Article Text

Abstract
Background and aims: The normal gastrointestinal microflora exerts a barrier effect against enteropathogens. The aim of this study was to examine whether lactobacilli, a minor genus of the resident gut microflora, exerts a protective effect against the cellular injuries promoted by the diarrhoeagenic Afa/Dr diffusely adhering Escherichia coli (Afa/Dr DAEC) C1845 strain in human intestinal cells.
Methods: Cultured human intestinal fully differentiated enterocyte-like Caco-2/TC7 cells were used. Antibacterial activity was examined by measuring the viability of the adhering C1845 bacteria. The distribution of brush border associated cytoskeleton and functional proteins was examined by immunofluorescence labelling coupled to confocal laser scanning microscopy analysis.
Results: The activity of Lactobacillus acidophilus strain LB isolated from the resident human gastrointestinal microflora was examined. A dose dependent decrease in viability of C1845 bacteria was observed after both direct contact in vitro between the spent culture supernatant (LB-SCS) and the bacteria, and when the bacteria were adherent on Caco-2/TC7 cells. Protection against the C1845 induced alterations in expression of F-actin, sucrase-isomaltase, dipeptidylpeptidase IV, alkaline phosphatase, and fructose transporter alterations was observed when the cells were exposed to LB-SCS.
Conclusion:L acidophilus strain isolated from the resident adult human gastrointestinal microflora, together with its antimicrobial activity, exerts a protective effect against the brush border lesions promoted by the diarrhoeagenic Afa/Dr DAEC strain C1845.
- intestinal cells
- brush border lesions
- gastrointestinal function
- lactobacillus
- antimicrobial
- diarrhoea
- MV, microvillus
- SI, sucrase-isomaltase
- DPP IV, dipeptidylpeptidase IV
- GPI, glycosylphosphatidylinositol
- AP, alkaline phosphatase
- TRITC, tetramethyl rhodamine isothiocyanate
- FITC, fluorescein isothiocyanate
- MRS broth, De Man, Rogosa, Sharpe broth
- SCS, spent culture supernatant
- TSA, tryptic soy agar
- PBS, phosphate buffered saline
- LDH, lactate dehydrogenase
- cfu, colony forming units
- CLSM, confocal laser scanning microscope
- DMEM, Dulbecco's minimal essential medium
Statistics from Altmetric.com
- MV, microvillus
- SI, sucrase-isomaltase
- DPP IV, dipeptidylpeptidase IV
- GPI, glycosylphosphatidylinositol
- AP, alkaline phosphatase
- TRITC, tetramethyl rhodamine isothiocyanate
- FITC, fluorescein isothiocyanate
- MRS broth, De Man, Rogosa, Sharpe broth
- SCS, spent culture supernatant
- TSA, tryptic soy agar
- PBS, phosphate buffered saline
- LDH, lactate dehydrogenase
- cfu, colony forming units
- CLSM, confocal laser scanning microscope
- DMEM, Dulbecco's minimal essential medium
Knowledge of the predominant genera and species of the gastrointestinal microflora as well as determination of their levels and biochemical activities are essential in the understanding of the microbial ecology of the gastrointestinal tract.1 The normal resident gastrointestinal microflora contains many diverse populations of bacteria which play an essential role in the development and well being of the host. Bacteroides thetaiotaomicron, a member of the intestinal microflora of mice and humans, modulates expression of genes involved in several important intestinal functions in colonised germ free mice.2 Moreover, this commensal bacterium changes a specific glycosylation process in cultured human intestinal cells.3 During the past 10 years, evidence has accumulated in support of a non-immune system of defence in the intestine. By continual release of antibiotic proteins, specialised cells of the intestinal epithelium may influence the extracellular environment and contribute to mucosal barrier function.4,5 In addition to the host cell non-immune system of defence, bacteria of the resident gut microflora exert a barrier effect against pathogens. We have recently reported that Escherichia coli, one of the first bacterial genera that colonise the intestine of humans, display antimicrobial activity against salmonella infection.6 Moreover, we have reported that other species of the endogenous human microflora such as bifidobacteria,7,8 a major species of the colonic microflora, and lactobacilli,9–14 a minor species of the gut microflora, exert antimicrobial activity by producing secreted antimicrobial substances.
To gain further insight into the mechanism by which resident bacteria of the human microflora could exert a protective role against enterovirulent pathogen induced cellular damage, we have chosen to examine the activity of a Lactobacillus acidophilus strain isolated from the resident adult human microflora for which the production of secreted antimicrobial substances has been well established both in vitro and in vivo.11–14 As an enterovirulent pathogen producing cellular damage in enterocytes, we have chosen the diarrhoeagenic wild-type Afa/Dr diffusely adhering E coli (DAEC) C1845 strain. Epidemiological studies have established a link between Afa/Dr DAEC and persistent diarrhoeal diseases, mostly in infants older than 24 months.15–18 Colonisation of the intestine by these pathogenic E coli is mediated by their harboured Afa/Dr adhesins. All of these adhesins mediate diffuse adhesion onto human epithelial intestinal cells and recognise as a receptor the decay accelerating factor (or CD55) by interacting with the short consensus repeat 2 and 3 domains of the molecule.19 We have recently demonstrated that after attachment onto intestinal brush border some members of this family of E coli promoted microvillus (MV) injury.20 These MV lesions resulted from apical cytoskeleton disassembly that follows activation of the CD55 molecule by Afa/Dr adhesins and subsequent Ca2+ dependent signalling.21 These structural lesions were accompanied by a dramatic alteration in the brush border expression of functional intestinal proteins such as hydrolases and transporters that play a pivotal role in intestinal function.22 As a cellular model, we have chosen the human colon adenocarcinoma cell line Caco-2,23 one of the human intestinal models most commonly used to study in vitro expression of intestine specific functional proteins.24 We have focused our study on the structural brush border associated protein F-actin, and brush border associated functional proteins, namely the hydrolases sucrase-isomaltase (SI) (EC 3.2.1.48–10) and dipeptidylpeptidase IV (DPP IV) (EC 3.4.14.5), the glycosylphosphatidylinositol (GPI) anchored protein alkaline phosphatase (AP), and the fructose transporter GLUT5.
MATERIALS AND METHODS
Reagents and antibodies
Fluorescein labelled phalloidin was from Molecular Probes (Junction City, Oregon, USA). BAPTA/AM (1,2-bis[2-amino-phenoxy]ethane-N,N,N`,N`-tetraacetic acid tetrakis [acetoxymethyl] ester) was obtained from Sigma-Aldrich Chimie SARL (L'Isle d'Abeau Chesnes, France).
Polyclonal rabbit anti-Dr adhesin antibody was a gift from BJ Nowicki (Texas University, Galveston, Texas, USA). The monoclonal antibodies antihuman SI (8A9) and anti-DPP IV (4H3) were a gift from S Maroux (ESA 6033 CNRS, Marseille, France). The rabbit polyclonal antibody directed against the fructose transporter GLUT 5 was kindly provided by E Brot-Laroche (INSERM U505, Paris, France). The rabbit polyclonal antibody directed against AP was from Dako (Tebu, Paris, France). The appropriate tetramethyl rhodamine isothiocyanate (TRITC) conjugated and fluorescein isothiocyanate (FITC) conjugated secondary antibodies were obtained from Boehringer and Immunoresearch ICN Laboratory and were diluted 1:20 to 1:200 in 0.2% gelatin-phosphate buffered saline (PBS).
Bacterial strains and growth conditions
The clinical isolate Afa/Dr DAEC C1845 strain harbouring the fimbrial F184525 was grown at 37°C for 18 hours in Luria broth.
L acidophilus LB strain was isolated from a human stool (Lacteol Laboratory, Houdan, France).11–14 LB bacteria were grown in De Man, Rogosa, Sharpe (MRS) broth (Biokar Diagnostic, Beauvais, France) for 18 hours at 37°C. Spent culture supernatant of LB (LB-SCS) was obtained by centrifugation at 10 000 g at 4°C for 30 minutes. Centrifuged LB-SCS was passed through a sterile 0.22 μm filter unit (Millex GS; Millipore, Molsheim, France). Filtered LB-SCS was assessed for the absence of bacterial colonies by plating on tryptic soy agar (TSA). A concentrated LB-SCS was obtained by freeze drying.
Cell culture
The low glucose consuming Caco-2/TC7 clone (Caco-2/TC7),26 established from the parental Caco-2 cell line,23 was used. This clone was similar to parental Caco-2 cells in terms of cell polarity and displayed an apical brush border endowed with functional proteins.26,27 Cells were routinely grown in Dulbecco's modified Eagle's minimal essential medium (DMEM) (25 mM glucose) (Life Technologies, Cergy, France), supplemented with 15% heat inactivated (30 minutes, 56°C) fetal calf serum (Life Technologies) and 1% non-essential amino acids (Life Technologies), as previously described.11–13 For maintenance purposes, cells were passaged weekly, using 0.02% trypsin in Ca2+-Mg2+ free PBS containing 3 mM EDTA. Experiments and maintenance of cells were carried out at 37°C in a 10% CO2/90% air atmosphere. The culture medium was changed daily. For assays of DAEC C1845 infection, Caco-2/TC7 cells were used at post-confluence after 15 days of culture (that is, they were fully differentiated cells).
Cell infection
The Caco-2/TC-7 monolayers were prepared in 24 well TPP tissue culture plates (ATGC, Paris, France). A cell infection assay was conducted as previously reported.20–22 Briefly, prior to infection, the cell monolayers were washed twice with PBS. C1845 bacteria suspended in the culture medium were added to each well of the tissue culture plate. The infection assay was conducted in the presence of 1% mannose to prevent type 1 fimbriae mediated binding. The plates were incubated at 37°C in 10% CO2/90% air for three hours and washed three times with sterile PBS.
Electron microscopy
For transmission electron microscopy, cells were rinsed three times with PBS, and fixed with 2.5% glutaraldehyde in 0.1 M sodium phosphate buffer (pH 7.4) for 30 minutes at room temperature. After washing with PBS, cells were postfixed for 30 minutes at room temperature with 1.5% osmium tetroxide in sodium phosphate buffer. Filters were then dehydrated in graded ethanol series, cut into strips, and embedded in epoxy resin. Ultrathin sections were cut from transversely orientated confluent monolayers. Ultrathin sections were double stained with uranyl acetate and lead citrate and examined with a Jeol JEM-1010 electron microscope.
Measurement of cell integrity
Cell integrity was determined by measuring lactate dehydrogenase (LDH) activity in the apical compartment of filter grown cells (Enzyline LDH kit; Biomérieux, Dardilly, France) according to the manufacturer's instructions.
Determination of antimicrobial activity
Antimicrobial activity of LB-SCS in vitro was examined as previously described.13,14 As indicator strain, the Afa/Dr DAEC C1845 strain was grown overnight for 18 hours at 37°C in Luria broth. Bacteria were centrifuged at 5500 g for five minutes at 4°C, washed once with PBS, and resuspended in PBS. C1845 bacteria were counted and a volume containing 108 colony forming units (cfu)/ml was used to determine the activity of LB-SCS. Colony count assays were performed by incubating 1 ml of PBS containing C1845 bacteria (108 cfu/ml) with 1 ml of LB-SCS at 37°C. At predetermined intervals, aliquots were removed, serially diluted, and plated on TSA to determine bacterial colony counts numbers.
In a second protocol, activity was determined in Caco-2/TC7 cells (108 cfu/ml) infected for three hours with C1845 bacteria. Infected cell monolayers were washed twice with sterile PBS to remove non-adhering bacteria and subjected to LB-SCS treatment for one hour at 37°C. To determine the viable cell associated C1845 bacteria, the infected cell monolayers were lysed by adding H2O. Appropriate dilutions were plated on TSA to determine the number of viable cell associated bacteria by bacterial colony counts.
Immunofluorescence
Monolayers of Caco-2/TC7 cells were prepared on glass coverslips which were placed in 24 well TPP tissue culture plates. After infection, cell preparations were fixed for 15 minutes at room temperature in PBS-3% paraformaldehyde. They were washed three times with PBS and then treated with 50 mM NH4Cl for 10 minutes for aldehyde function saturation.
Examination of F-actin was conducted by direct immunofluorescence labelling using fluorescein labelled phalloidin, as previously described.20–22 The coverslips were incubated for four minutes with PBS-0.2% Triton X-100 before incubation with fluorescein labelled phalloidin for 45 minutes at 22°C, after which the coverslips were washed three times with PBS.
Indirect immunolabelling of brush border associated SI, DPP IV, and AP was conducted with or without cell permeabilisation, as previously described.22 The monoclonal antibodies anti-SI 8A9 and anti-DPP IV 4H3 were diluted 1:200 and 1:50, respectively, in 0.2% gelatin-PBS. Polyclonal anti-AP antibody was diluted 1:20 in 0.2% gelatin-PBS. GLUT5 immunolabelling was conducted in Triton X-100 (0.2%) permeabilised cells.22 Polyclonal anti-GLUT5 was diluted 1:500 in 0.2% gelatin-PBS. Fixed monolayers were incubated with the primary antibody for 45 minutes at room temperature. After three washes in PBS, incubation with an FITC or TRITC conjugated secondary antibody (diluted 1:50 in PBS) was performed for 45 minutes at room temperature. After three washes, cells were mounted with Vectashield (Biosys SA, Compiégne, France) for immunofluorescence examination. No fluorescent staining was observed when the primary antibody was omitted.
For examination of adhering C1845 bacteria onto Caco-2/TC7 cells, indirect immunofluorescence microscopy was performed with a polyclonal antibody directed against Dr adhesin. Coverslips were incubated with primary antibody (diluted 1:100 in 0.2% gelatin-PBS) for 45 minutes at 22°C, washed three times with PBS, and then incubated with an FITC secondary antibody (diluted 1:50 in PBS) for 45 minutes at 22°C. The coverslips were then washed three times with PBS.
Specimens were mounted in Vectashield mounting medium (Vector Laboratory, Burlingame, California, USA). Specimens were examined by conventional epifluorescence microscopy using a Leitz Aristoplan microscope coupled to an Image Analyser Visiolab 1000 (Biocom, Les Ullis, France). Relative immunofluorescence intensity was measured and results expressed as arbitrary units. A confocal analysis was conducted using a confocal laser scanning microscope (CLSM) (model LSM 510 Zeiss, equipped with an air cooled argon ion laser 488 nm, and a helium neon laser 543 nm) configured with an Axiovert 100M microscope using a Plan Apochromat 63X/1.40 oil objective. Optical sectioning was used to collect 50 en face images 1 μm apart. Lateral views were obtained by integration of the images gathered at a step position of 1 on the x–y axis using the accompanying Zeiss software LSM510 2.5 on Windows NT4. Photographic images were resized, organised, and labelled using Adobe Photoshop software (San Jose, California, USA). The printed images (Kodak XLS 8600 PS, Eastman Kodak Co., Rochester, New York, USA) are representative of the original data. All photographs were taken on Kodak Electronic Imaging Paper (Eastman Kodak Co.).
Inhibitory assay
Inhibition of C1845 induced cell injury by LB-SCS was determined by infecting the Caco-2/TC7 cells with the C1845 bacteria (108 cfu/ml) at 37°C in 10% CO2/90% air for three hours with or without LB-SCS.
As a control, we used the cell permeable Ca2+ chelator BAPTA/AM which prevents C1845 induced F-actin cytoskeleton rearrangements.21,22 BAPTA/AM in methanol-dimethylformamide (50–50 v/v) was added to the culture medium 60 minutes before infection. The calcium chelator was maintained during the infection time course (three hours). The concentration used did not affect cell integrity, as measured by LDH release (control cells: 36 (5) U/l; H2O lysed cells: 2995 (25) U/l; BAPTA/AM treated cells: 39 (6) U/l ). Moreover, there was no change in C1845 binding (C1845 infected cells: 7.26 (0.14) log cfu/ml; C1845 infected BAPTA/AM treated cells: 7.29 (0.28) log cfu/ml) nor was expression of the apical F-actin network affected (control uninfected cells: 2.80 (0.18) immunofluorescence intensity arbitrary units; uninfected BAPTA/AM treated cells: 2.90 (0.25) arbitrary units).
Data analysis
Results are expressed as mean (SEM). For statistical comparisons, the Student's t test was performed.
RESULTS
Antibacterial activity against the Afa/Dr DAEC strain C1845
To determine whether the L acidophilus strain LB isolated from the human gut microflora exerted activity against a diarrhoeagenic pathogen, we focused our study on the SCS in which antibacterial compounds are present.11–14 The antibacterial activity of LB-SCS against the viability of Afa/Dr DAEC strain C1845 was examined in vitro after direct contact between the C1845 bacteria and LB-SCS. Examined as a function of LB-SCS concentration after three hours of contact, dose dependent antibacterial activity was observed (table 1). Highly significant activity was observed at a 2.0-fold concentrate of LB-SCS which induced a decrease of 3 logs in C1845 viability whereas MRS was almost inactive at this concentration. A highly significant 5 logs decrease in C1845 viability was observed with a 2.5-fold concentrate of LB-SCS whereas only a 2 logs decrease was found for the same MRS concentration.
Effect of LB-SCS and gentamicin treatments on the viability of C1845 bacteria.
Killing of C1845 bacteria adherent on human enterocyte-like Caco-2/TC7 cells
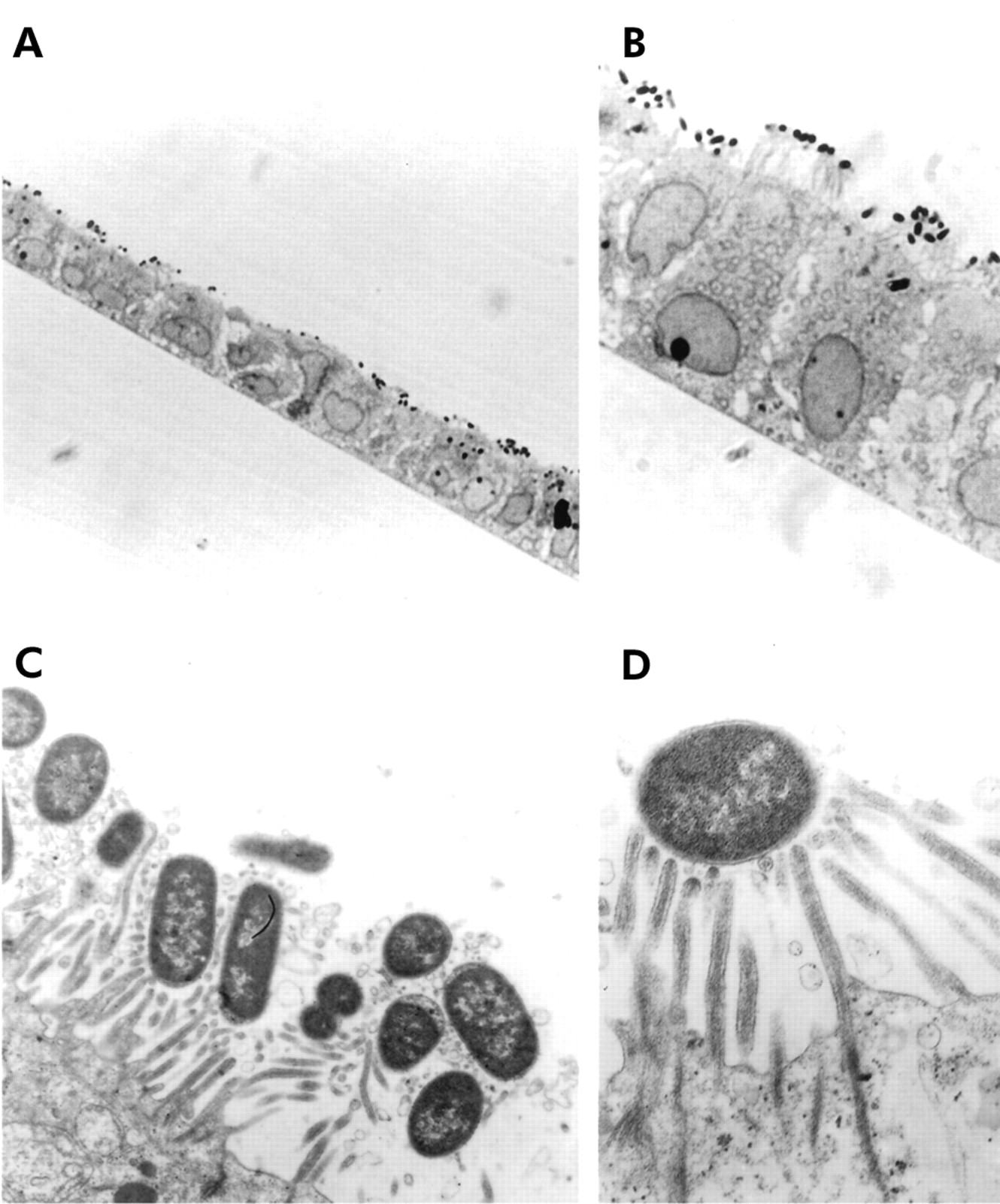
Adhering C1845 bacteria were observed in confluent differentiated Caco-2/TC7 cells infected apically (108 cfu/ml, three hours at 37°C). As can be seen in fig 1A and 1B, low and high magnifications of a thin section of the C1845 infected Caco-2/TC7 monolayer showed that the C1845 bacteria infected diffusely the brush border of the fully differentiated Caco-2/TC7 cells. Examined at the level of the brush border, a high magnification transmission electron micrograph showed that the infecting C1845 bacteria were strongly engulfed within elongated MV (fig 1C) and associated with a high number of elongated MV (fig 1D).

Adhering Afa/Dr DAEC C1845 bacteria at an early stage of adhesion onto human enterocyte-like Caco-2/TC7 cells. Confluent differentiated Caco-2/TC7 cells were infected apically at 37°C in a 10% CO2/90% air atmosphere for three hours with C1845 bacteria (108 cfu/well). (A, B) Infected cell monolayers processed for light microscopic examination of semithin sections perpendicular to the bottom of the flask showed apically adherent C1845 bacteria (magnification ×40 in (A); ×100 in (B)). (C, D) Low and high magnification of transmission electron micrographs of adhering bacteria showing bacteria engulfed within microvilli (C) and a bacteria associated with multiple elongated microvilli (D) (magnification ×4000 and ×8000, respectively).
The fact that adhering C1845 bacteria were in a close association with MV that engulf the bacteria could hinder their access by antibacterial treatment. The effect of LBS-SCS treatment against adhering C1845 bacteria onto Caco-2/TC7 cells was examined (table 1). A preliminary experiment examined whether or not LB-SCS treatment modified the cell and monolayer integrity of C1845 infected cells. For this purpose, cell viability was measured by determination of release of the intracellular enzyme LDH in the culture medium. No release was observed in LB-SCS treated infected cells compared with untreated cells (control C1845 infected cells: 36 (5) U/l; LB-SCS treated C1845 infected cells: 37 (4) U/l; H2O2 lysed cells: 4272 (61) U/l). A highly significant 3 logs decrease in viable adhering C1845 was observed in preinfected Caco-2/TC7 cells after one hour of treatment with a twofold concentrate of LB-SCS whereas only a 1 log decrease was observed with MRS at this concentration (table 1). To compare the antibacterial efficacy of LB-SCS against adhering C1845 bacteria with an antibiotic treatment, additional experiments were conducted using gentamicin, which was found to be effective against cell associated Afa/Dr DAEC.28 This antibiotic, which does not diffuse or penetrate the infected cells, kills extracellular bacteria.12,14 Examined as a function of concentration, gentamicin treatment for one hour at 37°C resulted in a dose dependent decrease in the viability of adhering C1845 bacteria onto Caco-2/TC7 cells (table 1). Gentamicin at a concentration of 100 μg/ml promoted a highly significant decrease in viable adhering C1845 bacteria, an effect which was comparable with that of a twofold concentrate of LB-SCS (table 1).
The long term effect of the antibacterial activity of LB-SCS was examined by determination of the level of viable adhering C1845 bacteria as a function of time post-treatment. For this purpose, cells were infected as reported above (108 cfu/ml, three hours at 37°C) followed by extensive washes with PBS to eliminate non-adhering bacteria. Then, C1845 infected cells were subjected for one hour to DMEM with or without LB-SCS (twofold concentrate). As shown in fig 2, a highly significant 3 logs decrease in viable adhering C1845 was observed in preinfected Caco-2/TC7 cells after one hour of treatment with LB-SCS. In C1845 infected cells treated with LB-SCS for one hour and then subcultured in the presence of DMEM, the level of viable adhering bacteria was determined at one, two, and three hours post-treatment. The level of remaining viable adhering bacteria after one hour of treatment remained stable over three hours post-treatment.

Activity of the spent culture supernatant of Lactobacillus acidophilus LB culture (LB-SCS) as a function of time on the viability of Afa/Dr DAEC strain C1845 that were adherent to human enterocyte-like Caco-2/TC7 cells. Confluent differentiated Caco-2/TC7 cells were infected apically at 37°C in a 10% CO2/90% air atmosphere for three hours with C1845 bacteria (108 cfu/well), and then subjected for one hour to De Man, Rogosa, Sharpe broth (MRS) or LB-SCS treatment (2.0-fold concentrate). After extensive washing, cells were subcultured to determine the viable adherent bacteria as a function of time post-treatment, as described in materials and methods. Control infected cells without treatment were cultured in the presence of minimal essential medium (MEM). Values are mean (SEM) of three experiments.
Protection of the F-actin cytoskeleton in enterocyte-like Caco-2/TC7 cells infected by the Afa/Dr DAEC strain C1845
We have previously reported that Afa/Dr DAEC C1845 infection in Caco-2 cells is followed by a dramatic alteration in the distribution of the major apical cytoskeleton associated proteins20,22 that play a pivotal role in MV architecture.29 We first examined whether the C1845 induced cytoskeleton injury develops as a function of the level of adhering bacteria (fig 3). For this purpose, cells were infected for three hours at 37°C with increasing concentrations of C1845 bacteria. The infected cells were processed for determination of viable adhering bacteria and for direct immunofluorescence labelling of F-actin using the fluorescein labelled phalloidin assay. Quantification of apical F-actin expression was conducted by measuring relative F-actin immunofluorescence intensity by a conventional epifluorescence microscope coupled to an image analyser. As shown in fig 3A, the decrease in F-actin immunofluorescence examined over a range of concentrations of infecting C1845 bacteria from 5.7 to 8.48 log cfu/ml demonstrated that dose dependent C1845 induced F-actin rearrangement developed. Indeed, an infecting concentration of 7.7 log cfu/ml corresponding to a level of adhering C1845 bacteria of 6.8 log cfu/ml was necessary to observe the appearance of the apical F-actin disassembly. This phenomenon was characterised by the disappearance of the homogenous, fine, and flocculated F-actin immunolabelling which corresponds to MV associated F-actin29 accompanied by a highly significant decrease in F-actin relative immunofluorescence intensity. The activity of LB-SCS (2.0-fold concentrate) was examined in Caco-2/TC7 cells infected for three hours at 37°C with C1845 bacteria (108 cfu/ml) (table 2). LB-SCS treatment inhibited C1845 adhesion. When examining F-actin expression in C1845-LB-SCS treated infected cells to which a small number of C1845 bacteria adhered, we found no alteration in the F-actin network as no change in F-actin relative immunofluorescence intensity was observed compared with control uninfected cells.
Effect of LB-SCS treatment on C1845 adhesion and C1845 induced rearrangements of brush border associated structural F-actin in enterocyte-like Caco-2/TC7 cells
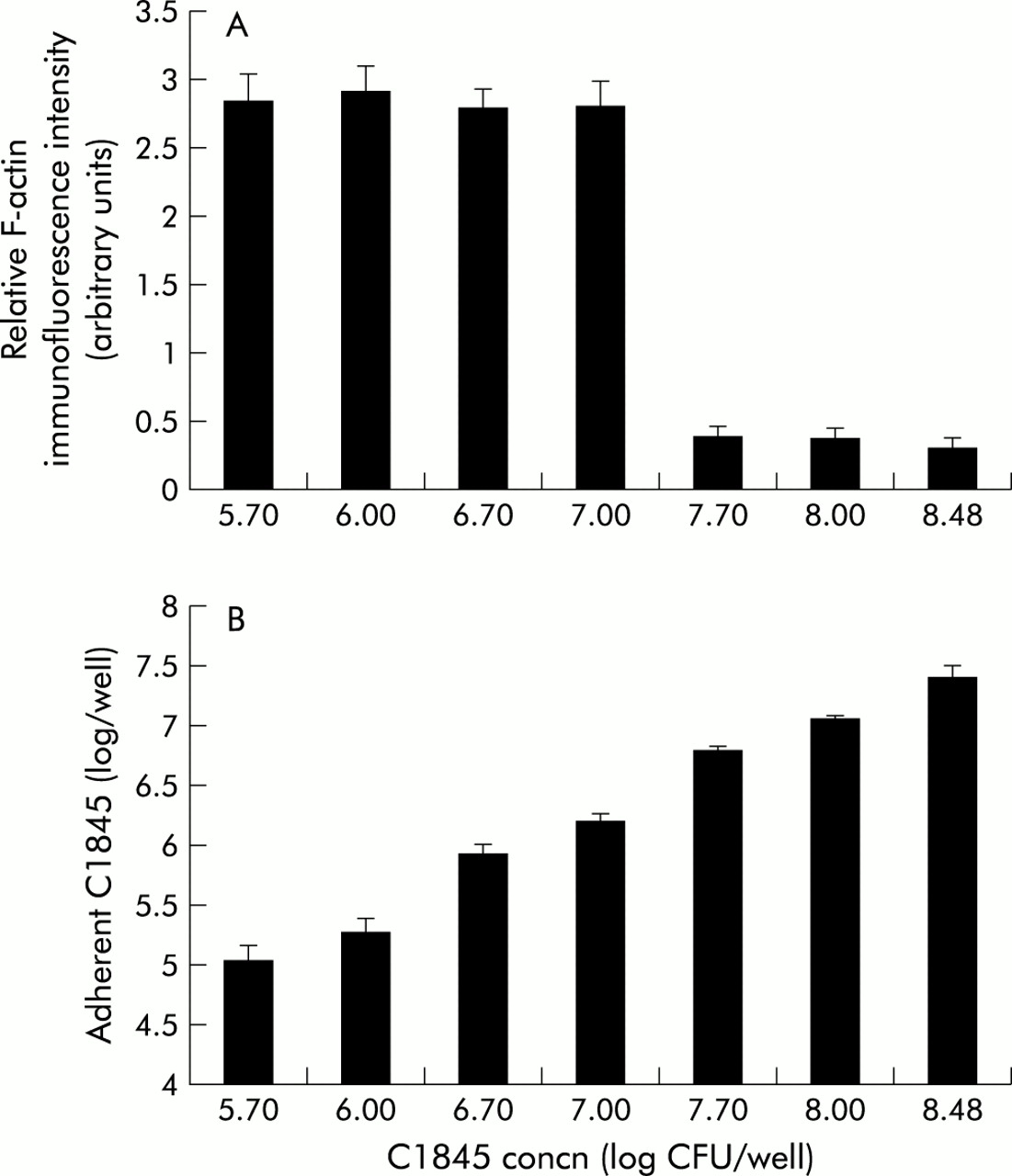
Afa/Dr DAEC C1845 induced F-actin rearrangements in human enterocyte-like Caco-2/TC7 cells develop as a function of the level of infecting bacteria. Confluent differentiated Caco-2/TC7 cells were infected apically at 37°C in a 10% CO2/90% air atmosphere for three hours with C1845 bacteria. Fixed and permeabilised cells were processed for direct immunofluorescence labelling of F-actin with fluorescein labelled phalloidin, and relative immunofluorescence intensity (arbitrary units) was measured as described in materials and methods. (A) Quantification of the F-actin rearrangements as a function of the level of infecting bacteria. (B) Determination of the level of adhering C1845 bacteria as a function of the level of infecting bacteria.
Because the resident lactobacilli in the intestine could secrete low levels of antimicrobial compounds, we conducted additional experiment to determine whether LB-SCS at a concentration which has no antibacterial activity modifies C1845 induced F-actin rearrangement. Infection of cells with C1845 bacteria (108 cfu/ml, three hours at 37°C) was conducted in the presence of LB-SCS (0.5-fold concentrate). At this concentration, LB-SCS had no activity on C1845 bacterial viability (table 1) or adhesion (C1845 infected cells: 7.25 (0.3) log cfu/ml; C1845 infected LB-SCS treated cells: 7.03 (0.4) log cfu/ml). As observed in fig 4E, high magnification of en face micrographs obtained by conventional epifluorescence microscopy showed that in the presence of LB-SCS, no C1845 induced F-actin disassembly was found compared with C1845 infected cells (fig 4B). Indeed, the normal fine flocculated labelling of F-actin centrally in C1845 infected LB-SCS treated cells (fig 4E) resembled the F-actin distribution in control uninfected cells (fig 4A). Activity was quantified by measuring relative F-actin immunofluorescence intensity in control uninfected untreated cells, untreated C1845 infected cells, and LB-SCS treated C1845 infected cells (table 3). A highly significant increase in F-actin immunofluorescence intensity was observed in LB-SCS treated C1845 infected cells compared with untreated C1845 infected cells. No significant difference was found between control uninfected untreated and LB-SCS treated C1845 infected cells.
Effect of LB-SCS treatment at a concentration which did not affect bacterial viability or adhesion on C1845 induced rearrangements of brush border associated structural and functional proteins in enterocyte-like Caco-2/TC7 cells

Effect of spent culture supernatant of Lactobacillus acidophilus LB culture (LB-SCS) treatment at concentrations which did not affect Afa/Dr DAEC C1845 viability or adhesion, on induced F-actin rearrangements in human enterocyte-like Caco-2/TC7 cells. Confluent differentiated Caco-2/TC7 cells were infected apically at 37°C in a 10% CO2/90% air atmosphere for three hours with C1845 bacteria (108 cfu/well) in the presence of LB-SCS (0.5-fold concentrate). Cells were processed for direct immunofluorescence labelling of F-actin with fluorescein labelled phalloidin, as described in fig 3. (A) Control uninfected cells. (B) C1845 infected cells. (C) Adhering C1845 bacteria in C1845 infected cells revealed by tetramethyl rhodamine isothiocyanate immunolabelling with an antiadhesin antibody. (D) Uninfected cells treated with LB-SCS (0.5-fold concentrate). (E) C1845 infected cells treated with LB-SCS (0.5-fold concentrate). (F) C1845 infected cells treated with the Ca2+ chelator BAPTA/AM (25 μM) (magnification ×100). In (E) and (F), note that adhering bacteria appeared at the cell surface. En face micrographs are representative of three experiments conducted in duplicate.
As shown in fig 4F, BAPTA/AM treatment of C1845 infected cells resulted in blockade of induced apical F-actin disassembly, as previously reported by us.21,22 BAPTA/AM activity quantified by measuring relative F-actin immunofluorescence intensity showed no significant difference compared with C1845 infected LB-SCS treated cells (control uninfected cells: 2.78 (0.18); C1845 infected cells: 0.32 (0.09); C1845 infected BAPTA/AM treated cells: 2.74 (0.15); C1845 infected LB-SCS treated cells: 2.70 (0.20)). It is interesting to note that protection of the F-actin network observed above for LB-SCS treatment (fig 4E) resembled the activity of BAPTA/AM treatment.
Inhibition of C1845 induced alterations in expression of functional brush border associated proteins
We have previously reported that infection of Caco-2/TC7 cells by Afa/Dr DAEC strain C1845 is followed by a dramatic decrease in the apical expression of the differentiation associated brush border associated hydrolases SI and DPP IV, and the fructose transporter GLUT5.22 The impact of LB-SCS at a concentration which does not affect bacterial viability or adhesion on expression of brush border associated functional proteins in C1845 infected cells was examined. Caco-2/TC7 cells were apically infected with C1845 alone (108 cfu/ml, three hours at 37°C) or in the presence of LB-SCS (0.5-fold concentrate). Indirect immunofluorescence labelling was conducted in unpermeabilised cells thus allowing us to detect the protein present within the brush border. We chose to examine expression of four brush border associated functional proteins, the hydrolases SI and DPP IV, the GPI anchored protein AP, and the fructose transporter GLUT5.
Immunolabeling of SI, DPP IV, and GLUT5 observed by conventional immunofluorescence microscopy in unpermeabilised control uninfected cells revealed the typical mosaic pattern of distribution of MV associated functional proteins.23,24,30 For AP distribution, the fine punctuate labelling was typical of the distribution of brush border GPI anchored proteins.31 In C1845 infected cells, the apical distribution of SI, DPP IV, GLUT5, and AP was profoundly disorganised compared with control uninfected cells. Central labelling in cells totally disappeared, showing instead lucent zones. The relative immunofluorescence intensities of SI, DPP IV, GLUT5, and PA measured in C1845 infected cells were highly significantly decreased compared with control uninfected cells (table 3). When SI, DPP IV, GLUT5, and AP immunolabelling was examined in C1845 infected LB-SCS treated cells, the apical distribution of this brush border associated protein was found to be maintained. The relative immunofluorescence intensities of SI, DPP IV, GLUT5, and AP measured in C1845 infected LB-SCS treated cells were not significantly different from those measured in control uninfected cells (table 3).
The observed disappearance of expression of brush border associated functional proteins from the apical surface of C1845 infected cells described above and the fact that on LB-SCS treatment these functional proteins remained present, prompted us to examine the intracellular distribution of these proteins. For this purpose, CLSM analysis in permeabilised cells was conducted which allowed observation of immunolabelled SI, DPP IV, GLUT5, and AP proteins at successive sections of the polarised cells (table 3). For example, as shown in fig 5, by reconstruction from x–z sections of CLSM, an optical section perpendicular to the plane of monolayer (x– z) shows that expression of DPP IV in control uninfected Caco-2/TC7 cells was strikingly localised at the apical surface of the cell monolayers showing a homogenous and tight band (fig 5D, table 3). This observed distribution is consistent with the known insertion of functional proteins in the brush border membrane of epithelial cells of the small intestine.24,30 In C1845 infected cells, CLMS analysis revealed that proteins remained present but significant enlargement of the bands was observed (fig 5E, table 3). In C1845 infected LB-SCS treated cells, SI, DPP IV, GLUT5, and AP immunolabelling remained tight without large lucent zones (fig 5F, table 3).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Effect of spent culture supernatant of Lactobacillus acidophilus LB culture (LB-SCS) treatment at concentrations which did not affect Afa/Dr DAEC C1845 viability or adhesion, on induced dipeptidylpeptidase IV (DPP IV) rearrangements analysed by confocal laser scanning microscopy (CLSM) in human enterocyte-like Caco-2/TC7 cells. Experimental conditions were as in fig 3. Cells were fixed with 3.5% paraformaldehyde, washed, and processed for indirect immunofluorescence labelling of DPP IV (tetramethyl rhodamine isothiocyanate labelling), as described in materials and methods. The samples were analysed by serial optical horizontal sectioning using a Zeiss confocal laser scanning microscope. Section starts at the apical domain of the cells and the following analysis was conducted until the basal domain. (A, D) Control uninfected cells. (B, E) C1845 infected cells. (C, F) C1845 infected cells treated with LB-SCS (0.5-fold concentrate). (A, B, C) Light microscopy micrographs of semithin sections perpendicular to the bottom of the flask (magnification ×40). (D, E, F) En face micrographs of the lateral views of the immunolocalisation of DPP IV obtained by CLSM analysis (vertical x–z optical section). In control cells, DPP IV labelling was localised at the apical domain in a homogenous band (D). In C1845 infected cells, apical DPP IV labelling was dramatically modified showing disruption in apical labelling and rearrangement of the protein in a non-homogenous band (E). In C1845 infected cells treated with LB-SCS (0.5-fold concentrate), DPP IV labelling remained localised at the apical domain in a homogenous band (F), as observed in control uninfected cells, and adhering bacteria appeared at the apical domain of the cells (C). Micrographs are representative of two experiments conducted in duplicate. Similar results were obtained for GLUT5 and AP (not shown).
DISCUSSION
We examined whether a lactobacillus strain isolated from the human adult microflora protected infected human intestinal cells against brush border lesions induced by a diarrhoeagenic E coli. We found that the spent culture supernatant of Lactobacillus acidophilus LB culture (LB-SCS) which contained antibacterial compounds13,14 efficiently killed the Afa/Dr DAEC C1845 bacteria, both when direct contact between the pathogen and LB-SCS was performed, and when LB-SCS treatment was applied to bacteria that adhered to the human intestinal cells Caco-2/TC7. The antibacterial activity of LB-SCS against strain C1845 resembled that previously observed against S typhimurium infecting cultured human enterocyte-like Caco-2 cells and conventional mice.13,14 We have previously demonstrated that the antibacterial substances secreted by the L acidophilus LB strain were heat stable, of low molecular weight, and distinct from lactic acid.13 Interestingly, we have recently reported that bifidobacteria strains isolated from the human infant microflora also produced low molecular weight antimicrobial substances which were active in vitro against pathogens and in vivo against S typhimurium infecting germ free mice.7 Some of the antimicrobial molecules produced by species of the gut microflora and involved in the barrier effect remain to be purified and chemically characterised.
We reported here that the human L acidophilus strain LB by its SCS antagonised alterations in expression of brush border associated structural and functional proteins promoted by a diarrhoeagenic E coli in cultured human enterocyte-like cells. In the gut epithelium, polarised organisation of intestinal cells is essential for achievement of specialised functions, including absorption and secretion.24 Indeed, this polarisation results in the presence of three domains in the plasma membrane: the apical surface facing the lumen, the lateral surface facing adjacent cells, and the basal surface underlying connective tissue. The brush border facing the external compartment is composed of a dense lawn of uniform MV for which the ultrastructure and biochemical composition have been extensively investigated.29,30 The cytoskeleton rearrangements observed on Afa/Dr DAEC infection in intestinal Caco-2/TC7 cells were characterised by a dramatic alteration in F-actin and villin protein distribution,20,22 which play a pivotal role in the establishment and maintenance of MV architecture.29 Potential implications in functional disorders resulting from alterations in the cytoskeleton of polarised epithelial cells have been demonstrated in a variety of disease processes.32 Alterations in MV associated proteins, such as enzymes, transporters, and ion channels, are characteristic of infectious diarrhoeic diseases. For example, loss of MV in chronic and protracted diarrhoea was accompanied by a substantial decrease in dissacharidase activities.33–37 Interestingly, epidemiological studies indicated that Afa/Dr DAEC strain C1845 was implicated in protracted diarrhoea in infants.15–18
To infect host cells, microbial pathogens use very sophisticated mechanisms of pathogenicity.38 Many enteroadherent and enteroinvasive pathogens hijack the host cell signal transducing pathways and/or target the host cytoskeleton molecules that play a pivotal role in cell architecture, thus promoting development of diarrhoea. It is well known that Ca2+ regulated events play a key role in the disassembly brush border organisation leading to functional disorders promoting diarrhoea.39 For example, the diarrhoeagenic rotavirus40 Ca2+ dependent brush border rearrangements observed in infected intestinal cells41,42 are accompanied by a decrease in brush border expression of SI.43 We have previously established that the C1845 induced rearrangements in MV associated F-actin in human intestinal cells result from Ca2+ dependent signalling as they are antagonised by chelation of intracellular Ca2+.21,22 Based on the results presented here and in order to explain the mechanism by which the strain LB by its SCS exerts a protective effect against the C1845 induced insult in the brush border, it is tempting to speculate that the observed antagonistic activity results from interference with C1845 induced cell signalling. An identical mechanism of action—that is, inhibition of the cross talk between a pathogen and target host cells—has previously been reported with antibiotics.44–47
One of the major roles of the gut microflora is as a barrier against colonisation of the gastrointestinal tract by pathogenic bacteria. The present report provides new insight into the mechanism by which species of the human gut microflora exert a protective role against pathogens. Indeed, the major result reported here is that a lactobacillus strain isolated from the human adult gastrointestinal microflora can antagonise cellular damage promoted by a diarrhoeagenic E coli without affecting the viability or adhesive capacity of the pathogen. To our knowledge, this is the first example of a species present in the human intestinal microflora being able to antagonise the structural and functional damage promoted by an enterovirulent pathogen in the brush border of enterocytes, probably by interfering with the cross talk between the pathogen and host cells.48,49
Acknowledgments
This work was supported by a research contract between the Institut National de la Santé et de la Recherche Médicale (INSERM) and Laboratoire du Lactéol (Houdan, France). We thank V Nicolas (Imagerie cellulaire-IFR75-ISIT) for his expert assistance on confocal laser scanning microscopy analysis, Alain Barbat (INSERM, Unité 504) for his expert assistance in transmission electron microscopy, J Grelier (INSERM SC6) for his skills in producing the art drawings, and M Métioui for his careful reading of the manuscript.