Article Text

Statistics from Altmetric.com
- MDP, minimal distending pressure
- l-NMMA
- NG-monomethyl-l-arginine
- NO, nitric oxide
- VIP, vasoactive intestinal polypeptide
- 5-HT, 5-hydroxytryptamine
Accommodation of the stomach to a meal consists of relaxation of the proximal stomach, providing the meal with a reservoir and enabling a volume increase without a rise in pressure. The accommodation response is mediated via a vagovagal reflex pathway which activates intrinsic non-adrenergic non-cholinergic neurones in the gastric wall.1 Several lines of evidence suggest a role for both nitric oxide (NO) and vasoactive intestinal polypeptide (VIP) as inhibitory neurotransmitters mediating gastric relaxation.2–,12 In in vivo studies in animals, gastric accommodation is significantly inhibited by NO synthase inhibitors.10–,12 In humans also, ingestion of a meal is accompanied by relaxation of the proximal stomach but it is not known whether the accommodation reflex in humans is mediated via activation of nitrergic neurones.
During fasting, gastric fundic tone in animals and humans is maintained by a vagally mediated cholinergic input.13,14 Recently, we demonstrated in cats that fasting fundic tone is also dependent on continuous nitrergic input.15 Thus fasting gastric tone may depend on a balance of cholinergic and nitrergic input. It is not known whether fasting gastric fundic tone in humans is also under a continuous nitrergic drive.
Functional dyspepsia is a clinical syndrome defined by chronic or recurrent upper abdominal symptoms without an identifiable cause by conventional diagnostic means.16 Several studies have demonstrated impaired accommodation in functional dyspepsia patients as a group.17–,20 We reported that accommodation is impaired in a subset of dyspeptic patients, and this was associated with symptoms of early satiety and weight loss.20 It is not known whether meal induced satiety is under nitrergic control in humans, and whether pharmacologically induced impairment of gastric accommodation would be accompanied by early satiety in healthy subjects.
NG-monomethyl-l-arginine (l-NMMA) is an inhibitor of NO synthase, suitable for use in humans. Recent studies using this agent have demonstrated involvement of NO in the control of interdigestive motility and in transient lower oesophageal sphincter relaxation in humans.21,22 We used these properties to test the hypothesis that NO is involved in the control of fasting and postprandial gastric tone and of meal induced satiety in humans.
MATERIALS AND METHODS
Study subjects
Twenty five healthy volunteers (16 males, aged 20–29 years) gave informed consent for participation in the study. None of the subjects had symptoms or a history of gastrointestinal disease or drug allergies, nor were they taking any medication. Informed consent was obtained from each participant. The protocol was previously approved by the ethics committee of the University Hospital.
Barostat study technique
Following an overnight fast of at least 12 hours, a double lumen polyvinyl tube (Salem sump tube 14 Ch; Sherwood Medical, Petit Rechain, Belgium) with an adherent plastic bag (1200 ml capacity; 17 cm maximal diameter), finely folded, was introduced through the mouth and secured to the subject's chin with adhesive tape. The position of the bag in the gastric fundic was checked fluoroscopically.
The polyvinyl tube was then connected to a computer driven programmable volume displacement barostat device (Synectics Visceral Stimulator, Stockholm, Sweden). To unfold the intragastric bag it was inflated with a fixed volume of 500 ml of air for two minutes with the study subject in a recumbent position, and then deflated completely. After a 10 minute equilibration period, the subjects were positioned in a comfortable sitting position with the knees slightly bent (80°) in a bed specifically designed for that purpose. After 30 minutes for adaptation, intrabag pressure was increased by 1 mm Hg every one minute to determine minimal intragastric distending pressure (MDP) as the lowest pressure level that provided an intrabag volume of 30 ml or more.23 This pressure level equilibrates intra-abdominal pressure.
Throughout all studies, systolic and diastolic blood pressures were measured at 10 minute intervals.
Influence of l-NMMA on fasting gastric tone and on the perception of gastric distensions
Seven healthy subjects (five males) underwent a gastric barostat examination three times, with an interval of one week. Graded isobaric distensions were performed in stepwise increments of 2 mm Hg starting from MDP, each lasting for two minutes, while the corresponding intragastric volume was recorded. Subjects were instructed to score their perception of upper abdominal sensations at the end of every distending step, using a graphic rating scale that combined verbal descriptors on a scale graded from 0 to 6.23 The end point of each sequence of distensions was established at an intrabag volume of 1000 ml or when subjects reported discomfort or pain (score 5 or 6). Subsequently, the pressure level was set at MDP+2 mm Hg for 30 minutes. Thereafter graded distensions were repeated. Twenty minutes prior to the second set of distensions, placebo, l-NMMA 4 mg/kg/h, or l-NMMA 8 mg/kg/h was administered in a double blind, randomised, crossover fashion via continuous intravenous infusion over 45 minutes.
Influence of l-NMMA on gastric accommodation to a meal
Seven different healthy subjects (five males) underwent a gastric barostat examination three times, with an interval of one week. The pressure level was set at MDP+2 mm Hg for at least 90 minutes. After 30 minutes, a liquid meal (200 ml, 300 kcal, 13% proteins, 48% carbohydrates, 39%; Nutridrink, Nutricia, Bornem, Belgium) was administered. Twenty minutes prior to the meal, placebo, l-NMMA 4 mg/kg/h, or l-NMMA 8 mg/kg/h was administered in a double blind, randomised, crossover fashion via continuous intravenous infusion over 60 minutes.
Satiety testing
Eleven healthy subjects (six males) participated in meal induced satiety studies. Following an overnight fast of at least 12 hours, a peristaltic pump (Minipuls 2; Gilson, Villiers-Le-Bel, France) filled one of two beakers at a fixed rate of 15 ml/min with a liquid meal (Nutridrink). Subjects were requested to maintain intake at the filling rate, thereby alternating the beakers as they were filled and emptied. At five minute intervals they were asked to score their satiety using a graphic rating scale that combined verbal descriptors on a scale graded 0–5 (1=threshold, 5=maximum satiety). Participants were instructed to cease meal intake when a score of 5 was reached. All subjects underwent the satiety test twice, with an interval of 3–7 days. Twenty minutes prior to the meal, placebo or l-NMMA 8 mg/kg/h was administered in a double blind, randomised, crossover fashion until 60 minutes after the start of the meal.
Data analysis
For the long distending periods at MDP+2 mm Hg before and after drug infusion, gastric tone was measured as mean balloon volume over consecutive five minute intervals. For each two minute distending period, the dependent variable was measured by averaging the recordings. The perception score corresponding to each distension was computed. Perception threshold was defined as the first level of pressure that evoked a perception score of 1 or more. Discomfort threshold was defined as the first level of pressure that provoked a perception score of 5 or more. Pressure-volume and pressure-perception curves were obtained from the stepwise distensions.
Average intraballoon volumes before and after administration of the drug were compared using the paired Student's t test. In addition, the amplitude of the maximal decrease in intraballoon volume was calculated as the difference between the average volume before drug administration and the lowest five minute interval mean volume after drug administration. To evaluate the effect of l-NMMA on gastric compliance and perception of gastric distension, the mean responses to the same pressures applied before and after drug administration, as well as the intercepts and slopes obtained by linear regression analysis of pressure-volume curves and distension-perception curves, were compared by Student's t test. The number of subjects that reported perception or discomfort at a given distending pressure were compared before and after drug administration using a logistic regression procedure with stratification, implying exact conditional inference. In addition, the perception and discomfort thresholds before and after l-NMMA were compared using the paired Student's t test.
Meal induced gastric relaxation was quantified by calculating the difference between the average intragastric volume for the 30 minutes before and the first 45 minutes after administration of the meal. Also, the amplitude of the maximal relaxation was calculated as the difference between the average preprandial volume and the highest postprandial five minute interval mean volume, and the time needed to reach the maximum postprandial volume was calculated. The amplitude of the relaxations after placebo and l-NMMA were compared using the Student's t test. In addition, analysis of covariance was performed on postprandial intraballoon volumes with pre-meal volume as a covariate.
For meal induced satiety testing, the average satiety score at the end of infusion of placebo or l-NMMA (60 minutes after the start of the meal) was determined and the amount of calories ingested until the occurrence of maximum satiety (score of 5) was calculated. Differences were considered to be significant at the 5% level.
RESULTS
Conduct of the study
All participants completed the studies as planned. No significant side effects occurred during administration of l-NMMA or placebo. Systolic and diastolic blood pressures were not significantly altered by administration of placebo (127 (3) v maximum 131 (3) mm Hg and 82 (2) v maximum 82 (3) mm Hg; NS) or l-NMMA 4 mg/kg/h (126 (3) v maximum 126 (2) mm Hg and 85 (3) v maximum 85 (3) mm Hg; NS). Administration of l-NMMA 8 mg/kg/h significantly increased systolic and diastolic blood pressures (127 (4) to a maximum of 132 (3) mm Hg and 81 (4) to a maximum of 89 (4) mm Hg, respectively; p<0.05).
Influence of l-NMMA on gastric fundic tone and on the perception of gastric distensions
The effect of placebo or l-NMMA on fundic tone was assessed in seven volunteers in whom MDP was 7.9 (0.3) mm Hg. The average intraballoon volume at MDP+2 mm Hg during the 15 minute period prior to the start of the infusion did not differ between the three groups (238 (60) ml, 242 (76) ml, and 272 (45) ml in the placebo, l-NMMA 4 mg/kg/h, and l-NMMA 8 mg/kg/h groups; NS). Mean intraballoon volume after the start of the infusion also did not differ significantly (217 (42) ml, 183 (48) ml, and 259 (53) ml in the placebo, l-NMMA 4 mg/kg/h, and l-NMMA 8 mg/kg/h groups, respectively; NS). The maximum volume decrease was 23 (26) ml after placebo, 72 (32) ml after l-NMMA 4 mg/kg/h, and −9 (14) ml after l-NMMA 8 mg/kg/h (NS).
Both before and after drug administration, distensions of the stomach with progressively higher set pressures produced progressively larger intragastric volumes. Prior to drug administration, the pressure-volume curves were similar in the three groups (table 1⇓). After administration of placebo, the pressure-volume curves did not differ significantly from each other or from the curves prior to drug administration (table 1⇓, fig 1⇓) (all NS). As summarised in table 1⇓, administration of placebo had no significant effect on the pressures inducing first perception or on the corresponding intraballoon volumes. Likewise, administration of l-NMMA 4 mg/kg/h or l-NMMA 8 mg/kg/h had no significant on the pressures inducing first perception or discomfort or on the corresponding intraballoon volumes (table 1⇓). These observations were confirmed by logistic regression analysis of pressure-perception curves and pressure-discomfort curves before and after placebo or l-NMMA (all NS).
Influence of placebo, NG-monomethyl-l-arginine (l-NMMA) 4 mg/kg/h, and l-NMMA 8 mg/kg/h on gastric fundus tone and perception of gastric distension in seven healthy volunteers

Pressure-volume relationships during administration of placebo, NG-monomethyl-l-arginine (l-NMMA) 4 mg/kg/h, or l-NMMA 8 mg/kg/h. No significant changes were observed. MDP, minimal distending pressure.
Influence of l-NMMA on gastric accommodation to a meal
During the 30 minute period before the meal, intragastric volumes were similar in the three groups (183 (22), 202 (49), and 172 (10) ml in the placebo, l-NMMA 4 mg/kg/h, and l-NMMA 8 mg/kg/h groups, respectively; NS). During the 40 minutes after the meal, mean intragastric volumes were 457 (21), 334 (91), and 309 (68) ml in the placebo, l-NMMA 4 mg/kg/h, and l-NMMA 8 mg/kg/h groups, respectively (fig 2A⇓) (NS). Meal induced accommodation (=mean postprandial increase in intraballoon volume) was significantly higher after placebo (274 (15) ml) compared with l-NMMA 4 mg/kg/h (132 (45) ml; p=0.03) or l-NMMA 8 mg/kg/h (82 (72) ml; p=0.03) (fig 2B⇓). Analysis of covariance confirmed a significant inhibitory effect on postprandial intraballoon volumes of both doses of l-NMMA compared with placebo (p<0.001). The maximum postprandial volume increase (368 (19), 248 (55), and 287 (98) ml in the placebo, l-NMMA 4 mg/kg/h, and l-NMMA 8 mg/kg/h groups, respectively; NS) and the time to maximum postprandial volume (20 (3), 13 (4), and 15 (6) minutes in the placebo, l-NMMA 4 mg/kg/h, and l-NMMA 8 mg/kg/h groups respectively; NS) did not differ between the three groups.

(A) Mean increase in intraballoon volume in the first 45 minutes after the meal during administration of placebo, NG-monomethyl-l-arginine (l-NMMA) 4 mg/kg/h, or l-NMMA 8 mg/kg/h. (B) Accommodation to a liquid meal during administration of placebo, l-NMMA 4 mg/kg/h, or l-NMMA 8 mg/kg/h. l-NMMA dose dependently decreased gastric accommodation. *p<0.05 compared with placebo.
Influence of l-NMMA on meal induced satiety
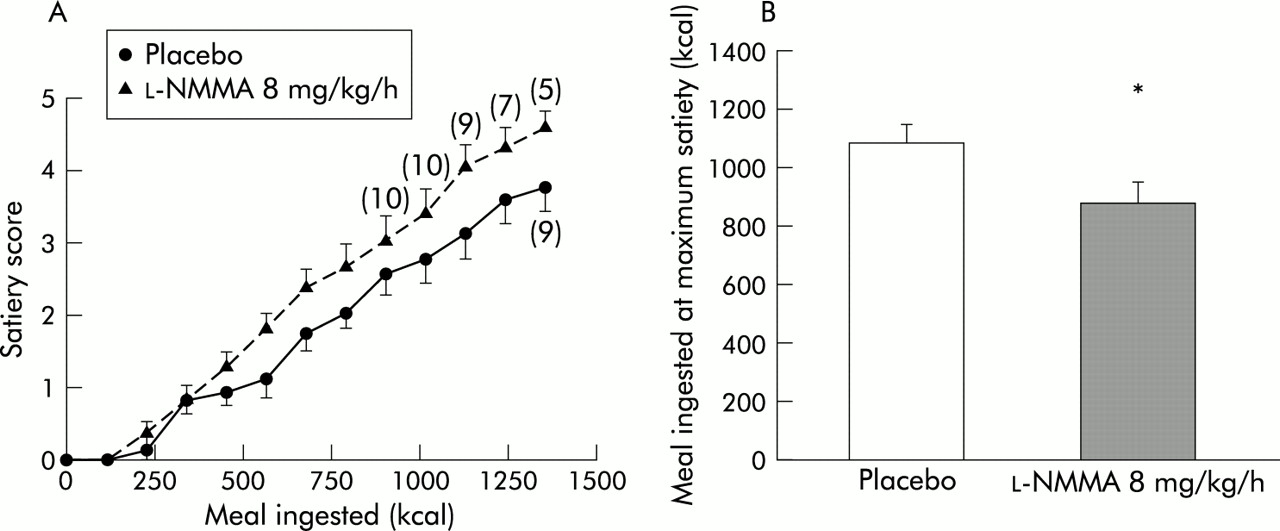
Both during placebo and l-NMMA treatment, a highly significant correlation existed between satiety scores and the amount of kcal ingested (both r=0.88, p<0.001). During the first 60 minutes of the test, the mean increase in satiety score was significantly higher after l-NMMA (0.37 (0.02)/1000 kcal) compared with placebo (0.30 (0.03)/1000 kcal; p=0.03). At the end of the infusion, 60 minutes after the start of the satiety test, the mean satiety score was significantly lower after placebo (3.73 (0.33)) than after l-NMMA (4.55 (0.21); p=0.02) (fig 3A⇓). The amount of food ingested at maximum satiety was still significantly decreased by pretreatment with l-NMMA (1058 (67) kcal v 892 (73) kcal; p<0.01) (fig 3B⇓).

{kind=link}
{kind=link}
{kind=link}
(A) Meal induced satiety scores during administration of placebo or NG-monomethyl-l-arginine (l-NMMA) 8 mg/kg/h (n=11 at each point unless otherwise indicated). (B) Calorie intake at maximum satiety during administration of placebo or l-NMMA 8 mg/kg/h. l-NMMA significantly enhanced meal induced satiety scores and decreased the amount of calories ingested at maximum satiety. *p<0.05 compared with placebo.
DISCUSSION
Recent studies indicate that impaired accommodation of the proximal stomach to a meal may be a major pathophysiological mechanism in functional dyspepsia.17–,20 Control of the accommodation reflex in humans is incompletely understood, thus hampering the development of therapeutic approaches for impaired accommodation. In the present study, we observed that NO synthase inhibition in humans did not influence fasting gastric tone or sensitivity to gastric distension. NO synthase inhibition dose dependently inhibited gastric accommodation to a meal. In addition, the NO synthase inhibitor l-NMMA (8 mg/kg/h) increased meal induced satiety.
Gastric fundic tone is the result of tonic contraction of muscle fibres of the proximal stomach. During fasting, proximal gastric tone results from a continuous state of tonic contraction that is maintained by a vagally mediated cholinergic input.13,14 In the cat, administration of the NO synthase inhibitor l-NAME results in an increase in resting fundic tone, an effect that is reversed by l-arginine, suggesting that resting fundic tone in this species is maintained by the balance of cholinergic and nitrergic drive.15 In the present study, administration of the NO synthase inhibitor l-NMMA did not result in a significant increase in resting fundic tone or in alteration of gastric compliance during stepwise distensions. In addition, NO synthase inhibition had no significant effect on sensitivity to gastric distension. These data suggest that no major nitrergic drive is contributing to resting fundic tone and to sensitivity to gastric distension in humans. Fasting gastric tone is susceptible to NO as administration of an NO donor induces proximal gastric relaxation in humans.24 A minor role for NO in balancing cholinergic resting fundic tone cannot be entirely ruled out as the doses of NO synthase inhibitor that can be safely used in humans are considerably lower than doses used in animal studies.
Variations in gastric tone are instrumental in achieving the reservoir function of the stomach by regulating both gastric accommodation and gastric emptying. The neural pathway initiating gastric accommodation to a meal is not entirely known. Gastric relaxation can be induced by nutrient perfusion into the oropharynx, into the stomach, or into the small intestine,25 and is mediated through a vagovagally driven non-adrenergic non-cholinergic mechanism.1 Both in vivo and in vitro studies suggest that the principal candidate neurotransmitters released by non-adrenergic non-cholinergic neurones during gastric accommodation are NO and VIP.2–,12 Depending on the species studied, both inhibitory neurotransmitters can act concurrently in mediating non-adrenergic non-cholinergic relaxations of the fundus,3,7–9,11 or NO can be the only mediator of non-adrenergic non-cholinergic relaxation.10,12 To our knowledge, the present study is the first to confirm that release of NO is involved in the gastric accommodation reflex in humans: meal induced relaxation was significantly decreased during NO synthase inhibition. Although a central effect of l-NMMA cannot be excluded, it seems most likely that inhibition of accommodation occurs at the level of the intrinsic gastric motor neurones, which is in keeping with animal studies and with in vitro experiments on human gastric muscle strips.8 No significant differences were noted between the two doses of l-NMMA tested but for safety reasons only a narrow dose range was studied. At the maximal concentration of l-NMMA used in this study, accommodation to a meal was not completely abolished, suggesting either incomplete inhibition of NO synthase at the dose used, involvement of another inhibitory neurotransmitter, or an additional role of decreased cholinergic tone postprandially. Lack of suitable pharmacological tools for in vivo use in humans currently precludes the use of very high doses of NO synthase inhibitor or further investigation of other putative neurotransmitters involved.
In a subset of dyspeptic patients, accommodation to a meal is impaired and this is associated with symptoms of early satiety and weight loss.20 Reversal of impaired accommodation by administration of the 5-hydroxytryptamine (5-HT)1 receptor agonist sumatriptan is associated with inhibition of meal induced satiety in these patients. In addition, prevention of gastric accommodation by administration of the fundus contracting agent erythromycin induces earlier meal induced satiety in healthy subjects26–,28 while fundus relaxing drugs such as cisapride or sumatriptan decrease meal induced satiety.27–,30 Although these observations suggest that an impaired accommodation reflex is the mechanism underlying the symptom of early satiety, a causal relationship remains to be established.31
In the present study, we demonstrated that treatment with an 8 mg/kg/h dose of l-NMMA significantly increased satiety scores during meal intake and decreased the number of calories that can be ingested until maximum satiety is reached. A previous study using a 4 mg/kg/h dose of l-NMMA failed to demonstrate a significant effect on appetite or caloric intake in humans.32 Both studies used a different method to assess meal induced satiety. In the study by Vozzo et al, subjects were allowed to eat from a buffet meal over a 30 minute period. The satiety drinking test in the present study imposes interrupted meal ingestion at a fixed caloric rate. It is unclear to what extent the different types of meal administration might affect the outcome of a pharmacological intervention. It is also possible that a dose of 4 mg/kg/h l-NMMA is inadequate to enhance meal induced satiety. However, at this dose we already obtained significant inhibition of meal induced accommodation of the proximal stomach, suggesting involvement of other pathways.
Inhibition of meal induced accommodation is only one possible explanation for the effect of l-NMMA on meal induced satiety. Several studies in animals have demonstrated that NO may be a physiological modulator of food intake and that inhibition of NO synthase decreases food intake and leads to weight loss in animals.33,34 This effect seems to involve peripheral as well as central effects. Intracerebroventricular administration of a NO synthase inhibitor, at doses that had no effect when administered peripherally, inhibited food intake in rats.35–,37 A similar central effect of NO synthase inhibition may have contributed to the effects of l-NMMA on meal induced satiety in the present study.
The influence of peripheral NO synthase inhibition on meal induced satiety may also involve factors other than gastric accommodation. As satiety also depends on the arrival of nutrients in the duodenum, loss of meal induced accommodation may lead to enhanced gastric emptying and thus may increase the perception of satiety during meal intake. Our study did not take into account the influence of l-NMMA on gastric emptying. However, neuronal NOS deficient mice do no exhibit rapid emptying but rather delayed emptying of both solids and liquids.38 In keeping with these findings, administration of an NO synthase inhibitor slowed gastric emptying rate as well as reduced food intake in mice.39 It has been suggested that delayed gastric emptying, through gastric distension and activation of vagal afferents, might underlie the decrease in food intake during NO synthase inhibition.39 However, studies in patients found no association between delayed gastric emptying and early satiety.20 In addition, reports of the satiety drinking test in humans showed that the effects of cisapride, erythromycin, and sumatriptan on meal induced satiety parallel their effects on gastric accommodation but not their effect on gastric emptying.20,27,28
In summary, we demonstrated that NO synthase inhibition in humans dose dependently inhibited gastric accommodation to a meal, suggesting involvement of a nitrergic pathway. This was accompanied by increased satiety scores during meal intake and a significant reduction in the amount of calories ingested at maximum satiety. Inhibition of NO synthase had no significant influence on fasting gastric tone or on sensitivity to gastric distension.