannot exclude in their study that the elevated activity was due to exposure to hyperosmotic laxatives used for colonoscopy preparation. In our previous report using faecal material and not colonic washes, no increase in mast cell tryptase was noted in faecal supernatants from IBS-D patients.2 Thus, the origin of the elevated serine protease level needs further evaluation, though our data suggest a microbial origin. Indeed bacteria contribute substantially to the production of colonic serine proteases,26 and quantitative as well as qualitative alterations in faecal microbiota, namely a decrease in lactobacilli, were described in IBS-D patients27 ,28 who have high antiprotease activity. Moreover, reduced colonic microflora obtained by oral antibiotic treatment resulted in a lower serine protease activity and was associated with a decreased expression of PAR-2 on the colonic epithelial cells of mice.29 PAR-2 has been reported to be activated by pathogen proteases as well, such as Porphyromonas gingivalis-produced gingipain that mediates inflammatory events in the pathogenesis of periodontitis through PAR-2.30
Enhanced colonic sensitivity to distension in IBS patients was first demonstrated in 197311 and, since then, numerous studies have confirmed altered gut sensitivity in IBS patients.10 ,12 ,31 In rats, PAR-2 activation with intracolonic infusion of PAR-2-activating peptide SLIGRL or trypsin provokes delayed long-lasting colorectal hypersensitivity to distension, which is associated with increased CPP.9 Our present study shows that allodynia is evoked by faecal supernatants of the IBS-D subgroup of patients, by the low volumes of distension seen in mice. This effect was fully prevented by the administration of serine protease inhibitors at 0.02 ml, and partially at 0.04 ml. In accordance with these results, IBS-D supernatants were not able to evoke any hypersensitivity in PAR-2-deficient mice, further supporting the role of serine proteases in the initiation of visceral hypersensitivity, despite the fact that we cannot fully exclude that other luminal factors, independently of serine proteases, might also be involved in alterations of visceral sensitivity.
Altered intestinal permeability has been described by several clinical studies as another characteristic feature of IBS. Impaired intestinal permeability was detected in postinfectious IBS patients. Furthermore, patients with impaired intestinal permeability were more likely to report increased stool frequency.13 These data confirmed earlier findings of altered gut permeability following acute Campylobacter enteritis and postdysenteric IBS in a larger population.14 A recent study emphasises that the increase in intestinal permeability is more pronounced in IBS-D patients than in other subgroups of IBS.15 Intracolonic administration of a serine protease inhibitor, aprotinin, resulted in reduced CPP in mice.29 In agreement with this, we show here that supernatants of IBS-D patients are able to evoke increased CPP in colonic strips of mice, an effect inhibited by serine protease inhibitors, which further supports the role of serine proteases in impaired gut permeability. Accordingly, supernatants of IBS-C patients or healthy subjects, who lack elevated serine protease activity, failed to induce such permeability changes. Some studies have reported that PAR-2 participates directly in the pathogenesis of IBS.6 Our present data on in vitro permeability studies in PAR-2-deficient mice clearly demonstrate that the permeability changes induced by supernatants from IBS-D patients with high serine protease activity are mediated through PAR-2.
Increased intestinal permeability when triggered by intracolonic PAR-2 activation was shown to be due to MLC phosphorylation.25 Our immunohistochemical studies revealed that colonic exposure to supernatants of IBS-D patients, with high protease activity, triggers a rapid phosphorylation of MLC and a subsequent, delayed internalisation of ZO-1 in colonocytes in vivo. This further supports the hypothesis of a serine protease-mediated mechanism in the alteration of tight junction permeability. Dysregulation of epithelial barrier function leads to increased exposure to lumenal antigens, bacterial translocation and activation of the mucosal immune system. Of interest, low grade inflammation of the intestinal mucosa has recently been verified by several studies on IBS.32–37 Therefore, we may speculate that release of inflammatory mediators, as a consequence of low-grade mucosal inflammation, is sufficient to sensitise sensory neuron nerve terminals, possibly resulting in a decreased threshold for visceral sensitivity in IBS patients.
In conclusion, we propose that serine protease activity represents a new biomarker to distinguish IBS-D from infectious diarrhoea and, in the absence of inflammatory markers, from inflammatory bowel diseases. We showed that the elevated serine protease activity seen in IBS-D patients is sufficient to trigger a PAR-2-mediated increase in colonic permeability and subsequent visceral hypersensitivity in mice, suggesting similar effects in humans. Thus, these data support an organic background for IBS offering novel therapeutic approaches in the treatment of the disease considering lumenal serine protease inhibition or PAR-2 antagonism as valid therapeutic targets.
Acknowledgments
The authors thank Ferenc Nagy, Tamás Molnár, Zsuzsanna Lénárt, Domonka Fodor and Ildikó Kovács for patient screening, and Bernard Joseph for technical assistance.
REFERENCES
Footnotes
-
Competing interests: None.
-
Funding: KG is a recipient of a postdoctoral fellowship from INRA. This work was supported by an institutional grant from INRA.
-
Patient consent: All subjects provided written and informed consent to participate.
-
Ethics approval: The study protocol was approved by the Ethical Committee of the University of Szeged. The animal experimental protocols were approved by the local Institutional Animal Care and Use Committee.