Article Text

Abstract
Background: A large oral dose of iron will reduce the absorption of a subsequent smaller dose of iron in a phenomenon known as mucosal block. Molecular analysis of this process may provide insights into the regulation of intestinal iron absorption.
Aims: To determine the effect of an oral bolus of iron on duodenal expression of molecules associated with intestinal iron transport in rats and to relate this to changes in iron absorption.
Methods: Rats were given an oral dose of iron and duodenal expression of divalent metal transporter 1 (DMT1), Dcytb, Ireg1, and hephaestin (Hp) was determined using the ribonuclease protection assay, western blotting, and immunofluorescence. Iron absorption was measured using radioactive 59Fe.
Results: A decrease in intestinal iron absorption occurred following an oral dose of iron and this was associated with increased enterocyte iron levels, as assessed by iron regulatory protein activity and immunoblotting for ferritin. Reduced absorption was also accompanied by a rapid decrease in expression of the mRNAs encoding the brush border iron transport molecules Dcytb and the iron responsive element (IRE) containing the splice variant of DMT1. No such change was seen in expression of the non-IRE splice variant of DMT1 or the basolateral iron transport molecules Ireg1 and Hp. Similar changes were observed at the protein level.
Conclusions: These data indicate that brush border, but not basolateral, iron transport components are regulated locally by enterocyte iron levels and support the hypothesis that systemic stimuli exert their primary effect on basolateral transport molecules.
- intestinal transport
- iron absorption
- rat
- βm, β-mercaptoethanol
- DMT1, divalent metal transporter 1
- DMT1(IRE), IRE splice variant of DMT1
- DMT1(non-IRE), non-IRE splice variant of DMT1
- GAPDH, glyceraldehyde-3-phosphate dehydrogenase
- Hp, hephaestin
- IRE, iron responsive element
- IRP, iron regulatory protein
- RPA, ribonuclease protection assay
Statistics from Altmetric.com
- βm, β-mercaptoethanol
- DMT1, divalent metal transporter 1
- DMT1(IRE), IRE splice variant of DMT1
- DMT1(non-IRE), non-IRE splice variant of DMT1
- GAPDH, glyceraldehyde-3-phosphate dehydrogenase
- Hp, hephaestin
- IRE, iron responsive element
- IRP, iron regulatory protein
- RPA, ribonuclease protection assay
Iron is essential for almost all forms of life but is toxic in excess and therefore body iron levels must be tightly regulated.1–3 As the body cannot actively excrete iron, the body iron content must be regulated at the point of absorption across the mature villus enterocytes of the duodenum.4 The main stimuli for iron absorption are changes in body iron stores and the demands of the developing erythroid mass.5–7 Although the mechanism by which the intestine is signalled of these requirements remains elusive, it is known that these signals take 2–3 days to influence absorption.8–10 It is widely accepted that this lag period represents the time taken for the immature crypt cells to migrate up the villus and differentiate, and that factors influencing iron absorption programme the crypt cells to absorb the required amount of iron on maturity.
One aspect of regulation that does not conform with the 2–3 day lag period described above is the mucosal block phenomenon, which was first reported by Hahn et al in 19438 and studied in more detail by Stewart et al in 1950.11 Both studies described the ability of a large oral dose of iron to reduce the absorption of a smaller dose administered several hours later. Due to the short time interval between doses, the initial dose must be having a direct effect on the mature enterocytes rather than the cells of the crypts. Although being discounted as the mechanism by which iron absorption is regulated by body iron requirements,12,13 examination of the molecular basis of the mucosal block phenomenon may provide insight into this process. The recent identification of several of the key molecules involved in intestinal iron absorption now allows such studies to be undertaken.
The first mammalian iron transporter to be identified was divalent metal transporter 1 (DMT1), a brush border ferrous iron transport protein.14,15 The mRNA transcript for this gene exists as at least two splice variants, one of which contains a putative iron responsive element (IRE).16 IREs are stem loop structures found in the non-coding region of various mRNA sequences and confer iron dependent regulation on these genes by virtue of their interactions with iron regulatory proteins (IRPs) (for review see Kühn17). The IRE containing splice variant of DMT1 (DMT1(IRE)) is expressed at greater levels than the non-IRE splice variant (DMT1(non-IRE)) in the duodenum,16,18,19 suggesting a role for IRPs in the regulation of intestinal DMT1 expression. The physiological relevance of the IRE in DMT1(IRE) has yet to be determined but studies have demonstrated a rapid decrease in DMT1 message and protein following an oral dose of iron in rats.20,21 Other more recently discovered molecules involved in intestinal iron absorption include the putative basolateral iron transporter Ireg1,22–24 the putative brush border ferrireductase Dcytb,25 and the intracellular ferroxidase hephaestin (Hp).19,26 The IRE/IRP system is considered unlikely to play a role in the regulation of any of these molecules in the small intestine.
Although a potential role for IRPs and DMT1 in the mucosal block phenomenon has been reported,20,21 no investigation into the effect of an oral dose of iron on duodenal expression of Ireg1, Dcytb, or Hp has been carried out, nor have the relative contributions of the IRE and non-IRE splice variants of DMT1 been evaluated. In this study we conduct a detailed examination of expression of these genes and consider the importance of luminal iron in modulating the response of the intestine to variations in systemic iron requirements.
METHODS
Animals and diets
Newly weaned male Sprague-Dawley rats were used for all experiments. Rats were weaned onto an iron deficient diet (iron content 3 mg/kg wet weight), prepared as described by Valberg and colleagues.27 They were allowed unlimited access to food and deionised water and were maintained on this diet for two weeks. Rats were then given an intragastric dose of iron (10 mg for the time course and 0–20 mg for the dose-response) as FeSO4, in 250 μl 10 mM HCl following an overnight fast. One hour after dosing, rats were returned to the iron deficient diet for the remainder of the experiment. At appropriate time points animals were anaesthetised (44 mg/kg ketamine and 8 mg/kg xylazine), and duodenal enterocytes were isolated as previously described28 and snap frozen in liquid nitrogen. All experiments described in this study were approved by the Queensland Institute of Medical Research Animal Ethics Committee.
Evaluation of iron absorption
Iron uptake and transfer were determined in duplicate rats as follows. At appropriate time points, rats were anaesthetised as described above and a midline incision made in the abdomen. The duodenum was exposed and three loose ligatures tied around it: two approximately 0.5 cm distal to the pylorus and one at the ligament of Treitz. An incision was made proximal to the upper ligature, a cannula inserted, and the upper ligature tightened. A second incision was made distal to the lower ligature. The small intestine was flushed with approximately 5 ml of saline (prewarmed to 37°C) injected through the cannula. The lower ligature was then tightened to seal off the distal end of the duodenum. The duodenal loop was then infused with 100 μl of 5 mM HCl containing 1.25 mg/ml (4.5 μM) FeSO4, and 10 μCi/ml 59Fe, and the residual solution flushed from the cannula with 150 μl saline. The second ligature at the proximal end of the loop was tightened and the cannula removed. The abdomen of the animal was covered in damp gauze to prevent drying and continually moistened with prewarmed saline.
Thirty minutes after administration of the test solution the animal was sacrificed by cervical dislocation and the duodenal loop carefully excised. The lumen was rinsed extensively with saline (approximately 30 ml) and the rinse solution retained. Radioactivity in the rinse solution, duodenum, and carcass was counted separately using a Ram DA Counter with PM-11 tube (Rotem Industries, Arava, Israel) at a distance of 10 cm. Brush border iron uptake was calculated by adding the counts in the carcass and the duodenum and expressing this as a percentage of the total radioactivity injected. Basolateral transfer was represented by the counts in the carcass and was expressed as a percentage of uptake. Whole body iron absorption assays were performed for the dose-response and were carried out as previously described.19
Ribonuclease protection assay
Total RNA was extracted from tissue samples using Trizol reagent (Invitrogen, Melbourne, Australia) as per the manufacturer’s instructions. Ribonuclease protection assays (RPAs) were performed as previously described19 using 5 μg of total RNA. The housekeeping gene glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used for normalisation. The riboprobes used corresponded to the following cDNA sequences (the data presented in parentheses indicate section of cDNA and Genbank accession number): Hp (nt1360-1573, AF246120), Ireg1 (nt1190-1365, U76714), DMT1(IRE) (nt1413-1628, AF029757), DMT1(non-IRE) (nt1413-1690, AF029757), Dcytb (nt20-259, BF558425), GAPDH (nt536-691, AF106860).
IRP RNA binding assay
Assays for IRP activity were performed as described below. A radiolabelled IRE containing riboprobe was synthesised as described above for the RPA using plasmid pGL66 which contains part of a rat L-ferritin pseudogene29 (kindly supplied by Dr Elizabeth Leibold). Before use, the probe was heated to 70°C for 10 minutes and allowed to cool to room temperature to allow the IRE to form. Protein was extracted from isolated duodenal cells by resuspending them in bandshift buffer (10 mM HEPES, 3 mM MgCl2, 40 mM KCl, 5% glycerol, 2 mM DTT, pH 7.5) containing 0.5% NP-40 and protease inhibitors (Complete; Roche Molecular Biochemicals, Sydney, Australia). The solution was incubated on ice for 15 minutes and insoluble material was removed by centrifugation at full speed in a microfuge for 10 minutes at 4°C. The protein concentration in the supernatant was determined using the Bio-Rad Protein Assay (Bio-Rad Laboratories, Hercules, California, USA).
Extracted protein (0.5 μg) was incubated in bandshift buffer with 100 000 cpm of the riboprobe for 30 minutes at room temperature to allow binding of IRPs to the labelled IRE. Loading controls were prepared by adding β-mercaptoethanol (βm) to a final concentration of 2% and incubating the samples for 10 minutes at room temperature prior to the addition of the riboprobe. This induces the full activity of IRP1 by converting all of the protein to the RNA binding form. βm has no effect on IRP2 as this protein, unlike IRP1, has constitutive RNA binding activity and is regulated instead by protein degradation.30 Unbound riboprobe was digested with 1 U of RNase T1 (Roche Molecular Biochemicals) for 10 minutes at room temperature. Heparin (Sigma-Aldrich, Castle Hill, Australia) was then added to a final concentration of 5 mg/ml and the solution incubated for a further 10 minutes at room temperature. Samples were mixed with 2 μl of gel loading buffer (40% sucrose, 0.25% bromophenol blue) and electrophoresed on a 6% non-denaturing TBE PAGE gel at 200 V for two hours. The gels were then dried and exposed to x ray film.
Antibody production
Polyclonal antibodies to Hp, Ireg1, and Dcytb were raised as previously described.19,31 A DMT1 antiserum recognising both the IRE and the non-IRE isoforms was raised in rabbits using the peptide CVKPSQSQVLRGMFV32 conjugated to diphtheria toxoid.
Immunohistochemistry
DMT1 and Dcytb protein were localised by immunofluorescence microscopy using frozen sections cut from duodenal tissue snap frozen in OCT embedding compound (Sakura Finetek USA, Torrance, California, USA) as previously described.31
Western blot analysis
Protein was extracted from isolated enterocytes and expression of Hp and Ireg1 determined by western blotting as described previously.19,31 Ferritin expression was determined in the same fashion using a rabbit antiferritin antibody (ICN Biomedicals, Seven Hills, Australia) at 1 in 5000 dilution and using 100 μg of protein sample. A rabbit antiactin antibody (Sigma-Aldrich) was used as a loading control.
Statistical analysis
All values are expressed as mean (SEM). Statistical differences between means were calculated with Microsoft Excel using the Student’s t test correcting for differences in sample variance.
RESULTS
Rats were treated with an intragastric 10 mg blocking dose of iron before being administered a test dose of oral radioactive iron at appropriate time points. The percentage of this test dose transported across the brush border membrane (uptake) and the basolateral membrane (transfer) of the duodenal enterocytes is shown in fig 1. Uptake across the brush border membrane decreased rapidly following a blocking dose of iron, from control levels of 84.6 (2.3)% to a minimum of 57.1 (3.9)% within three hours. Uptake persisted at this reduced level until the 12 hour time point before recovering to near control levels by 72 hours. Basolateral transfer followed a similar pattern, although it did not decline as rapidly, decreasing from a control value of 74.7 (2.6)% to 33.6 (7.8)% after 12 hours. Again, this was a transient decrease and transfer returned to near control levels by 72 hours.

Duodenal uptake and transfer of 59Fe in rats following a blocking dose of non-radioactive iron. Rats were fasted overnight before being administered 10 mg of oral iron. At various time points thereafter, animals were anaesthetised and uptake (A) and transfer (B) of 59Fe determined as described in the methods section. The number of hours following the initial administration of iron is shown. Data are mean (SEM) of six animals. Statistical significance is shown relative to control animals: *p<0.05, **p<0.01.
Having confirmed that a mucosal block to absorption had occurred in the rats, IRP RNA binding activity in duodenal enterocytes was examined to provide a measure of the biologically accessible iron pool in these cells. Animals used in this study had been maintained on iron deficient diets prior to dosing with iron, and thus the enterocytes were expected to be iron deficient. This was confirmed by the high RNA binding activity of both IRP1 and IRP2 in control animals (fig 2A). This decreased rapidly following the blocking dose of iron, with IRP1 RNA binding activity substantially decreased after 30 minutes and IRP2 within one hour. There was minimal RNA binding activity for both IRP1 and IRP2 at the 6–24 hour time points, but by 72 hours almost complete recovery was seen for both proteins.
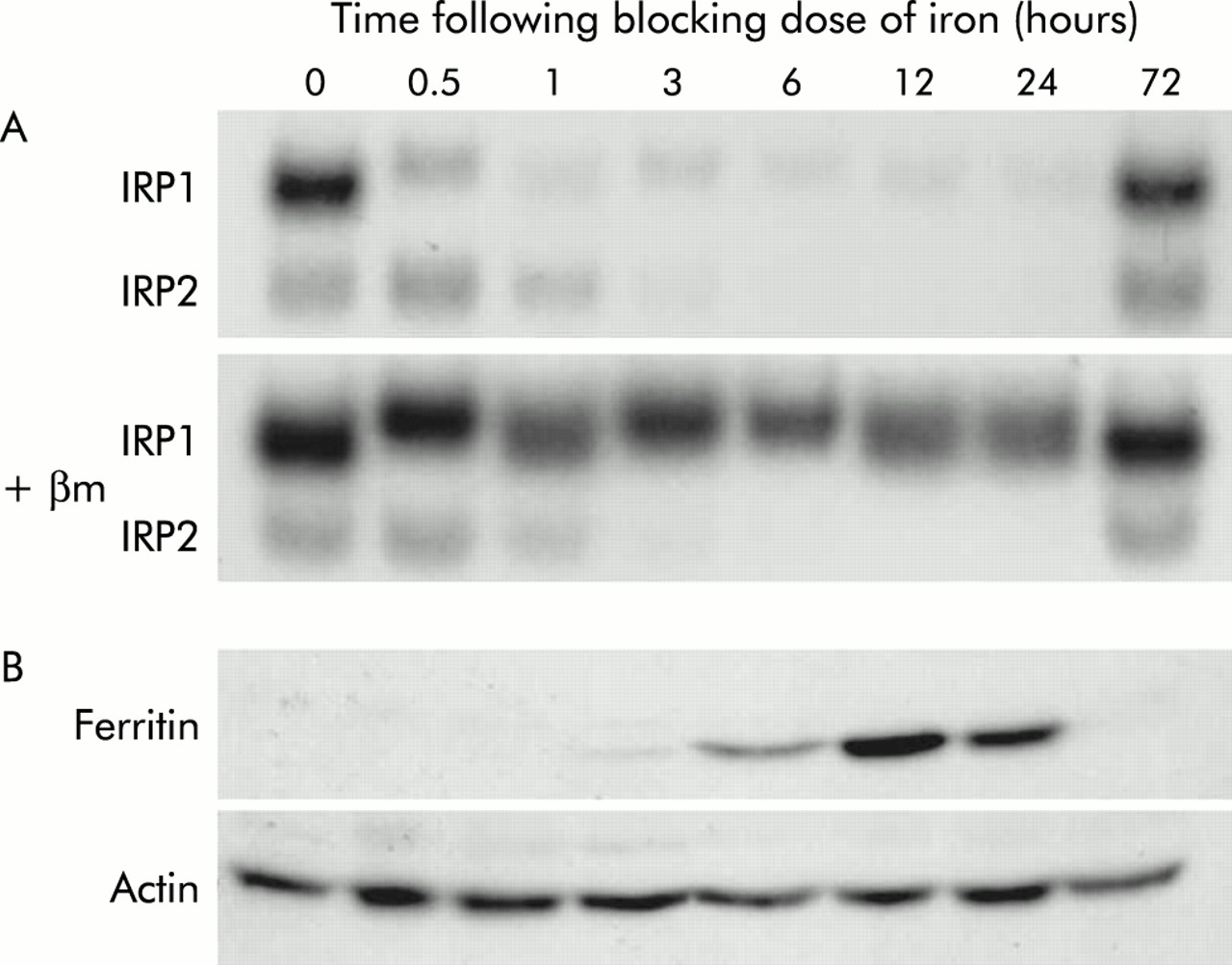
Iron regulatory protein (IRP) activity and ferritin expression in rats following orally administered iron. Rats were fasted overnight before being administered 10 mg of oral iron. At various time points thereafter, animals were sacrificed and duodenal enterocytes isolated as described in the methods section. Protein was extracted from enterocytes and analysed by IRP bandshift assay (A) and by western blotting using antibodies specific for ferritin (B). Representative assays from triplicate experiments are shown. The number of hours following iron administration is indicated. βm, β-mercaptoethanol.
Binding of IRPs to IRE sequences affects expression of a number of iron metabolism genes.17 To ensure that the change in IRP RNA binding activity seen in this study could elicit normal changes in gene expression, the amount of ferritin protein in duodenal enterocytes was examined by western blotting (fig 2B). As would be expected in animals on an iron deficient diet, no detectable ferritin was found in control animals. Ferritin protein was first detected three hours after administration of iron, reaching a maximum at 12 hours. Again, the effect was transient, with ferritin decreasing to undetectable levels by 72 hours.
To further investigate the molecular basis of the mucosal block phenomenon, expression of the genes proven or likely to be involved in iron absorption was determined using RPA analysis. Representative gels are shown in fig 3A and quantitation in fig 3B. Expression of the brush border uptake components DMT1(IRE) and Dcytb decreased dramatically following an oral dose of iron. DMT1(IRE) expression was reduced to 28.8% of control levels by three hours and reached a minimum of 4.4% of control levels by 12 hours. The decrease in Dcytb message was slower and less pronounced, reaching 10.9% of control levels by 12 hours. Expression of both DMT1(IRE) and Dcytb partially recovered to 51.0% and 55.7% of control levels, respectively, by 72 hours. No strong regulation of DMT1(non-IRE), Hp, or Ireg1 mRNAs was seen but a modest decrease in Ireg1 message to 56.9% of controls was recorded at 12 hours.

Gene expression following orally administered iron. Rats were fasted overnight before being administered 10 mg of oral iron. At various time points thereafter, animals were sacrificed and duodenal enterocytes isolated as described in the methods section. Total RNA was extracted from enterocytes and gene expression determined by ribonuclease protection assay (RPA) using 5 μg of RNA. Representative RPAs are shown for each gene (A). The number of hours following administration of iron is indicated. Band intensities were quantitated by densitometry, corrected for loading using glyceraldehyde-3-phosphate dehydrogenase (GAPDH) as a control, and graphed as a proportion of GAPDH (B). Data represent mean (SEM) of three animals. Statistical significance is shown relative to the 0 hour time point: *p<0.05, **p<0.01. DMT1, divalent metal transporter 1; DMT1(IRE), IRE splice variant of DMT1; DMT1(non-IRE), non-IRE splice variant of DMT1; Hp, hephaestin; IRE, iron responsive element.
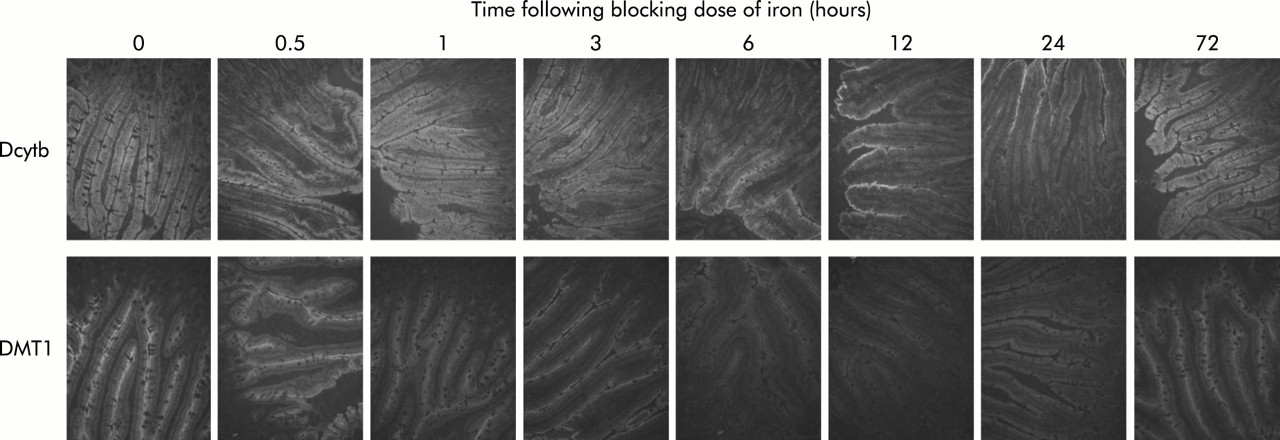
The changes in DMT1 and Dcytb mRNA expression described above were confirmed at the protein level using immunofluorescence. Both DMT1 and Dcytb protein were detected as strong fluorescence in the villus enterocytes of control animals (fig 4). This agrees with the published localisations for these two proteins.25,32,33 Following an oral dose of iron, DMT1 protein expression decreased to very low levels within six hours. Dcytb protein also declined considerably by 12 hours although some residual fluorescence remained on the brush border. Both proteins recovered to near control levels by 72 hours.

Immunofluorescence for divalent metal transporter 1 (DMT1) and Dcytb protein in the duodenum of rats following oral administration of iron. Rats were fasted overnight before being administered 10 mg of oral iron. At various time points thereafter, animals were sacrificed and duodenal sections analysed by immunofluorescence using antibodies specific for DMT1 or Dcytb, as described in the methods section. Immunohistochemistry was carried out on sections from three animals and representative sections are shown. The number of hours following administration of iron is shown. Original magnification 200×.
As there was very little change in Ireg1 or Hp message levels, any change in protein expression may be too subtle to be detected by immunofluorescence and therefore western blotting was employed to investigate this. As shown in fig 5, no change in expression of either protein was seen throughout the experiment.

Detection of Ireg1 and hephaestin (Hp) protein in the duodenum of rats following oral administration of iron. Rats were fasted overnight before being administered 10 mg of oral iron. At various time points thereafter, animals were sacrificed and duodenal enterocytes isolated, as described in the methods section. Protein was extracted from enterocytes and analysed by western blotting using antibodies specific for Ireg1 (A) or Hp (B). Representative blots are shown for each protein. The number of hours following iron administration is indicated. The sizes (×10−3) of the molecular weight standards are indicated on the right of each blot. Band intensities were quantitated by densitometry, corrected for loading using actin as a control, and graphed as a proportion of actin. Data are mean (SEM) of 5–6 animals. No statistically significant differences were seen between the groups.
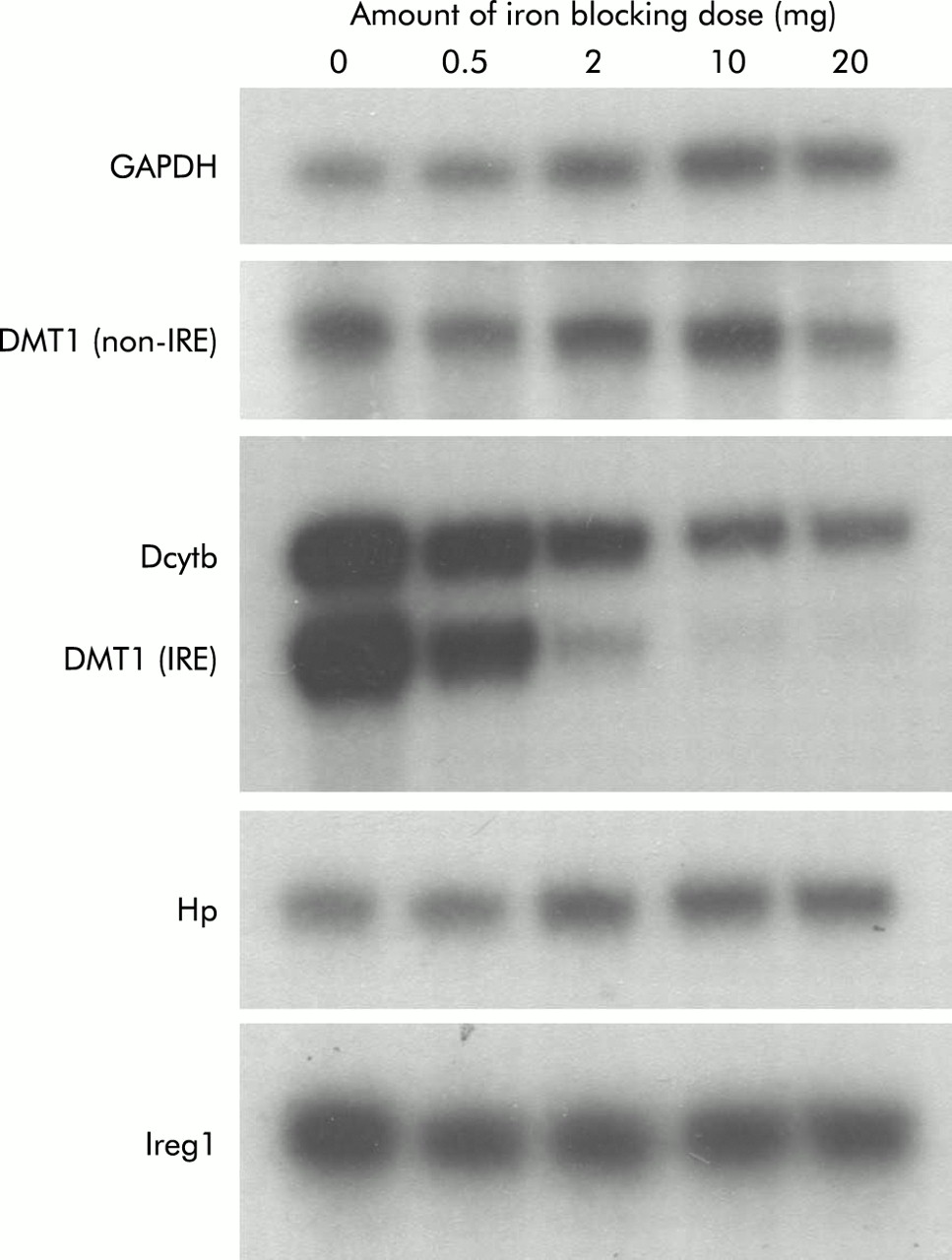
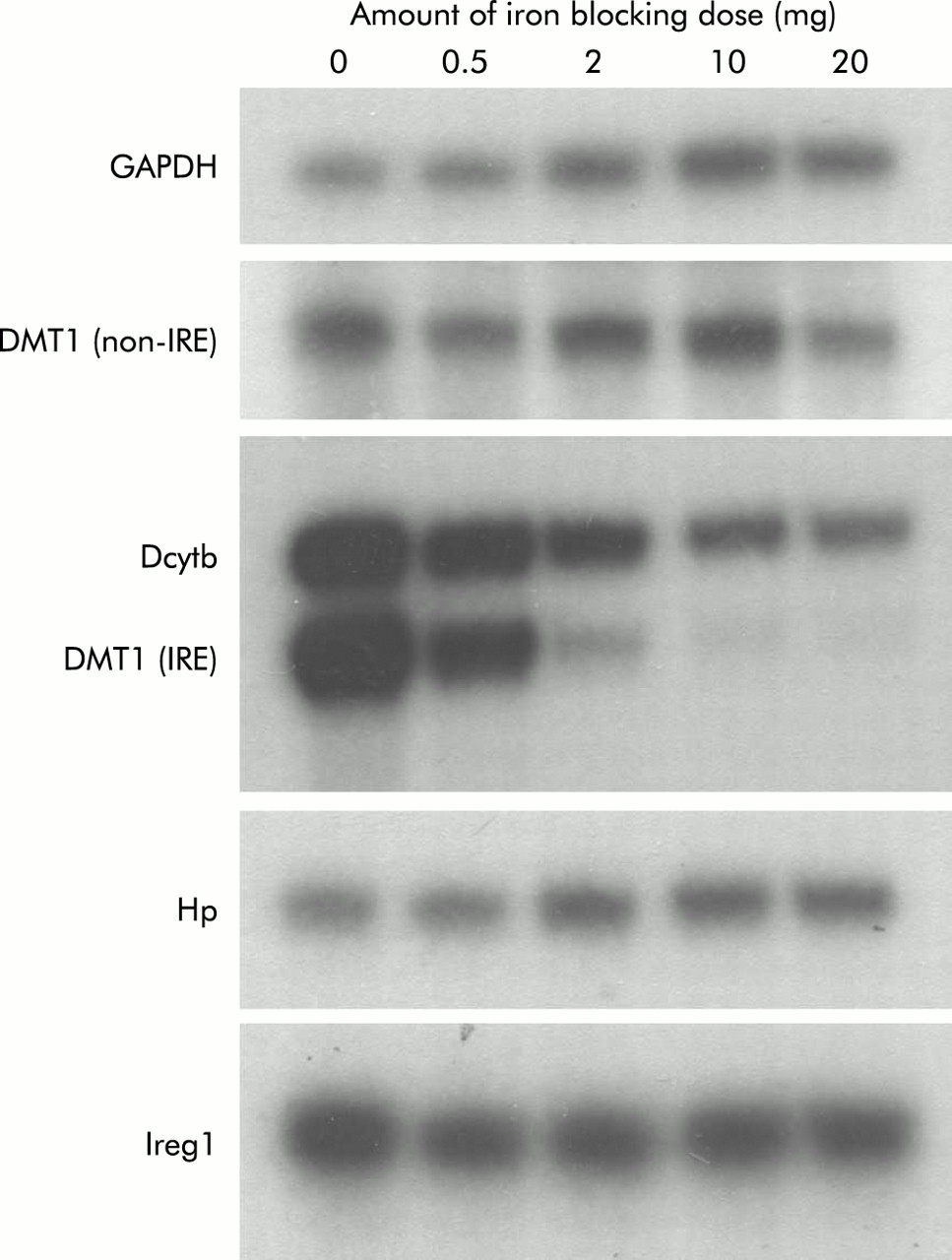
To determine whether smaller doses of iron would produce smaller changes in both absorption and brush border gene expression, we carried out a dose-response study. For these experiments, all parameters were measured six hours after administration of iron as both absorption and gene expression had changed dramatically at this time in the studies described above. Absorption was determined using intact animals, as described in the methods section, and the results are shown in fig 6. While a general trend towards decreasing absorption was seen with 0.5 mg and 2 mg of iron, statistical significance was not reached until a 10 mg dose was used. A further decrease in absorption was observed using 20 mg of iron. Examination of duodenal gene expression in these animals by RPA analysis revealed a progressive reduction in DMT1(IRE) and Dcytb expression (fig 7). Once again the decrease in Dcytb mRNA was not as dramatic as that seen for DMT1(IRE). There was little change in expression of DMT1(non-IRE), Hp, or Ireg1 messages.
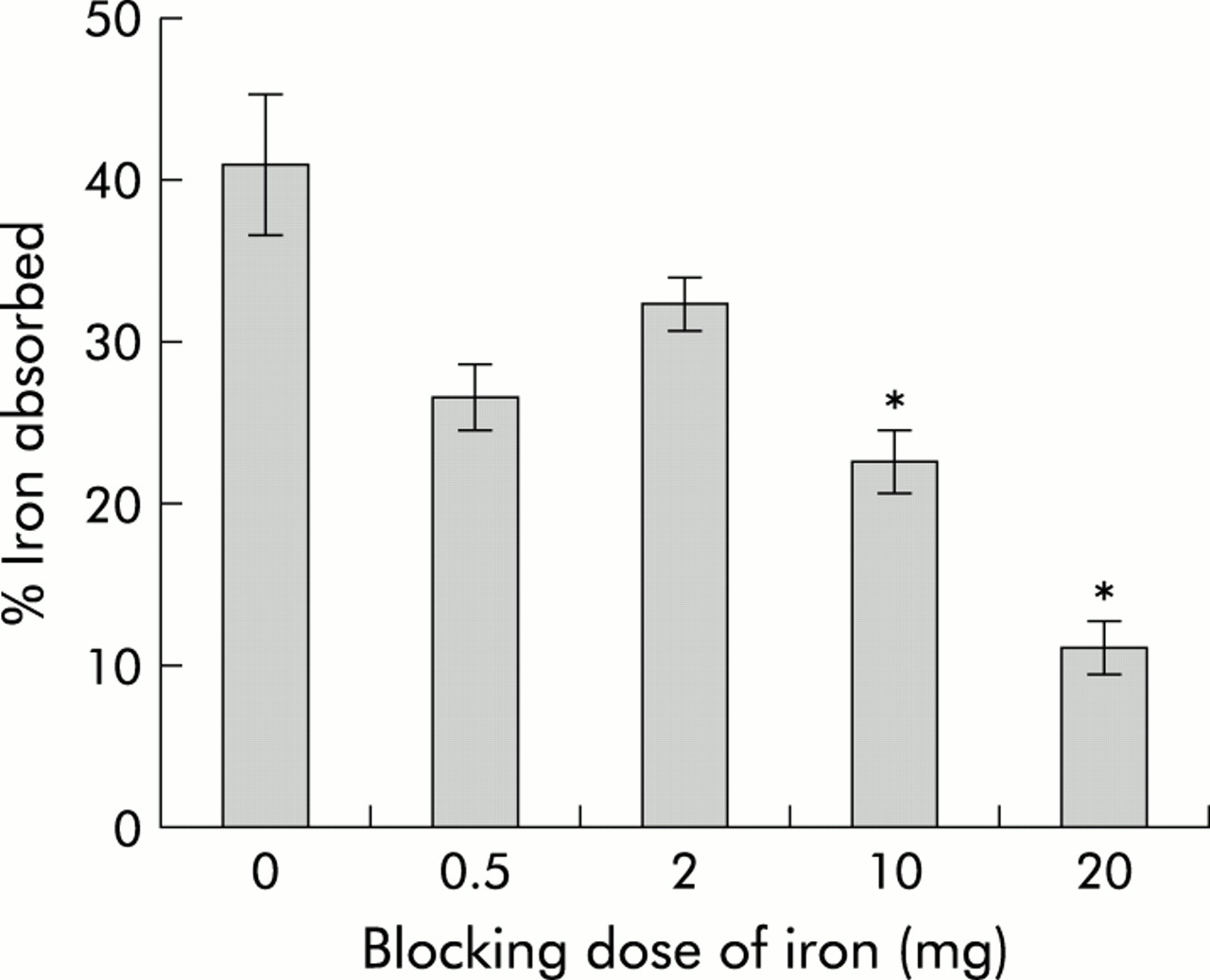
Iron absorption following varying doses of iron. Rats were fasted overnight before being administered an oral dose containing varying amounts of iron. Control animals were given 10 mM HCl only. Intestinal iron absorption was determined using an oral dose of 59Fe administered six hours after the initial dose of iron. Absorption is presented as the percentage of radioactivity retained by the animals five days after dosing. The amount of iron received by each group is indicated (blocking dose of iron). Data are mean (SEM) of three animals. Statistical significance is shown relative to control animals: *p<0.05.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Gene expression following varying doses of oral iron. Rats were fasted overnight before being administered an oral dose containing varying amounts of iron. Control animals were given 10 mM HCl only. Six hours after this dose, animals were sacrificed and duodenal enterocytes isolated, as described in the methods section. Total RNA was extracted from enterocytes and gene expression determined by ribonuclease protection assay using 5 μg of RNA. Representative assays from triplicate experiments are shown. The amount of iron received by each group is indicated (iron blocking dose). DMT1, divalent metal transporter 1; DMT1(IRE), IRE splice variant of DMT1; DMT1(non-IRE), non-IRE splice variant of DMT1; Hp, hephaestin; IRE, iron responsive element.
DISCUSSION
Our understanding of the molecular basis of intestinal iron absorption has been greatly enhanced by the recent discoveries of several key molecules crucial for this process. By applying this knowledge to the mucosal block phenomenon, we have provided some significant insights into the regulation of this important physiological process. We have shown that the mucosal block to absorption is caused by a reduction in both the brush border uptake and basolateral transfer of iron in duodenal enterocytes and this is in agreement with previous studies.34,35 The decrease in brush border uptake appears to be due in part to a rapid decrease in expression of DMT1 message and protein, again as reported previously.20,21 This study however is the first to show that the mucosal ferrireductase Dcytb is also rapidly affected by dietary iron. Our results indicate that the decrease in DMT1 message is entirely due to a decrease in the level of the DMT1(IRE) isoform as there was no detectable decrease in DMT1(non-IRE). This, together with the corresponding decrease in IRP binding activity and increase in ferritin expression, supports the involvement of IRP/IRE interactions in the regulation of enterocyte DMT1 expression. IRP/IRE interactions cannot directly explain the accompanying decrease in Dcytb message however, as mRNA does not contain an IRE motif. An alternative mechanism for the regulation of Dcytb message is also suggested by the slower and less pronounced decrease in expression that occurs following an oral dose of iron. Dcytb could be regulated at the transcriptional level, or its message could be under the control of a protein whose expression is IRP dependent, explaining the slightly delayed response. It is also possible that DMT1 is under transcriptional control but recent studies using cultured cells suggest that this is not the case.36 Demonstration that changes in the expression of DMT1 and Dcytb protein mimicked the changes in message level suggests that these proteins have a high rate of turnover and/or that they may be degraded in an iron dependent manner.
The observed decrease in iron export from enterocytes does not appear to involve a change in expression of the basolateral transporter Ireg1 or the intracellular ferroxidase Hp at the protein level. There are several possible explanations for this. The most likely explanation is the mixing of radioactive iron from the test dose with cold iron from the blocking dose that remained within the cell. This would lead to an apparent decrease in transfer due to a reduction in the specific activity of the intracellular iron available for basolateral export. The decrease in transfer may be exacerbated by increasing ferritin synthesis which reached a peak at 12 hours. This ferritin may sequester some of the iron entering the cell and impede its passage into the circulation. In time, the size of the enterocyte iron pool will decrease (as iron is moved across the basolateral membrane and cells are exfoliated) and this could explain the recovery of transfer at 24 hours despite high ferritin levels. An alternative explanation for the observed reduction in enterocyte iron export is a post-translational decrease in the activity of Ireg1 and/or Hp. Whether these proteins can be regulated in this manner has not been investigated, but irrespective of this, it is not necessary to invoke such changes to explain the reduced transfer.
The studies described in this paper provide some fundamental insights into the relative contributions of luminal and systemic signals to the regulation of iron absorption. While expression of the brush border uptake components DMT1 and Dcytb is regulated, at least in part, by intracellular iron levels, the basolateral transporter Ireg1 is not. However, we and others19,23,24 have previously shown that Ireg1 expression can be regulated by the body’s iron requirements, being increased in iron deficiency and decreased in iron loading. These observations suggest that basolateral transfer is regulated by signals from the body rather than by the local iron concentration and support our earlier hypothesis that basolateral transfer is the primary point at which intestinal iron absorption is regulated.37 In this hypothesis, signals to alter iron absorption are detected by the intestinal epithelium and result in changes in expression of Ireg1. For example, an increase in Ireg1 expression would lead to increased basolateral transfer and a decrease in enterocyte iron levels. This in turn would lead to an increase in IRP RNA binding activity, stabilising DMT1(IRE) mRNA and increasing DMT1 protein expression. The decrease in intracellular iron level would also increase Dcytb expression. The net result would be an increase in brush border iron uptake subsequent to an increase in basolateral transfer. The opposite would occur when iron requirements are low and Ireg1 expression is reduced. Iron would build up within the enterocyte, reducing IRP activity and subsequently decreasing DMT1 and Dcytb expression and, consequently, brush border iron uptake. It is possible (if not likely) that DMT1 and Dcytb levels are also modulated by systemic influences on enterocytes, but importantly expression of these molecules can be overridden by local regulation that does not affect Ireg1.
The rapid changes in DMT1 and Dcytb expression in response to intracellular iron may be a mechanism by which a constant supply of iron can be provided to the basolateral transfer mechanism regardless of dietary iron content. If the amount of dietary iron entering the cell exceeds the capacity of the basolateral transport pathway, iron will accumulate in the enterocyte leading to a reduction in expression of brush border iron transport components, which in turn will prevent a further build up of iron within the cell. If dietary intake is insufficient to meet body demands however, DMT1 and Dcytb expression can be dramatically increased to extract as much iron as possible from an iron deficient diet. This is evident from the dose-response study in which iron absorption did not change significantly with doses of 0.5 mg and 2 mg of iron despite large decreases in DMT1(IRE) and Dcytb mRNA expression. Such data show that under iron deficient conditions expression of DMT1 and Dcytb is far in excess of that necessary to adequately supply the basolateral transport mechanism.
In summary, our results indicate that brush border iron uptake is most strongly regulated by intracellular iron levels and support the proposal that basolateral transfer is regulated by systemic factors which reflect body iron requirements. The nature of these body signals is unclear but the recently identified hepcidin molecule may provide some of the answers. It is proposed that circulating hepcidin signals the body’s iron requirements to the cells of the intestine38 and we have recently reported an exquisite inverse relationship between hepcidin expression in the liver and iron transporter expression in the duodenum.31 The studies presented here suggest that a primary target of hepcidin may be Ireg1.
Acknowledgments
We thank Dr Elizabeth Leibold for providing the pGL66 plasmid. DMF was supported by a postgraduate scholarship from the Royal Children’s Hospital Foundation, Brisbane, Australia. This work was supported in part by grants from the National Health and Medical Research Council of Australia (to GJA), from the National Institute of Diabetes and Digestive and Kidney Diseases (R01 DK-57800-1 to CDV, GJA, and ATM), and from the Human Frontier Science Program (grant RGY0328/2001-M to ATM, CDV, and GJA).