Article Text

Abstract
Objective Autoimmune hepatitis and primary sclerosing cholangitis are chronic inflammatory disorders of unknown aetiology, frequently associated with the presence of perinuclear antineutrophil cytoplasmic antibodies (p-ANCAs) directed against an unknown antigen of myeloid cells.
Methods and Results Here, it is reported that p-ANCAs in autoimmune liver disorders react with β-tubulin isotype 5 (TBB-5) as autoantigen as well as with its evolutionary bacterial precursor protein FtsZ. Both proteins were confirmed as antigens of p-ANCAs in autoimmune liver disorders by demonstrating reactivity of ANCA-positive sera with recombinant TBB-5 (72–88%) and FtsZ (64–82%) on immunoblots and antigen-specific abrogation of ANCA immunofluorescence when sera had been preabsorbed with tubulin and FtsZ. Using sera from interleukin 10-deficient mice (Il10–/–), an animal model of inflammatory bowel disease, it was also demonstrated that antibodies against TBB-5 are generated in response to intestinal microorganisms. However, unlike autoimmune liver disorders, human antibodies to FtsZ in the absence of TBB-5 antibodies were also a frequent finding in non-autoimmune liver diseases (up to 95%). Reactivity to TBB-5 without the presence of FtsZ antibodies was found in very few cases (<1%) in autoimmune liver disorders.
Conclusions Thus, p-ANCAs in autoimmune liver diseases are directed against human TBB-5 cross-reacting with the bacterial protein FtsZ, probably reflecting an abnormal immune response to intestinal microorganisms in susceptible, possibly genetically predisposed individuals.
- ANCA
- autoimmune hepatitis
- autoimmunity
- primary sclerosing cholangitis
Statistics from Altmetric.com
Introduction
Autoimmune hepatitis (AIH) and primary sclerosing cholangitis (PSC) represent chronic inflammatory disorders of unknown aetiology. They are considered as immune-mediated disorders, because aberrant immune functions are found in a high proportion of affected patients. These abnormalities comprise a high prevalence of autoantibodies (96%) such as perinuclear antineutrophil cytoplasmic autoantibodies (p-ANCAs) and a frequent association with other putative autoimmune diseases such as ulcerative colitis.1–6 However, the role of autoantibodies in the immunopathogenesis of autoimmune liver disorders is debated controversially, particularly with respect to ANCAs because their underlying autoantigen has remained elusive. Recently, we clarified that p-ANCAs in AIH and PSC differ from p-ANCAs associated with vasculitides by a distinct pattern of perinuclear immunofluorescence combined with intranuclear fluorescent spots.7–10 Our own previous data pointed to an acidic myeloid cell-specific protein with an approximate molecular weight of 50 kDa localised at the inner side of the nuclear envelope as the antigen of p-ANCAs in autoimmune liver disorders and chronic inflammatory bowel diseases (IBDs).11 12 Unlike all other proteins thus far proposed as candidate antigens of p-ANCAs in autoimmune liver disorders, the here identified protein was detected in >90% of the sera positive for p-ANCAs.13
Here, we describe the molecular identification of this 50 kDa ANCA autoantigen as β-tubulin isotype 5 (TBB-5). The identified target antigen shares an extraordinarily high structural homology with the microbial cell division protein FtsZ, which represents an evolutionary precursor protein of β-tubulin, present in almost all bacteria of the intestinal microflora.14–16 This fact prompted us to test the hypothesis of whether ANCA autoreactivity in autoimmune liver disorders might reflect cross-recognition between FtsZ and TBB-5.
Methods
Patients
Serum samples from 134 ANCA-positive patients with autoimmune liver disorders (PSC n=39, AIH n=95) were enrolled in this study. For control purposes, 30 ANCA-positive sera from patients with ulcerative colitis, 37 serum samples from ANCA-negative healthy controls as well as 84 serum samples from disease controls (non-cirrhotic chronic hepatitis B (HBV) n=22, non-cirrhotic chronic hepatitis C (HCV) n=19, alcoholic liver cirrhosis n=43) were included (table 1). All sera were stored at −20°C until analysis. Diagnoses of the patients were based on established clinical, endoscopic, histological, radiological and serological criteria and scores.17–19 All study procedures were executed in accordance with the current revision of the Declaration of Helsinki of 2000. The study was approved by the local institutional review board at the University of Bonn.
Characteristics of the study population
Immunofluorescence microscopy for detection of ANCAs
Detection of ANCAs was performed on commercially available slides with ethanol-fixed human neutrophils (INOVA Diagnostics, La Jolla, California, USA) according to the guidelines of the First International Workshop on ANCAs.7 20 Accordingly, the slides were incubated with serum samples, diluted 1:10 in phosphate-buffered saline (PBS). All staining patterns were independently read by two investigators, who were unaware of the clinical diagnosis of the patients. Three different staining patterns were distinguished: a diffuse staining of the cytoplasm (c-ANCAs), a fine rim-like fluorescence of the perinuclear cytoplasm (‘classical’ p-ANCAs) and an inhomogeneous rim-like fluorescence of the nuclear periphery along with multiple intranuclear fluorescent spots (previously termed ‘atypical’ p-ANCAs).7 11 Serum end point titres ≥1:10 were considered positive.
Cells
Human promyelomonocytic HL-60 cells (American Tissue Cell Collection (ATCC), CCL-240; Manassas, Virginia, USA), HeLa cells, a human epithelial cervix cancer cell line (ATCC CCL-2) and human hepatoblastoma liver cells (HepG2; ATCC HB-8065) were grown in RPMI 1640 (PAA, Pasching, Austria) supplemented with 10% (v/v) heat-inactivated fetal bovine serum (Biochrome, Berlin, Germany).21
Preparation of cytoplasmic and nuclear envelope extracts
Cytoplasmic and nuclear envelope cell extracts from HL-60, HeLa and HepG2 cells were prepared as described previously.12 22 The obtained nuclear envelope fraction was diluted 1:1 with sample buffer (250 mM Tris-HCl, pH 6.8; 4% (w/v) sodium dodecyl sulfate (SDS); 0.005% (w/v) bromphenol blue; 20% (v/v) glycerol; 5% (v/v) β-mercaptoethanol).
Preparation of tubulin extracts from HL-60, HeLa and HepG2 cells
Using repeated cycles of tubulin polymerisation and depolymerisation, we purified tubulin from HL-60 and HeLa cells, adopting standard protocols for the preparation of tubulin from bovine brain.23 The cells were pelleted (3000 rpm, 4°C), resuspended in ‘microtubuli buffer’ (0.1 M (2-N-morphilinoethanesulfonic acid), 1 mM EDTA, 0.5 mM MgCl2, pH 6.5) and cell membranes were broken up using a Dounce homogeniser. After centrifugation (13 000 rpm, 1 h, 4°C), equal proportions of cold ‘microtubuli buffer’ and 8 M glycerol (Sigma, St Louis, Missouri, USA) were added to the supernatant to induce depolymerisation of tubulin. For re-polymerisation, the isolated tubulin was incubated with 1 mM guanosine triphosphate (GTP) (Sigma) (30 min, 37°C) followed by another cycle of tubulin depolymerisation.
SDS–polyacrylamide gel electrophoresis (PAGE) and immunoblotting
One-dimensional (1D) SDS–PAGE was performed under reducing conditions according to standard protocols.24 Cytoplasmic, nuclear envelope or tubulin extracts from HL-60, HeLa and HepG2 cells were resuspended in sample buffer and electrophoretically resolved on SDS–polyacrylamide gels (10% separation gels, 4% stacking gels). The resolved proteins were stained with either Coomassie blue solution (0.05% (w/v) Coomassie brilliant blue R (Sigma), 50% (v/v) ethanol, 10% (v/v) acetic acid) or silver nitrate (Silver stain plus, Biorad, Hercules, California, USA).
Semi-dry western blotting was performed according to standard protocols.25 Reactive proteins were visualised by probing nitrocellulose membranes with ANCA-positive sera, prediluted 1:200–1:500 in blocking solution (5% (w/v) non-fat dry milk in PBS, 0.01% (v/v) Tween-20 (Sigma)). The resolved proteins were visualised by enhanced chemiluminescence (ECL Plus, Amersham Pharmacia Biotech).
Two-dimensional (2D) gel electrophoresis
2D gel electrophoresis was performed according to standard protocols.26 27
First dimension
Commercially available isoelectric focusing gels, containing immobilised pH gradients (11 cm, pH 4–7 and pH 4.7–5.9), were used (Ready Strips IPG, Biorad). After loading the samples onto the gels (diluted 1:1 in sample buffer containing 8 M urea, 2% 3-((3-cholamidopropyl)dimethylammonio)-1-propanesulfonate (CHAPS), 50 mM dithiothreitol (DTT), 0.2% Bio-Lyte ampholytes (Biorad)), the gels were passively rehydrated (12 h, room temperature) and submitted to isoelectric focusing (Protean IEF Cell, Biorad; 20 000 V h).
Second dimension
After isoelectric focusing, SDS–PAGE was performed as described above. Before loading the gel strips onto a 10% separation gel, the gels were equilibrated (equilibration buffer I: 6 M urea, 0.375 M Tris-HCl, pH 8.8, 2% (w/v) SDS, 20% (v/v) glycerol, 2% (w/v) DTT; equilibration buffer II: 6 M urea, 0.375 M Tris-HCl, pH 8.8, 2% (w/v) SDS, 20% (v/v) glycerol, 2.5% (w/v) iodoacetamide (Sigma), 10 min each). Proteins were resolved at 200 V for 45 min and detected with Coomassie blue or silver nitrate staining as described above.
Peptide mass fingerprinting
Peptide mass fingerprinting was independently performed at the Center for Molecular Medicine, University of Cologne, Germany, as well as at the SWISS-2D Service, Central Laboratory for Clinical Chemistry, University of Geneva, Switzerland using standard procedures for in-gel digestion and matrix-assisted laser desorption ionisation-time of flight mass spectrometry (MALDI-TOF-MS) of in-gel digested proteins.28 29 The proteins were identified from MALDI-TOF-MS fingerprints by searching either the NCBInr public database (National Center for Biotechnology Information, Bethesda, Maryland, USA), the SmartIdent database (Expert Protein Analysis System (Expasy), the proteomics server from the Swiss Institute of Bioinformatics, Basel, Switzerland) or the MASCOT search engine for rapid protein identification (version 1.9; Matrix science, London, UK). In this analysis, probability scores >45 were considered positive. Alignments of amino acid sequences were carried out using CLUSTAL W (version 1.81; Multiple Sequence Alignments Program of the Center for Molecular and Biomolecular Informatics, Nijmwegen, The Netherlands).
Cloning of myeloid cell-specific cDNA of TBB-5
To obtain myeloid cell-specific cDNA of TBB-5, total RNA was extracted from human promyelomonocytic HL-60 cells (RNAeasy, Qiagen, Hilden, Germany), polyadenylated mRNA was isolated (Omniscript, Qiagen) and applied for the synthesis of single-stranded cDNA by reverse transcription using oligo(dT) nucleotides according to the instructions of the manufacturer. Single-stranded cDNA was amplified by PCR using Taq polymerase (HotStarTaq DNA polymerase, Qiagen) and the oligonucleotides 5′-atgcgggagatcgtgcacc (forward primer) and 3′-ctaggccacctcctcctcc (reverse primer). Primer design was based on the nucleotide sequence of TBB-5 from human brain available in the NCBI GenBank (accession no. BC007605). After sequencing of the cDNA, the amplified cDNA was cloned into the eukaryotic expression vector pcDNA3.1/lacZ+ (Invitrogen, Carlsbad, California, USA) that introduced an Xpress sequence tag (-Asp-Leu-Tyr-Asp-Asp-Asp-Lys) into the N-terminus of the recombinant protein. The resulting plasmid was used for transient transfection of eukaryotic Cos-7 cells.
Transient transfection and recombinant expression of TBB-5
Transfection of eukaryotic monkey Cos-7 cells (ATCC CRL-1651) was performed using a lipofection technique following the instructions of the manufacturer (Biorad). The transfected cells were broken up by repeated freeze and thaw cycles (liquid nitrogen, 37°C) and the obtained pellet was used for further analysis by 1D or 2D gel electrophoresis and immunoblotting. To detect recombinant TBB-5, specific primary antibodies directed against the integrated sequence tag (eg, mouse-anti-Xpress 1:1000 (Santa Cruz, Biotechnology, Santa Cruz, California, USA)) and p-ANCA-positive sera or antibodies against distinct isotypes of β-tubulin were used. For control purposes, recombinant TBB-1, -2 and -4 with an integrated glutathione S-transferase (GST) tag were used (Abnova, Taiwan). Protein concentrations were applied according to the instructions of the manufacturer.
Recombinant expression of FtsZ
For recombinant expression of FtsZ, genomic DNA was extracted from Escherichia coli (strain M15) (MiniPrep, Qiagen) according to the instructions of the manufacturer. The cDNA was amplified using Taq polymerase (HotStarTaq DNA Polymerase, Qiagen) and the oligonucleotides 5′-cgccatggatgtttgaaccaatggaac (forward primer) and 3′-cgctcgagatcagcttgcttacgcagg (reverse primer), cleaned (QIAquick PCR Purification Kit, Qiagen) and subcloned into the prokaryotic/eukaryotic pQe-TriSystem expression vector (Qiagen) introducing a 6× histidine sequence tag at the N-terminus of the recombinant protein. For recombinant expression of FtsZ in E coli M15, standard procedures (Qiagen) were applied. Using Ni-NTA (nickel nitrilotriacetic acid) resins (Qiagen), 6×His-tagged FtsZ fusion proteins were eluted from the cell lysate in a pH-dependent manner (pH 4.5–5.3) and transferred into sample buffer for further analysis.
Affinity purification of p-ANCAs with human tubulin
Proteins of nuclear envelope extracts and tubulin preparations of HL-60 were resolved by SDS–PAGE30 transferred onto a nitrocellulose membrane using semi-dry western blotting, and ANCAs bound to tubulin were eluted from the membrane using 200 mM glycine (pH 2.8) and 1 mM EDTA for 20 s. The acid elution buffer was neutralised (pH 7.4) by adding 1 M Tris base. Randomly selected sera from eight patients (PSC n=3, AIH n=5, ANCA-negative controls n=2) containing p-ANCAs at high serum titres were used.
Preabsorption of sera containing p-ANCAs with tubulin extracts from HL-60 cells or with recombinant FtsZ
Sera from patients with AIH (n=10), PSC (n=10), ulcerative colitis (n=10) and disease control sera (n=4) were prediluted in PBS (1:20, 1:100, 1:500 and 1:1000) and then absorbed with tubulin extracts from HL-60 cells, bovine serum albumin (BSA)or FtsZ (20 μg/μl for each protein) at 37°C for 3 h. Finally, preabsorbed sera were analysed for serum end point titre by indirect immunofluorescence microscopy on human neutrophils.
Interleukin 10-deficient mice
Interleukin 10-deficient (Il10–/–) mice provide an established model for IBD which also leads to the generation of ANCAs. To investigate the effect of the intestinal microflora on the generation of antibodies against TBB-5 and FtsZ, 5- to 14-month-old Il10–/– mice (C3H/HeJBir n=9 or C57BL/6J n=10 genetic background) were raised either in plastic film isolators under germ-free conditions or in individually ventilated cages enabling normal microbial colonisation of the gut.31 Sera from these mice were investigated for the presence of ANCAs by immunofluorescence microscopy on ethanol-fixed human neutrophils as well as on immunoblots for reactivity with myeloid cell-specific tubulin extracts and recombinant FtsZ.
Statistics
Data are reported either as medians or as means±SEM, as indicated. For comparison of categorical data, Fisher exact test or Wilcoxon matched pairs rank test was used. p Values <0.05 were considered to be significant.
Results
Identification of TBB-5 as the antigen of p-ANCAs in autoimmune liver disorders using peptide mass fingerprinting
Our own previous data obtained by 1D and 2D gel electrophoresis pointed to an acidic myeloid cell-specific 50 kDa protein at the inner side of the nuclear envelope as the target antigen of p-ANCAs in AIH and PSC.12 The reactive protein band detected in 1D gel electrophoresis using nuclear HL-60 extracts actually comprised a complex mixture of various nuclear envelope proteins with the same molecular weight but different isoelectric points (pIs). To improve protein resolution, commercially available 2D gels with immobilised pH gradients were applied. Using sera positive for ANCAs from patients with autoimmune liver disorders, two reactive protein spots with an approximate molecular weight of 50 kDa and acidic pIs between 4.9 and 5.1 were detected on immunoblots (figure 1A,B). Reactivity with these two protein spots was found in 94% (32/34) of the tested ANCA-positive sera from patients with PSC (n=4/5) and AIH (n=28/29). The selected sera for 2D immunoblotting comprised a representative proportion of the entire study population. Importantly, none of the tested ANCA-negative control sera (n=5) recognised any of the two protein spots.

Peptide mass fingerprint analysis of nuclear envelope proteins recognised by perinuclear antineutrophil cytoplasmic antibodies (p-ANCAs). Extracts of nuclear envelopes from HL-60 cells were resolved by 2D gel electrophoresis. For isoelectric focusing, gel strips with immobilised pH gradients (range 4.7–5.9) were used. Among multiple protein spots detected on the 2D gel by silver nitrate staining (A), two reactive spots with an approximate molecular mass of 50 kDa and a pI value between 4.9 and 5.1 were identified by immunodetection (B). For immunoblotting, serum from a patient with autoimmune hepatitis and p-ANCAs (serum end point titre 1:2560) was used in the shown examples. Peptide mass fingerprint analysis (C) identified β-tubulin isotype 5 in each of the spots (TBB-5; SWISS-Prot accession number P07437; NCBI accession number BC007605).
MALDI-TOF-MS analysis of either reactive protein spot revealed a spectrum of peptides, which were assigned to the amino acid sequence of TBB-5 based on extraordinarily high probability scores (range 318–780, positive >45) calculated by the MASCOT search engine for rapid protein identification (SWISS-PROT accession number, PO7437; NCBI accession number, BC007605). Peptide mass fingerprint analysis allowed direct identification of 44% of the full-length amino acid sequence of TBB-5 (figure 1C). Identified peptides were clustered within the first 391 out of 444 amino acids of the TBB-5 amino acid sequence, whereas peptides located in the C-terminus of TBB-5 were not identified in any of the fingerprint analyses, irrespective of the involved mass spectrometry facility.
p-ANCAs in autoimmune liver disorders specifically recognise autoantigen in tubulin extracts
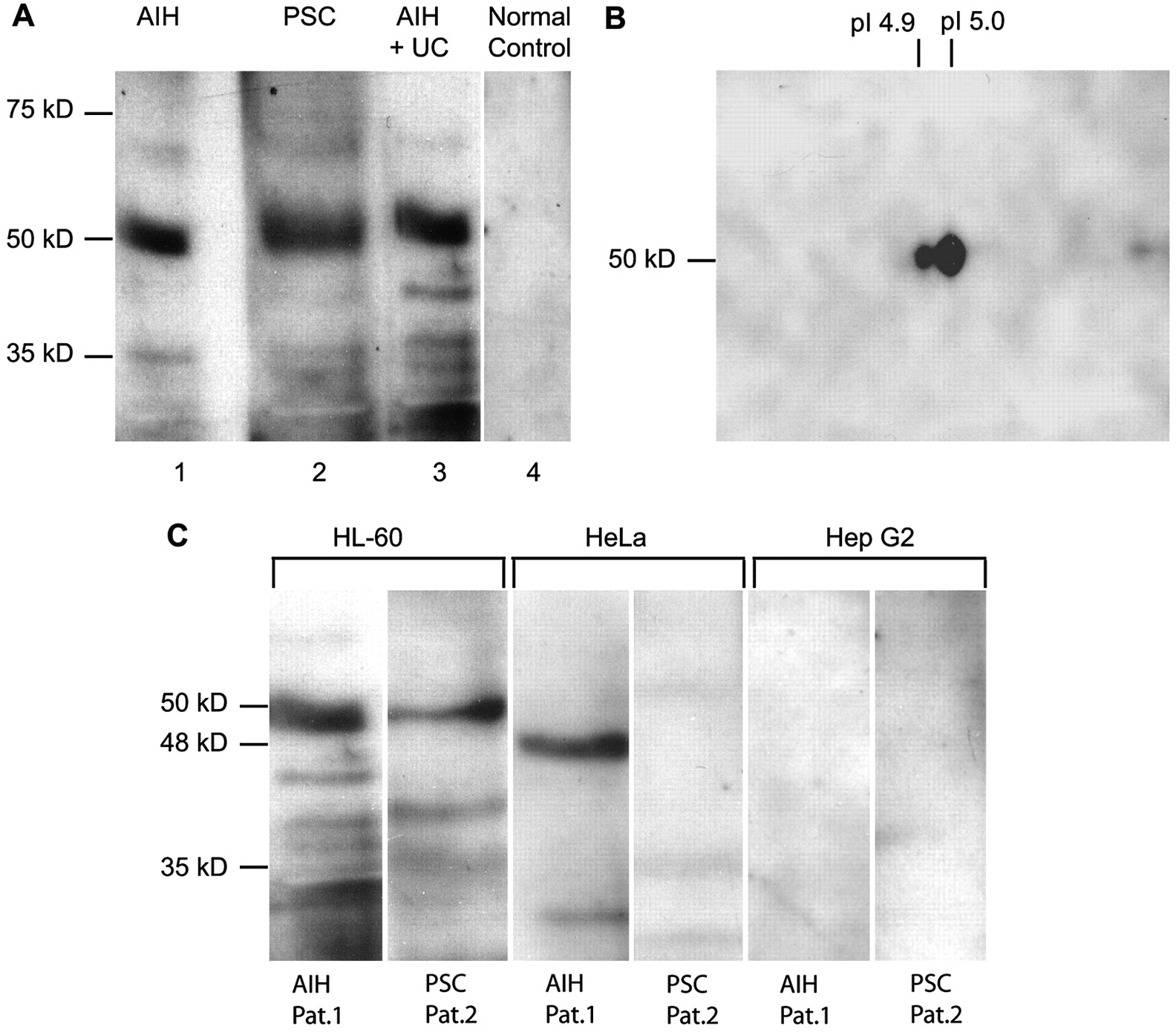
To confirm the identity of the TBB-5-related protein as the target antigen of p-ANCAs in autoimmune liver disorders, three different experimental approaches were used. First, purified tubulin extracts were prepared from myeloid cell-specific HL-60 cells using repeated depolymerisation/repolymerisation cycles of tubulin preparations.23 The obtained tubulin preparations comprised a mixture of different tubulins and tubulin-binding proteins. On 1D immunoblots of human tubulin extracts, 88% of the ANCA-positive sera from patients with AIH (84/95) as well as 72% of sera from patients with PSC (28/39) recognised a reactive protein band of 50 kDa approximate molecular mass, while none of the sera from healthy controls and only a few sera from disease controls (HCV 5% (1/19, serum ANCA positive), HBV 5% (1/22, serum ANCA positive), alcoholic liver cirrhosis 16% (7/43, 3/7 sera ANCA positive in indirect immunofluorescence)) reacted with the human tubulin preparations (figure 2A). In addition, immunoreactivity with human tubulin extracts was found in 58% (11/19) of the sera from patients with PSC without associated IBD, and in 70% (14/20) of the sera from patients with concomitant ulcerative colitis (69%) and Crohn's disease (71%) (p>0.05), respectively. None of these autoantibody reactivities was correlated to clinical aspects of PSC (disease activity, non-fibrotic stage and liver cirrhosis) or IBD (disease extent, intake of drugs and treatment response). Unlike autoimmune liver disorders, only 40% (12/30) of the sera of ANCA-positive patients with ulcerative colitis without associated PSC recognised human tubulin, and the presence of this antibody also appeared unrelated to disease activity or disease extent. Additional 2D gel electrophoresis of tubulin preparations gave rise to two reactive spots with an approximate molecular weight of 50 kDa and acidic pIs between 4.9 and 5.0 when immunoblots were probed with p-ANCA-positive sera from patients with autoimmune liver disorders (figure 2B). The spectrum of peptides obtained by MALDI-TOF-MS of the myeloid cell-specific human tubulin preparations confirmed TBB-5 as the antigen (probability scores 640–780).

Reactivity of perinuclear antineutrophil cytoplasmic antibodies (p-ANCAs) with myeloid cell-specific proteins from tubulin preparations. Tubulin extracts prepared from myeloid cell-specific HL-60 cells or non-myeloid cell lines (HeLa, HepG2) were resolved by either 1D or 2D gel electrophoresis. For isoelectric focusing, gel strips with immobilised pH gradients (range 4.7–5.9) were used. Tubulin proteins were visualised by immunodetection using sera from patients with autoimmune liver disorders positive for p-ANCAs. (A) The majority of the sera (97%; 33/34) which contained p-ANCAs recognised a tubulin protein with a molecular weight of 50 kDa in 1D gel electrophoresis (lane 1, autoimmune hepatitis (AIH), p-ANCAs 1:5120; lane 2, primary sclerosing cholangitis (PSC), p-ANCAs 1:320; lane 3, AIH and ulcerative colitis (UC), p-ANCAs 1:1280). Sera from normal healthy controls did not react with any of the resolved proteins (lane 4). (B) 2D immunoblots revealed two proteins with an approximate molecular mass of 50 kDa and pI values of 4.9–5.0 recognised by ANCA-positive sera from patients with autoimmune liver disorders. For immunoblotting, serum from a patient with AIH (p-ANCAs 1:5120) was used. Fingerprint analysis of the reactive protein spots again confirmed β-tubulin isotype 5 (TBB-5). (C) Specific reactivity of p-ANCAs in autoimmune liver disorders was directed against a 50 kDa tubulin protein from myeloid cells such as HL-60 cells. A small proportion of sera positive for p-ANCAs also recognised proteins with an approximate molecular mass of 48 kDa in tubulin preparations from non-myeloid HeLa cells. Peptide mass fingerprinting of the 48 kDa spot revealed a complex mixture of proteins not related to TBB-5. Lanes 1, 3, 5, ANCA-positive serum (1:1280) from a patient with AIH (patient 1); lanes 2, 4, 6, ANCA-positive serum (1:320) from a patient with PSC (patient 2). All sera were prediluted 1:200.
Secondly, to check cell type specificity of the putative target antigen we studied whether p-ANCA-positive sera from patients with autoimmune liver disorders reacted with tubulin preparations from epitheloid cells such as HeLa and HepG2 cells. Of note, only 6% and 15% of the sera containing p-ANCAs reacted with tubulin preparations from HepG2 cells (2/34) and HeLa cells (5/34), respectively. However, these reactive protein bands showed lower molecular masses (40–48 kDa) than proteins detected on immunoblots of tubulin preparations from myeloid HL-60 cells (figure 2C). MALDI-TOF-MS fingerprint analysis of the reactive proteins from HeLa and HepG2 cell preparations indicated a complex mixture of various proteins not related to TBB-5 (data not shown).
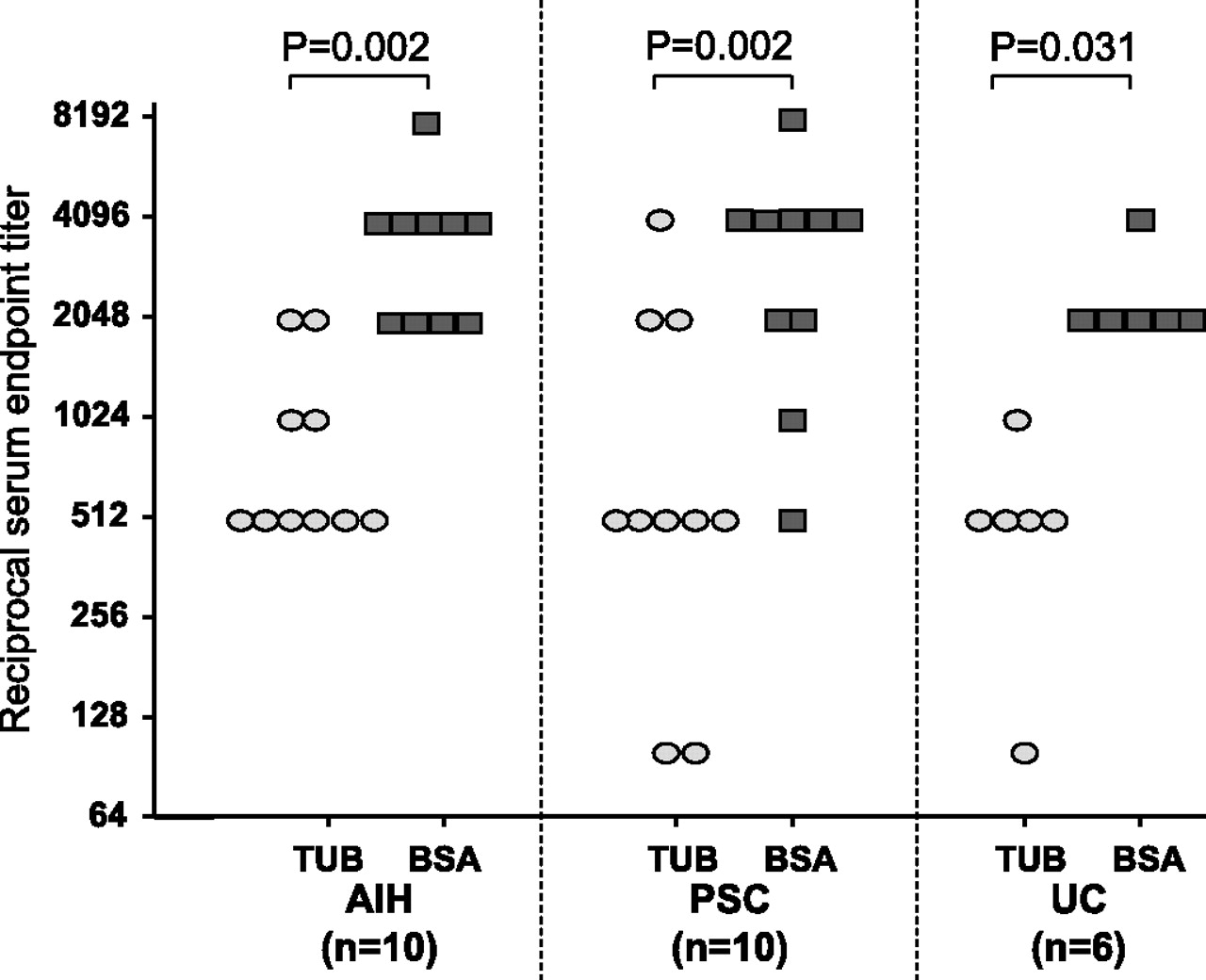
Finally, specific immunoglobulins in p-ANCA-positive sera were either affinity purified or preabsorbed against the reactive 50 kDa protein of tubulin extracts from HL-60 cells. Affinity-purified antibodies retained the characteristic p-ANCA staining pattern associated with autoimmune liver disorders, yielding a rim-like inhomogeneous staining of the nuclear periphery along with multiple intranuclear fluorescent foci on ethanol-fixed neutrophils (Supplementary figure 1A,B online). In contrast, the intensity of the ANCA-specific immunofluorescence pattern continously decreased after the sera had been preabsorbed with myeloid cell-specific tubulin extracts at increasing concentrations (Supplementary figure 1C,E), but remained unchanged after preabsorption of the sera with the control protein BSA (Supplementary figure 1F–H). Serum titres of atypical p-ANCAs after preabsorption with HL-60 tubulin were significantly lower compared with preabsorption of sera with BSA (figure 3).

Preabsorption of atypical perinuclear antineutrophil cytoplasmic antibodies (p-ANCAs) with tubulin extracts or bovine serum albumin (BSA)—comparison of serum titres after preabsorption. Preabsorption of sera with tubulin (TUB) preparations (20 μg/μl) gradually decreased the intensity of the ANCA immunofluorescence staining on human neutrophils, whereas the intensity of ANCA staining basically remained unaffected after preabsorption with BSA (20 μg/ml). The serum titres of ANCAs in sera from patients with autoimmune hepatitis (AIH), primary sclerosing cholangitis (PSC) or UC were significantly lower in each single patient after preabsorption with human tubulin as compared with BSA. For statistical analysis, Wilcoxon matched pairs rank test was performed.
Recombinant TBB-5 is recognised by ANCA-positive sera
To confirm TBB-5 as the target antigen of p-ANCAs in autoimmune liver disorders, recombinant TBB-5 with an integrated Xpress sequencing epitope was expressed in eukaryotic Cos-7 cells. Overexpression of recombinant TBB-5 was hampered by the fact that the gene products exerted toxic effects on Cos-7 cells, resulting in remarkable disintegrity of the microtubular network and rapid protein degradation. Thus, only comparatively low quantities of recombinant TBB-5 were available for 2D electrophoresis and immunoblotting. Using antibodies directed against the integrated Xpress epitope tag as well as sera positive for p-ANCAs, two reactive spots with molecular masses of 50 kDa and acidic pI (4.7–4.9) were detected in 97% of the p-ANCA-positive sera already investigated for reactivity on 2D immunoblot using human tubulin extracts (Supplementary figure 2A,B). Immunoblots of recombinant TBB-5 also revealed faint reactive spots at molecular masses of 48 kDa and pI of 4.0, indicating degradation of the recombinant protein. In control experiments using Cos-7 cells transfected with empty vector, ANCA-positive sera did not recognise any reactive proteins (Supplementary figure 2C). Furthermore, atypical p-ANCAs did recognise recombinant TBB-1, but not recombinant TBB-2 and TBB-4 using 1D immunoblots (Supplementary figure 3). Of note, the amino acid sequences of TBB-1 and TBB-5 are highly homologous and differ only in four amino acids; however, MALDI-TOF-MS analysis previously unequivocally identified TBB-5 as the responsible target antigen of ANCAs due to the presence of a unique characteristic peptide.
Cross-reactivity of p-ANCAs with TBB-5 and microbial protein FtsZ
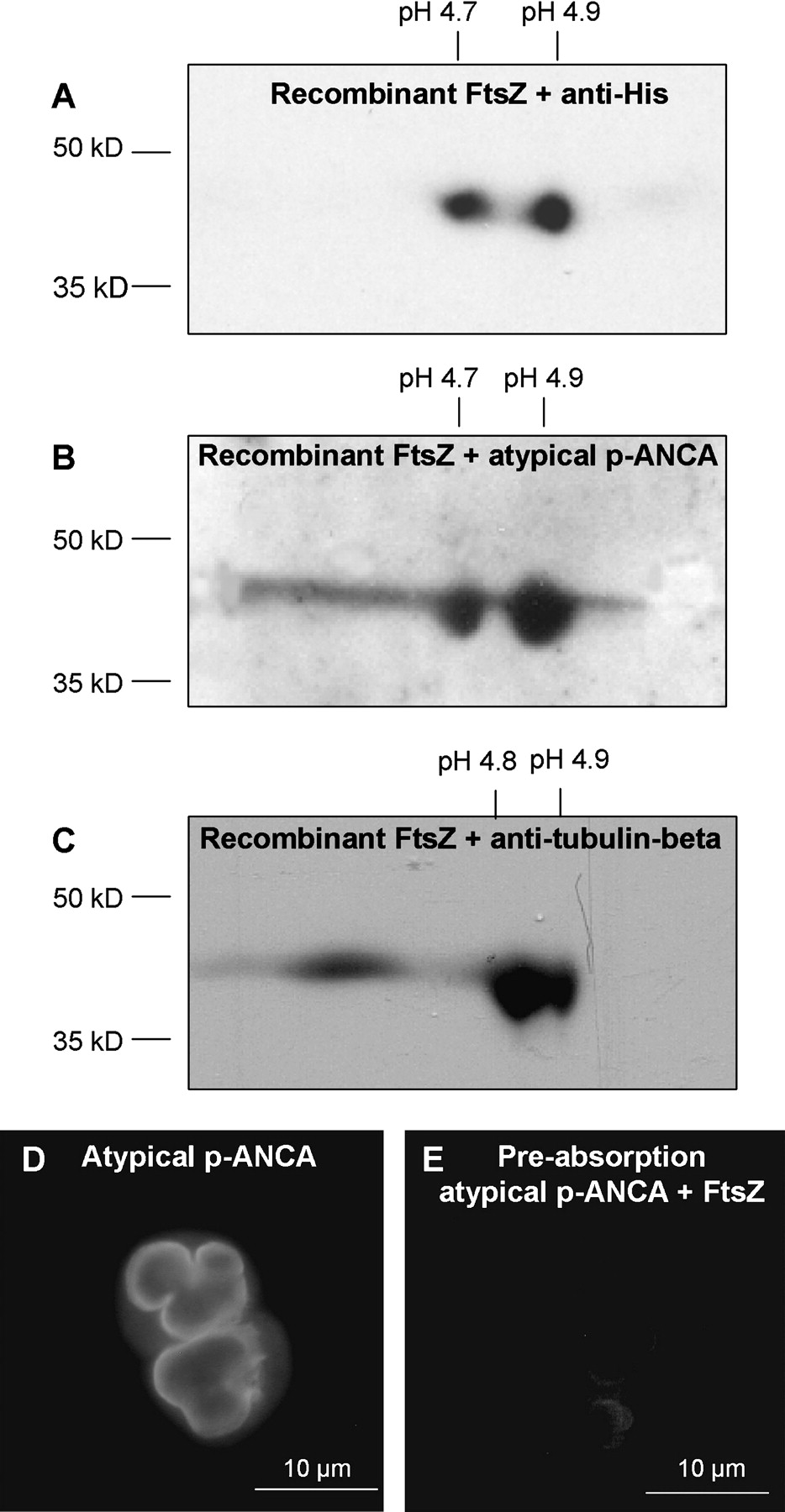
The microbial cell division protein FtsZ represents an evolutionary precursor protein of β-tubulin, which is present in almost all bacteria of the intestinal microflora. Although both proteins reveal amino acid sequence homology of <20%, they share an extraordinarily high structural homology particularly in the functionally important GTP-binding region due to evolutionarily conserved epitopes.16 Furthermore, 2D immunoblots of recombinant FtsZ using randomly selected sera from patients with autoimmune liver disorders revealed two reactive protein spots with an approximate molecular mass of 42 kDa and pIs between 4.7 and 4.9 (figures 4A–C). MALDI-TOF-MS analysis of either spot confirmed FtsZ as the reactive protein. The MASCOT search engine for rapid protein identification revealed high probability scores of 323 (positive >45; SwissProt accession number P0A9A6). In addition, sera from patients with autoimmune liver disorders lost their characteristic p-ANCA immunofluorescence staining pattern on human neutrophils when they had been preabsorbed with recombinant FtsZ (figure 4D,E).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Detection of recombinant FtsZ. Recombinant FtsZ overexpressed in E coli M15 was resolved by 2D gel electrophoresis followed by immunoblotting. (A) Using antibodies against the integrated polyhistidine sequence tag of recombinant FtsZ, two characteristic spots at a molecular mass of 42 kDa and pHs of 4.7 and 4.9 were detected. (B) Incubation of the immunoblots with sera containing perinuclear antineutrophil cytoplasmic antibodies (p-ANCAs) (1:1280) revealed the same reactive spots with an approximate molecular weight of 42 kDa and pHs at 4.7 and 4.9 as detectable with antibodies directed against the polyhistidine sequence tag. (C) Commercially available antibodies against β-tubulin also recognised the aforementioned reactive spots. Mass spectrometry analysis confirmed these reactive spots as FtsZ. (D) Sera containing p-ANCAs at high serum end point titres (1:2560) were preabsorbed with recombinant FtsZ. After preabsorption, the characteristic fluorescence pattern of p-ANCAs was no longer detectable (E).
These initial findings prompted us to study the role of the intestinal microflora for generation of antibodies against TBB-5 and FtsZ. Thus, we first also investigated sera from Il10–/– mice. These mice developed an IBD-like syndrome when housed in individually ventilated cages enabling normal microbial colonisation of their guts. In contrast, all Il10–/– mice remained healthy when grown under germ-free conditions. Of note, 80% of the sera from Il10–/– (8/10) mice with normal gut colonisation contained antibodies that revealed the characteristic fluorescence pattern of p-ANCAs on ethanol-fixed neutrophils in indirect immunofluorescence microscopy. The same sera also reacted with recombinant FtsZ and human tubulin preparations on immunoblots. This dual reactivity against tubulin and FtsZ was found in Il10–/– mice strains irrespective of differences in genetic backgrounds. In contrast, none of the sera from Il10–/– mice grown under germ-free conditions (n=9) revealed any specific immunofluorescence staining on human neutrophils or immunoreactivity with recombinant FtsZ or human tubulin preparations (table 2).
Distinct reactivity of sera from Il10–/– mice mice grown under germ-free conditions or in individually ventilated cages with electrophoretically resolved human tubulin preparations or recombinant FtsZ
Reactivity of sera with human tubulin preparations from HL-60 cells revealed a 50 kDa protein on immunoblots; sera recognising recombinant FtsZ showed a 42 kDa protein band on immunoblots. Sera from Il10–/– mice containing ANCAs yielded a perinuclear immunofluorescence staining of the nuclear periphery along with intranuclear fluorescent foci as described for human p-ANCAs in sera from patients with autoimmune liver disorders.
Finally, we evaluated a large panel of sera to determine whether ANCA-positive sera from patients with autoimmune liver disorders, sera from patients with non-autoimmune liver diseases (HBV, HCV, alcoholic liver cirrhosis) and ANCA-negative sera from healthy controls recognised FtsZ and/or tubulin (table 3). As expected, sera from patients with autoimmune liver disorders were reactive with both proteins, recombinant FtsZ (AIH 82% (75/95), PSC 64% (25/39)) and tubulin (AIH 88% (84/95), PSC 72% (28/39). Of note, dual reactivity against tubulin-β and FtsZ was present in all these sera. Subgroup analysis of patients with PSC revealed that concomitant IBD, in particular ulcerative colitis, slightly increased the prevalence of sera reacting with FtsZ (PSC without IBD 58% (11/19), PSC with IBD 70% (14/20)). Unexpectedly, ANCA-negative sera from patients with non-autoimmune liver disorders also recognised the recombinant FtsZ protein at particularly high frequencies (HBV 91%, HCV 84%, alcoholic liver cirrhosis 95%), whereas FtsZ antibodies were less frequently detectable in healthy controls (16% (6/37)). Nevertheless, dual reactivity to tubulin proteins as well as to FtsZ in these disorders represented an infrequent finding (HBV 5%, HCV 5%, alcoholic liver cirrhosis 16% and healthy controls 5%). Of note, reactivity to TBB-5 without the presence of FtsZ antibodies was found in a very few cases (<1%) of all sera studied.
Reactivity of atypical p-ANCAs with TBB-5 and/or FtsZ in autoimmune liver disorders, liver disease controls and healthy controls
Discussion
The high prevalence of p-ANCAs in sera from patients with autoimmune liver disorders generated ambitious efforts to identify the target antigen. Several cytoplasmic proteins have been proposed as putative autoantigens such as bactericidal/permeability-increasing protein, catalase, cathepsin G, enolase or lactoferrin. Thus far, all candidate autoantigens, however, reacted with <35% of the ANCA-positive sera.13 32–36 Recent studies have provided circumstantial evidence for a myeloid cell-specific 50 kDa nuclear envelope protein as the ANCA autoantigen.9 11 12 Here, we demonstrate the molecular identity of the 50 kDa protein as TBB-5, a member of the β-tubulin family. The close relationship of the p-ANCA autoantigen to TBB-5 is supported by MALDI-TOF-MS fingerprint analysis that revealed exceedingly high probability scores for TBB-5, which unequivocally allowed us to distinguish the 50 kDa protein from other known highly homologous members of the β-tubulin family, such as TBB-1. In addition, antibodies directed against TBB-1 to TBB-4 did no show relevant reactivity with recombinant TBB-5, whereas antibodies against TBB-1 revealed the same reactive spots as ANCA-positive sera. This observation reflects the extraordinarily high amino acid sequence homology between TBB-5 and TBB-1, only differening in a unique peptide identified in our MS analyses.
Our study provides further evidence to support TBB-5 as the p-ANCA antigen in autoimmune liver disorders. For instance, tubulin preparations from myeloid-specific HL-60 cells, but not from non-myeloid epithelial HepG2 or HeLa cells, showed reactivity with p-ANCA-positive sera from patients with autoimmune liver diseases. Of note, ANCA-positive sera which had been preabsorbed with tubulin preparations from HL-60 cells lost their characteristic p-ANCA fluorescence pattern on human neutrophils. On the other hand, when antibodies bound to myeloid cell-specific tubulin were eluted from immunoblots, the specific immunofluorescence pattern associated with p-ANCAs in autoimmune liver disorders was still detectable. Importantly, the majority of p-ANCA-positive sera (97%) from patients with AIH or PSC reacted with recombinant TBB-5 expressed in Cos-7 cells, which conspicuously contrasts TBB-5 to all other previously reported candidate antigens of p-ANCAs.
β-Tubulins along with α-tubulin form the structural subunit of microtubules, which are part of the cytoskeleton in virtually every cell. Thus, identification of TBB-5 as the target antigen of p-ANCAs in autoimmune liver disorders was an unexpected finding. However, TBB-5 may qualify as the autoantigen of atypical p-ANCAs for two major reasons. First, β-tubulins comprise several distinct isotypes, which show a selective specific expression pattern depending on species, cell type and developmental stage.37–43 Secondly, the TBB-5 autoantigen is quite probably present in a novel non-microtubular form, which might be particularly accessible to autoantibodies in haematopoetic cells.
The complexity of tubulin isotypes is further increased owing to their extensive post-translational modifications such as phosphorylation, acetylation or glutamylation, which can lead to secondary or tertiary protein structures peculiar to certain tubulin isotypes.44 45 The presence of such post-translational modifications may explain why the same TBB-5 amino acid sequence was obtained when the two reactive spots with identical molecular masses but different pIs on the 2D immunoblots were analysed by MS in this study. However, to date, our attempts to identify these secondary modifications have remained inconclusive. Nevertheless, selective sequence analysis by MALDI-TOF-MS of each separate spot has confirmed the same TBB-5 sequence. Differential expression of human TBB-5 has not been characterised in great detail yet. However, the highly conserved murine TBB-5 homologue is ubiquitously expressed during embryonic development and in newborn mice, but shows limited expression in adult animals, mainly in brain, thymus and spleen.40 In this context, atypical p-ANCAs did not recognise tubulin preparations from other tissues—for example, bovine brain (data not shown).
Although tubulins constitute the principal components of microtubules, which are necessary to move around subcellular structures such as chromosomes, mitotic spindles and other organelles inside the cell, recent data demonstrated that distinct isotypes of tubulin can be detected in nuclei of non-dividing cells, particularly in association with the heterochromatin.46–48 In these studies, disruption of microtubuli by colchemide treatment or low temperature did not abolish the nuclear binding of tubulin antibodies, indicating that nuclear forms of tubulin exist which were present in a soluble non-polymerised state. In support of a nuclear form of tubulin, Kourmouli and co-workers demonstrated that binding of heterochromatin protein 1 to the nuclear envelope is regulated by a soluble form of tubulin.48 These non-microtubular nuclear forms of TBB-5 which are particularly accessible in distinct cell types may be a second reason why TBB-5 is selectively recognised as an autoantigen in myeloid cells, providing a myeloid cell-specific staining of the nuclear periphery in immunofluorescence microscopy. Unfortunately, we have not been able to study further the functional role of TBB-5 in stably transfected cells thus far because overexpressed TBB-5 induced extraordinary disintegrity of the microtubular network resulting in early protein degradation and cell death.49
β-Tubulins belong to a multigene family of >800 members, also comprising microbial proteins such as the bacterial protein FtsZ.50 FtsZ is expressed in almost all bacteria of the intestinal microflora during cell division and is considered to represent an evolutionary precursor protein of β-tubulins. In particular, both TBB-5 and FtsZ share an extraordinarily high degree of structural homology in the area of the evolutionarily conserved epitopes, such as the functionally essential GTP-binding region. Thus, to check for a possible link between immune responses against bacterial antigens and the generation of ANCAs, we investigated sera from Il10–/– mice. These knock-out mice represent a well-characterised animal model to study the role of intestinal bacteria for chronic IBD, because Il10–/– mice exposed to a normal microbial environment develop an ulcerative colitis-like syndrome and ANCAs in their sera. In contrast, Il10–/– animals remain healthy when housed under germ-free conditions. Cross-reactivity between ANCAs and bacterial antigens was supported by the observation that only Il10–/– mice with normal microbial gut colonisation, but not disease-free animals kept under germ-free conditions, developed antibodies with dual reactivity against recombinant FtsZ from E coli and human TBB-5. Thus, exposure to the gut flora represents an essential prerequisite for ANCA generation and points to a pivotal role for abnormal immune responses to enteric bacterial antigens in the immunopathogenesis of AIH and PSC.51–55
However, when the reactivity of p-ANCAs with FtsZ and TBB-5 was evaluated in a large panel of human sera also comprising disease controls, several unexpected observations were obtained. In contrast to autoimmune liver disorders, p-ANCAs in patients with ulcerative colitis without associated PSC recognised TBB-5 in a comparatively low percentage of cases (40%). In this context, it has been a longstanding observation in earlier immunofluorescence studies that p-ANCAs in autoimmune liver disorders differ in their staining characteristics from those in IBDs, for example additional staining of monocytes in autoimmune liver disorders (reviewed in Semrad et al56.) Thus, other autoantigens seem to be involved in pure IBDs compared with autoimmune liver disorders. Even more puzzling was the observation that a high proportion of disease controls also reacted with FtsZ, indicating that antibodies to this protein most probably represent a non-specific frequent finding. Nevertheless, almost all ANCA-positive sera from patients with autoimmune liver disorders—irrespective of associated IBD—showed dual reactivity to both proteins TBB-5 and FtsZ, whereas most sera from the non-autoimmune disease controls exclusively reacted with FtsZ, but not TBB-5. This divergent antibody reactivity pattern between patients with autoimmune versus non-autoimmune liver disorders may reflect individual susceptibility present in subjects predisposed to autoimmunity.
Taken together, p-ANCAs in autoimmune liver diseases are directed against human TBB-5 cross-reacting with the bacterial protein FtsZ. Dual antigen reactivity to both FtsZ and TBB-5 probably reflects an abnormal immune response to intestinal microorganisms in susceptible, possibly genetically predisposed individuals.
Acknowledgments
We gratefully acknowledge valuable discussions with Howard J Worman, Departments of Medicine and of Anatomy and Cell Biology, Columbia University, New York.
References
Supplementary materials
Web only app
Footnotes
Linked articles 203091.
Funding Lise-Meitner-Foundation, Germany, and Bonfor Foundation, University of Bonn, Germany.
Competing interests None.
Patient consent Obtained.
Ethics approval This study was conducted with the approval of the University of Bonn ethics committee.
Provenance and peer review Not commissioned; externally peer reviewed.
Patent application An international patent application (PCT/EP05/54104) was filed.