Article Text

Abstract
Background and aims:Bacteroides vulgatus induces colitis in gnotobiotic HLA-B27 transgenic (TG) rats while broad spectrum antibiotics prevent and treat colitis in specific pathogen free (SPF) TG rats although disease recurs after treatment ends. Lactobacilli treat human pouchitis and experimental colitis. We investigated if Lactobacillus rhamnosus GG (L GG) can prevent colitis in TG rats monoassociated with B vulgatus and if L GG or Lactobacillus plantarum 299v (LP 299v) can treat established colitis in SPF TG rats and prevent recurrent disease after antibiotics were stopped.
Methods: Germfree B27 TG rats were monoassociated with B vulgatus for four weeks following two weeks of colonisation with L GG or no bacteria. SPF B27 TG rats received oral vancomycin and imipenem for two weeks, or water alone, followed by four weeks of treatment with oral L GG, LP 299v, or water only. Disease activity was quantified by blinded gross and histological scores, caecal myeloperoxidase (MPO) activity, and levels of interleukin (IL)-1β, tumour necrosis factor (TNF), transforming growth factor β, and IL-10.
Results:L GG did not prevent colitis in B vulgatus co-associated TG rats or treat established disease in SPF rats. However, L GG but not LP 299v prevented colitis relapse in antibiotic treated rats with reduced gross and histological scores, caecal MPO, IL-1β, and TNF whereas caecal IL-10 was increased.
Conclusions:L GG does not prevent colitis in gnotobiotic TG rats or treat established disease in SPF rats, but is superior to LP 299v in the prevention of recurrent colitis. These studies suggest that antibiotics and probiotic agents provide synergistic therapeutic effects, perhaps mediated by altered immunomodulation with selective activity of different lactobacillus species.
- colitis
- cytokines
- lactobacillus
- rat
- IBD, inflammatory bowel diseases
- GF, germ free
- TG, transgenic
- SPF, specific pathogen free
- CFU, colony forming units
- L GG, Lactobacillus rhamnosus GG
- LP 299v, Lactobacillus plantarum strain 299v
- BBE, bacteroides bile esculin
- ELISA, enzyme linked immunosorbent assay
- IL, interleukin
- PBS, phosphate buffered saline
- TNF, tumour necrosis factor
- TGF-β, transforming growth factor β
Statistics from Altmetric.com
- IBD, inflammatory bowel diseases
- GF, germ free
- TG, transgenic
- SPF, specific pathogen free
- CFU, colony forming units
- L GG, Lactobacillus rhamnosus GG
- LP 299v, Lactobacillus plantarum strain 299v
- BBE, bacteroides bile esculin
- ELISA, enzyme linked immunosorbent assay
- IL, interleukin
- PBS, phosphate buffered saline
- TNF, tumour necrosis factor
- TGF-β, transforming growth factor β
The role of intestinal bacteria in the pathogenesis of human inflammatory bowel diseases (IBD), particularly Crohn’s disease, is well recognised.1,2 These diseases typically occur at sites with the highest concentrations of intestinal bacteria, such as the colon and terminal ileum. Antibiotics and faecal diversion are effective treatment modalities for Crohn’s disease1–3 whereas re-establishing continuity of the bypassed distal bowel or infusion of intestinal contents into the excluded ileum leads to disease recurrence.4
The role of intestinal bacteria in the initiation and perpetuation of chronic intestinal inflammation is most convincingly demonstrated in experimental models of chronic intestinal inflammation in which genetically susceptible hosts develop spontaneous colitis in the presence of commensal intestinal organisms whereas no disease occurs in the germ free (GF) state.5–8
HLA-B27 transgenic (TG) rats develop colitis, gastritis, and systemic inflammation in a specific pathogen free (SPF) environment.9 However, GF TG rats and non-TG rats colonised with SPF bacteria show no evidence of colitis, gastritis, or arthritis.5,10 Colitis and gastroduodenal inflammation in TG rats is attenuated by metronidazole therapy and can be prevented and treated with broad spectrum antibiotics.11 Moreover, caecal bacterial overgrowth within an experimental blind loop exacerbates colitis whereas bypass of the caecum attenuates disease in this model, which correlates with concentrations of luminal bacteroides species.12Bacteroides vulgatus preferentially induces colitis in TG rats after monoassociation for four weeks13 whereas monoassociation with Escherichia coli does not cause disease. These findings indicate that not all bacteria are equal in their capacity to induce colitis.
Probiotics are defined as living commensal micro-organisms which are important to the health and well being of the host.14 Lactobacilli, although not predominant enteric organisms, are present throughout the gastrointestinal tract of healthy humans and rodents. Several lactobacillus strains, such as L plantarum 299v (LP 299v), L acidophilus, L fermentum, and Lactobacillus rhamnosus GG (L GG) can colonise the human gastrointestinal tract.15 Several human clinical trials have shown the efficacy of several species of probiotics in viral and bacterial induced infectious colitis as well as in antibiotic associated diarrhoea.14,16,17 Rectal administration of native murine L reuteri prevented colitis in interleukin (IL)-10 deficient mice and a similar effect was shown by the oral prebiotic lactulose, which stimulated the growth of endogenous lactobacilli species.18 Continuous oral treatment of LP 299v in GF IL-10 deficient mice colonised with SPF flora attenuated established colitis.19 A probiotic preparation including four species of lactobacilli, three bifidobacteria, and one Streptococcus salivarius (VSL# 3) maintained remission of refractory pouchitis after transient antibiotic therapy.20 This probiotic cocktail VSL#3 was also beneficial in the treatment of colitis in IL-10 deficient mice.21
The first aim of our study was to investigate if the probiotic species L GG can prevent colitis in B27 TG rats monoassociated with the disease inducing B vulgatus. Secondly, we determined if L GG or LP 299v could treat established colitis in SPF B27 TG rats or prevent recurrent disease after broad spectrum antibiotics were stopped. Thirdly, we studied the protective mechanisms of probiotics in the prevention of relapse model.
MATERIALS AND METHODS
Rats
TG rats of the 33-3 line on an inbred F344 background, bearing several copies of the genes for HLA-B27 and human β2 microglobulin,9 and their non-transgenic littermates were originally obtained from Dr Joel Taurog, Southwestern Medical School (Dallas, Texas, USA). After caesarean section they were housed in Trexler isolators in GF conditions at the Gnotobiotic Core facility at the College of Veterinary Medicine (Raleigh, North Carolina, USA). SPF TG rats and their non-TG littermates were born and maintained in filter top cages in a pathogen free environment.
Bacteria
Freeze dried cultures of L GG and LP 299v were kindly provided by ConAgra (Omaha, Nebraska, USA).
Before administration, lactobacilli strains were grown aerobically overnight at 37°C in MRS broth. The next day bacteria were harvested by centrifugation. After washes, bacteria were resuspended in sterile phosphate buffered saline (PBS) and added to the drinking water. Quantification and purity of bacterial suspensions as bacterial colony forming units (CFU) was performed in serial dilutions of the drinking water on lactobacillus MRS-vancomycin plates. B vulgatus was kindly provided by Dr AB Onderdonk (Harvard University, Cambridge, Massachusetts, USA) and was originally derived from guinea pigs with carrageenan induced colitis.22 This strain has been used to induce colitis in monoassociated TG rats.13B vulgatus was cultured anaerobically in BHI broth before colonisation of gnotobiotic rats.
Experimental design
Prevention study
In the first part of the study, GF TG rats and their non-TG littermates were divided into three groups of six rats each at the age of two months. The first two groups were monoassociated with either B vulgatus or L GG for four weeks, the third group was selectively colonised with B vulgatus for four weeks but was colonised with L GG for two weeks before co-colonisation with B vulgatus. Colonisation with B vulgatus was performed by oral and rectal swabbing using stool from other B vulgatus monoassociated rats. L GG or LP 299v was administered once in the drinking water at 1010 CFU per rat per day after overnight culture at 37°C in MRS broth. The amount of bacteria consumed by each rat per day was calculated by measured consumption of 30 ml of water per day times 3.3×108 CFU/ml=1010 CFU. Successful colonisation was assessed after one week by plating stool suspensions on bacteroides bile esculin (BBE) and MRS plates for bacteroides spp and lactobacillus, respectively, for two days of anaerobic culture, as well as by performing Gram stains. No contaminating organisms were discovered. Rats were killed after four weeks of B vulgatus or L GG colonisation. At necropsy caeca and colons were fixed in 10% buffered formalin for histological evaluations. Caeca were also snap frozen for subsequent myeloperoxidase (MPO) and cytokine analysis. Caecal contents were collected for microbiological analysis, as described below.
Prevention of relapse study
At 10 weeks of age, SPF HLA-B27 TG rats were divided into the following four treatment groups, each consisting of six rats: group 1 received only drinking water ad libitum for six weeks; group 2 received 50 mg/kg each of vancomycin/imipenem in their drinking water for two weeks followed by four weeks of water; group 3 received 50 mg/kg each of vancomycin/imipenem for two weeks followed by daily administration of L GG or LP 299v in the drinking water at 1010 CFU/rat/day for four weeks; and group 4 received two weeks of water followed by four weeks of 1010 CFU/rat/day L GG or LP 299v in their drinking water. All rats were killed six weeks after the start of treatment. Caeca and colons were collected for histology and snap frozen for MPO and cytokine analysis, as described below. Caecal contents were collected for quantitative microbiological analysis.
Histology
Tissues were prepared as previously described.10 A validated histological inflammatory score ranging from 0 to 4 was used for blinded evaluation of colonic and caecal inflammation.10
Caecal cytokine analysis
Frozen caecal tissues were thawed and lysed in PBS containing a cocktail of protease inhibitors (antipain, aprotinin, leupeptin, pepstatin A, all at 1 μg/ml and 1 mM PMSF; Sigma, St Louis, Missouri, USA).23 Tissue was then homogenised, after which the homogenate was assayed for IL-1β, tumour necrosis factor (TNF), IL-10, and transforming growth factor β (TGF-β). Concentrations of IL-1β were measured by a rat IL-1β enzyme linked immunosorbent assay (ELISA) developed by S Poole, as described previously.10 Caecal TNF and IL-10 concentrations were determined by rat cytokine specific ELISAs. TGF-β1 concentrations were measured after acidification and neutralisation according to the manufacturer using a TGF-β1 specific ELISA (Promega, Madison, Wisconsin, USA)
MPO assay
Homogenised caeca were assayed for MPO activity (units per gram of tissue), as described previously.24
Determination of caecal bacterial concentrations
Caecal contents were removed aseptically, immediately placed into an anaerobic chamber, and dissolved in sterile pre-reduced PBS. Serial 100-fold dilutions were plated and cultured anaerobically for two days on lactobacilli spp MRS vancomycin agar for quantification of lactobacillus, on BBE agar plates for bacteroides spp, and on Brucella agar (Remel, Lenexa, KS) for total anaerobic bacteria. Morphology was determined by Gram stain. Results are expressed as CFU per gram caecal contents dry weight.
Inhibition of in vitro adherence to IEC-6 cells
The rat epithelial cell line IEC-6 cells (ATCC # CRL 1592; Rockville, Maryland, USA) were grown to confluence in a 12 well tissue culture plate in antibiotic free Dulbecco’s modified Eagle’s medium containing 5% heat inactivated newborn bovine serum and 10 IU (international units)/ml insulin (Life Technologies, Burlington, Ontario, Canada).
Intestinal epithelial adherence assay was performed according to Mack and colleagues.25 Briefly, after removal of non-adherent cells, an adherent IEC-6 monolayer was incubated at 37°C with 109 CFU B vulgatus in the presence or absence of 109, 108, or 107 CFU L GG per well in 2 ml of antibiotic free complete medium under aerobic conditions. After three hours of incubation, non-adherent bacteria were removed by washing three times with warm PBS, IEC-6 cells plus adherent bacteria were collected by trypsinisation. Collected cells and adherent bacteria were then serially diluted and plated anaerobically on BHI plates for 48 hours. Bacteria were counted by colony identification, confirmed by Gram staining.
Statistical analysis
Statistics were performed using the Student’s t test or Mann-Whitney test for comparison between groups. Significance was considered to be p<0.05. All data are presented as mean (SEM).
RESULTS
Prevention study
Monoassociation of TG rats with B vulgatus resulted in mild caecal and colonic inflammation (histology score for the caecum 1.5 (0.1) and colon 1.2 (0.1)). However, pretreatment of GF TG with L GG followed after two weeks by co-association with B vulgatus did not prevent the development of mild colitis induced by B vulgatus (histology scores for the caecum 1.6 (0.2) and colon 1.1 (0.2)). However, monoassociation of gnotobiotic TG rats with L GG did not induce colitis (caecal and colonic scores both 0.2 (0.1)). Similarly, gnotobiotic non-TG rats colonised with B vulgatus, L GG, or both did not develop colitis (histology score for the caecum or colon 0.2 (0.1)).
Caecal IL-1β concentrations from B vulgatus monoassociated TG rats (12 (3) pg/mg caecal wet weight) were not significantly different from those obtained from B vulgatus and L GG co-associated TG rats (11 (2) pg/mg). Caecal IL-1 β concentrations from L GG monoassociated TG rats (5 (2) ng/g) were similar to those from gnotobiotic non-TG littermates colonised with B vulgatus, L GG, or both (2 (1) pg/mg).
The concentration of bacteroides spp in the caecal content of TG rats co-associated with B vulgatus and L GG did not differ significantly from B vulgatus monoassociated TG rats (61 (8)×109 versus 47 (4)×109 CFU/g caecal content in B vulgatus and B vulgatus plus L GG associated TG rats, respectively). The concentration of caecal L GG (0.45 (0.04)×109 CFU/g) in the B vulgatus plus L GG co-associated rats was 100-fold lower than B vulgatus concentrations but did not differ significantly from the concentration of caecal L GG in L GG monoassociated TG rats (0.49 (0.10)×109 CFU/g). Gram staining, and aerobic and anaerobic culture of caecal contents from gnotobiotic rats excluded contamination with other organisms.
Prevention of colitis relapse in SPF B27 TG rats
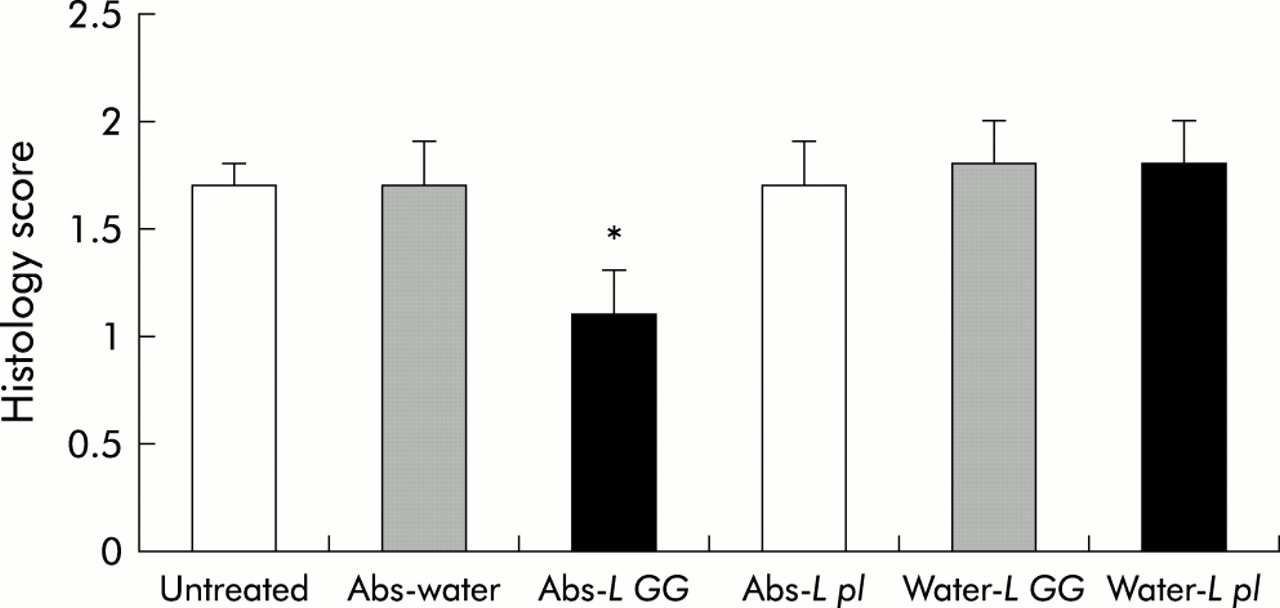
Oral administration of broad spectrum antibiotics (oral imipenem/vancomycin) for two weeks to SPF B27 TG rats followed by cessation of this treatment resulted in relapse of colitis after four weeks. Caecal gross gut scores (1.7 (0.2)) as well as total colonic histology scores (1.7 (0.2)) of antibiotic-water treated rats four weeks after stopping antibiotics did not differ from untreated control TG animals (caecal gross score 1.7 (0.2), total colonic histology score 1.7 (0.1)) (figs 1–3). Also, L GG administration alone could not effectively treat disease (caecal gross score 1.3 (0.3), total colonic histology score 1.8 (0.2)). However, if antibiotic treatment was followed by daily oral administration of L GG, relapse of colitis could be partially prevented, as shown by a significant decrease in caecal gross score (0.6 (0.1)) and total colonic histological inflammatory score (1.1 (0.3)) of this treatment group (figs 1–3). In contrast with L GG treatment, oral administration of LP 299v alone or after antibiotic treatment had no effect; neither caecal gross scores (data not shown) nor colonic histology scores (fig 3) differed significantly between treatment groups.
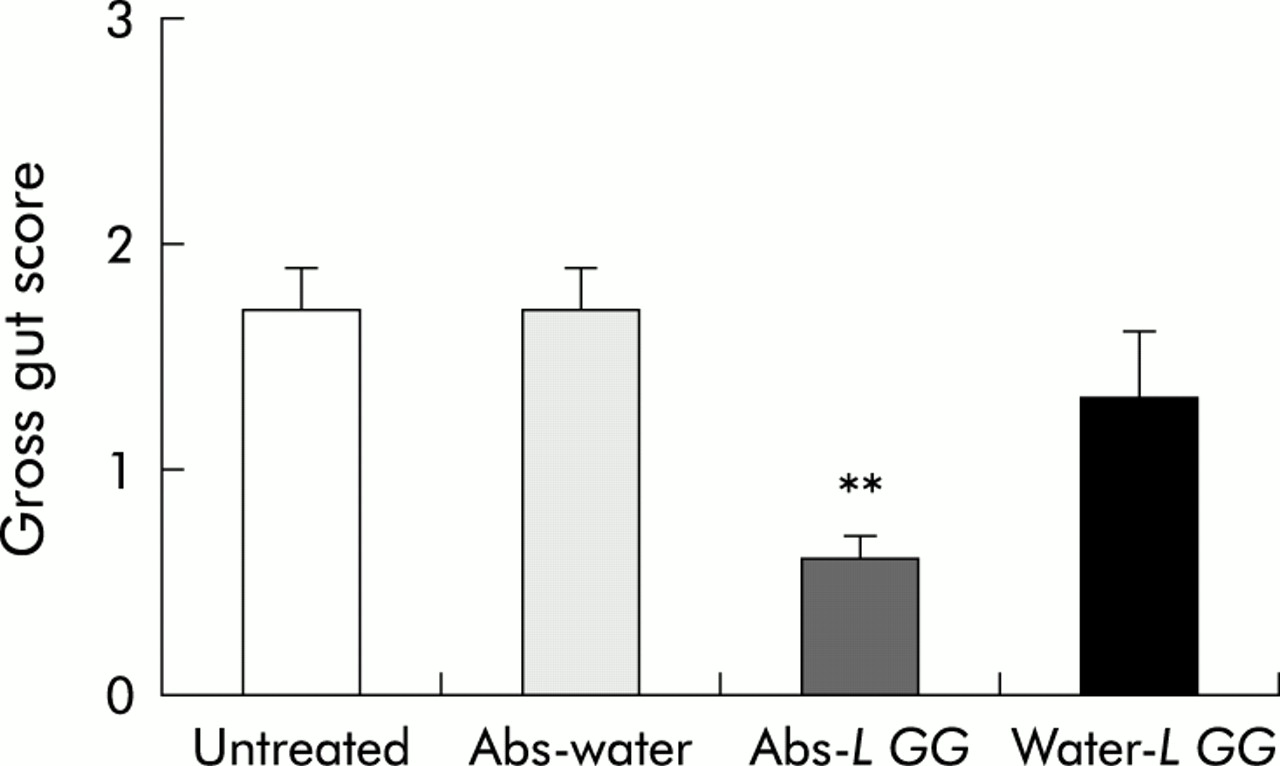
Gross gut scores (0–4) of caeca from specific pathogen free (SPF) transgenic (TG) rats treated with vancomycin/imipenem (Abs) or water followed by oral Lactobacillus rhamnosus GG (L GG) or water administration. Gross gut scores from untreated SPF TG rats are also given. Each group consisted of six rats. Data are expressed as mean (SEM). **p<0.01 versus untreated control rats.
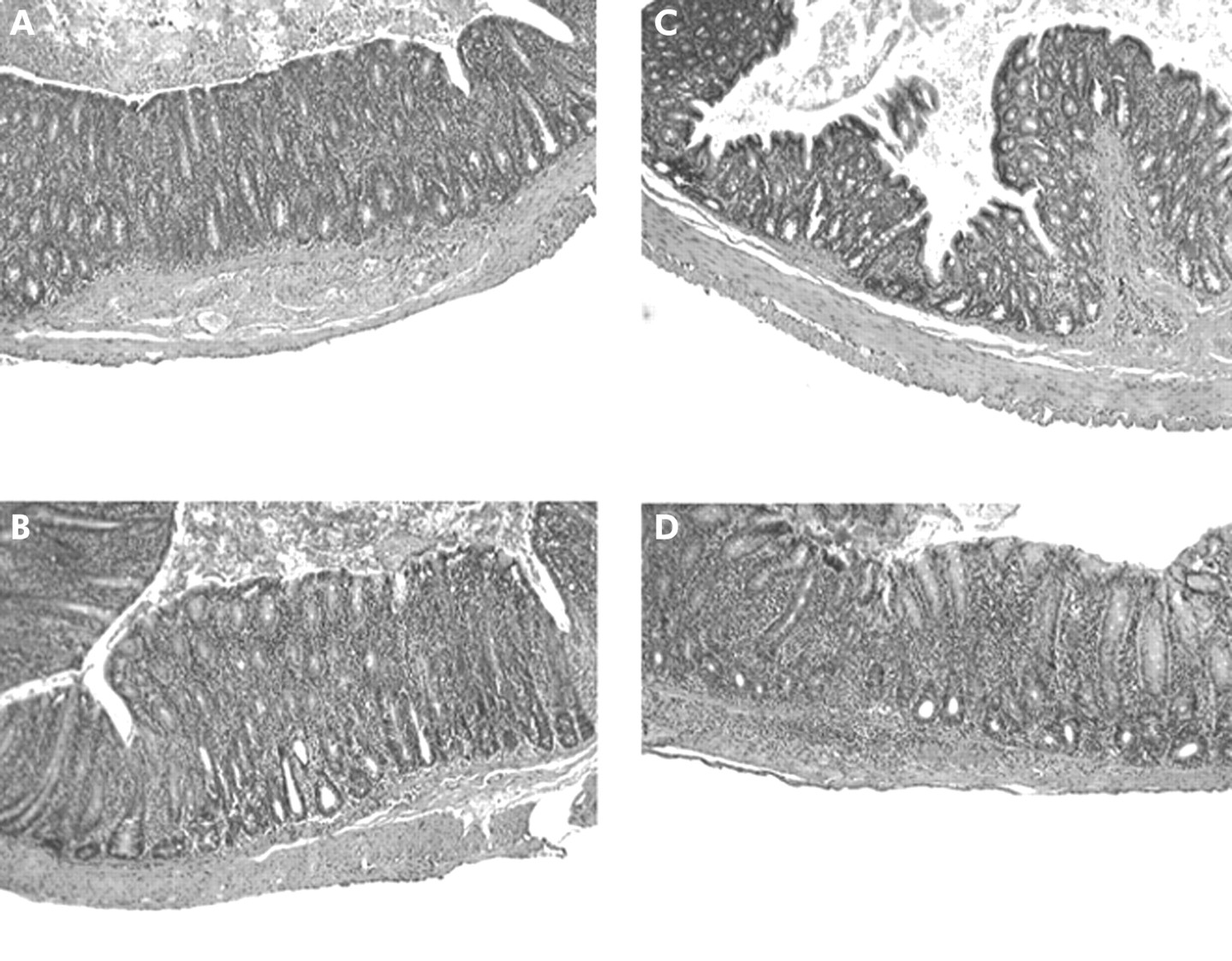
Representative photomicrographs of tissue sections (×40) from caeca of 16 week old specific pathogen free (SPF) transgenic (TG) rats which were treated with: (A) water; (B) two weeks oral vancomycin/imipenem followed by four weeks of water; (C) two weeks oral of vancomycin/imipenem followed by daily administration of Lactobacillus rhamnosus GG (L GG) over four weeks; and (D) two weeks of water followed by four weeks of oral L GG. Note the extensive mucosal and some submucosal inflammation as well as significant crypt hyperplasia in caeca in (A), (B), and (D). Only mild to modest mucosal inflammation was seen in TG rats treated with the combination of broad spectrum antibiotics followed by L GG (C).

Blinded total colonic inflammatory scores in specific pathogen free (SPF) transgenic (TG) rats treated for two weeks with oral broad spectrum antibiotics (Abs) or water followed by four weeks of Lactobacillus rhamnosus GG (L GG), Lactobacillus plantarum strain 299v (L pl), or water. Also, histology scores from SPF TG untreated rats are given. Values represent mean (SEM) of total histology scores (0–4). Vancomycin/imipenem followed by daily oral L GG administration significantly decreased total colonic histology scores versus other treatment groups. Feeding LP 299v did not prevent colitis relapse nor did it treat established colitis. *p<0.05 versus other treatment groups. Each group consisted of six SPF TG rats.
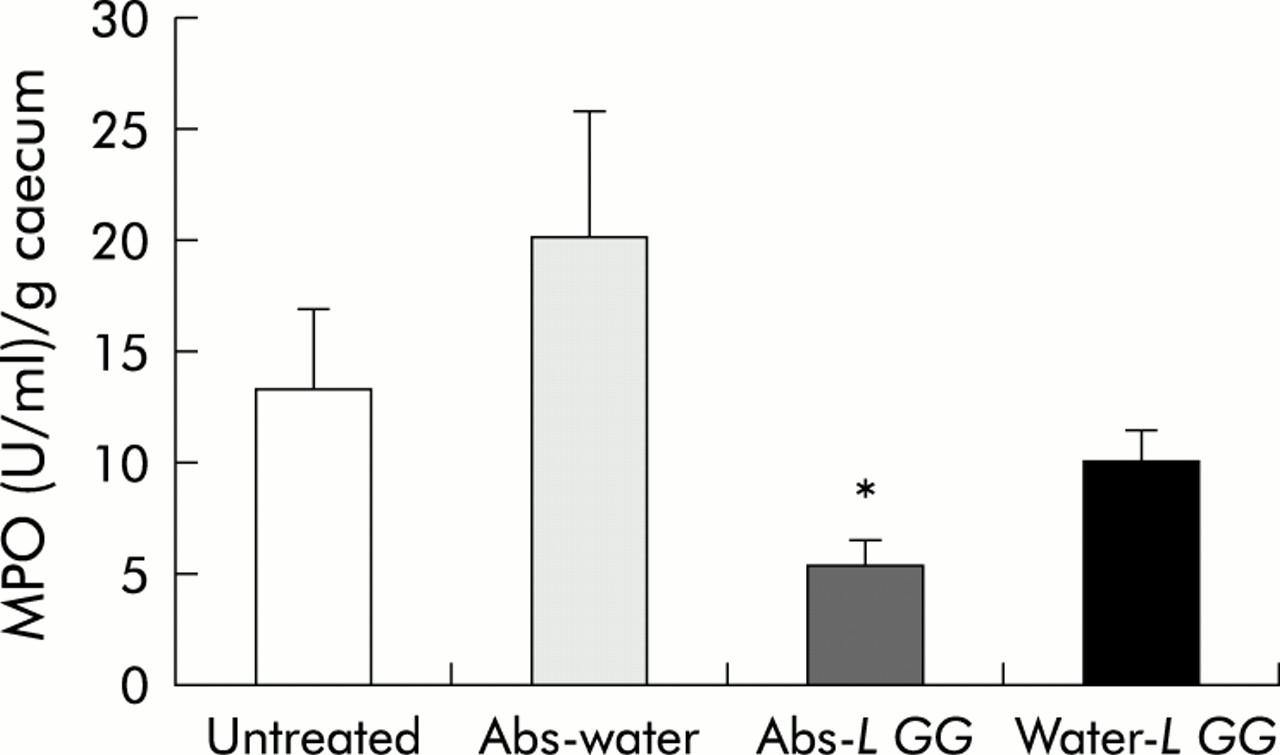
Caecal MPO values reflected the gross and histological scores; only TG rats treated with antibiotics followed by L GG showed a significant reduction in caecal MPO (5.6 (1.0) U/g tissue) versus all other treatment groups whereas MPO in caecal homogenates from TG rats treated with antibiotics alone (20.3 (5.6) U/g) or with L GG alone (10.2 (1.4) U/g) did not differ from untreated TG controls (13.3 (3.7) U/g) (fig 4). Caecal MPO from TG rats which had been fed L plantarum with or without antibiotic pretreatment did not differ significantly from other treatment groups (data not shown).

Caecal myeloperoxidase (MPO) concentrations in specific pathogen free (SPF) transgenic (TG) rats treated for two weeks with oral broad spectrum antibiotics (Abs) or water followed by four weeks of Lactobacillus rhamnosus GG (L GG), or water. Values from untreated TG control rats are also given. Data represent mean (SEM) MPO levels in units per gram of caecum. *p< 0.05 versus untreated controls.
Mucosal cytokines
Caecal homogenates from SPF TG rats treated with antibiotics and/or L GG contained significantly less IL-1β than the other treatment groups (fig 5A). A similar pattern was shown for caecal TNF, although TNF concentrations were about 10-fold less than caecal IL-1β (fig 5B). Interestingly, IL-10 concentrations were significantly increased in caecal homogenates from antibiotics-L GG treated TG rats versus other treatment groups whereas caecal TGF-β levels were no different among the groups (fig 5B).

Caecal interleukin (IL)-1β (A), and tumour necrosis factor (TNF), IL-10, and transforming growth factor β (TGF-β) (B) concentrations (pg/mg) from transgenic (TG) rats treated for two weeks with oral broad spectrum antibiotics (Abs) or water followed by four weeks of Lactobacillus rhamnosus GG (L GG) or water as well as from untreated controls. Values are mean (SEM). *p<0.05 versus untreated controls. The combination of antibiotics followed by L GG significantly decreased caecal IL-1β and TNF but also increased mucosal IL-10 concentrations.
Bacteriology of caecal contents
No differences were found in total bacterial counts of caecal contents between the antibiotic and/or L GG treatment groups in SPF TG rats. Also, concentrations of caecal anaerobic and aerobic bacteria, as determined by culture on Brucella and SBA plates, respectively, were not significantly different (data not shown). Oral administration of L GG resulted in a 10-fold increase in lactobacillus spp in caecal contents (1.0 (0.5)×107 CFU/g with antibiotics alone v 12 (3)×107 CFU/g caecal content in antibiotics-L GG treated rats). However, caecal bacteroides spp did not differ significantly among the treatment groups (2.0 (1.4)×1010 with antibiotics alone v 2.7 (0.3)×1010 CFU/g caecal content in the antibiotics-L GG treated group).
In vitro inhibition of bacterial adherence
B vulgatus adhered to IEC-6 cells after an incubation of three hours. However, coculture of 109B vulgatus and 109L GG per well significantly reduced the amount of adherent B vulgatus. No effect was noted with concentrations of 108/well (fig 6) or 107/well L GG (data not shown).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Dose dependent inhibition of in vitro adherence of 109 CFU B vulgatus/2 ml well to IEC-6 cells by Lactobacillus rhamnosus GG (L GG). Data represent concentrations (×103 CFU) of adherent B vulgatus after three hours of incubation with IEC-6 monolayers in the presence or absence of 108 or 109 CFU L GG/2 ml well. Values are mean (SEM). **p<0.01 versus B vulgatus control.
DISCUSSION
Broad spectrum antibiotics can prevent as well as treat established colitis in SPF B27 TG rats.11 Similar results were obtained in other experimental colitis models in rats and mice,11,26,27 emphasising the important role of intestinal bacteria for induction and perpetuation of chronic intestinal inflammation. Oral vancomycin/imipenem treatment resulted in undetectable levels of bacteroides spp, which are capable of selectively inducing colitis in B27 TG rats.10,13
We found that colitis relapsed within four weeks of stopping vancomycin/imipenem treatment of established colitis in SPF TG rats, resulting in disease relapse within four weeks with gross and histological scores, MPO, and proinflammatory cytokine levels nearly identical to untreated controls. This finding correlated with the reappearance of luminal bacteroides spp as caecal bacteroides concentrations were not significantly different in antibiotic treated rats versus untreated TG controls. These findings are in agreement with clinical observations in Crohn’s disease which recurs after stopping antibiotic treatment28 and can be explained by the reappearance of disease inducing intestinal bacteria in genetically susceptible hosts.
A clinically relevant finding of our study was that oral L GG treatment partially prevented relapse of colitis after antibiotic treatment whereas L GG administration alone failed to show a beneficial effect. This conclusion was supported by significantly decreased gross caecal inflammation and colonic histology scores, as well as a significant decrease in non-specific inflammatory markers and cytokines, such as caecal MPO, IL-1, and TNF in antibiotic-L GG treated SPF TG rats versus other treatment groups. This is the first report of an interactive effect of antibiotic and probiotic therapy in experimental colitis. These results are in agreement with Gionchetti et al in which relapsing pouchitis was prevented in 85% of refractory pouchitis with an oral probiotic cocktail VSL#3, which includes four strains of lactobacilli, three bifidobacteria species, and Streptococcus salivarius subsp thermophilus after the broad spectrum antibiotic rifaximin was stopped.20 Similarly, prospective treatment with rifaximin followed by VSL#3 could also prevent endoscopic postoperative recurrence of Crohn’s disease.29
Another major finding of our study was the specificity of prevention of relapsing colitis by various lactobacillus species, as demonstrated by the lack of efficacy of LP 299v in preventing colitis relapse in SPF TG rats after antibiotic treatment. However, this result is in contrast with our previous reports that oral LP 299v attenuated colitis in IL-10 KO mice,20 whereas L GG had no effect.30 A preventive effect of LP 299v was also shown in methotrexate induced enterocolitis in rats.31 These contrasting responses suggest that different lactobacilli may have variable host specificity or different efficacy in various inflammatory conditions, indicating that not all probiotics are equally protective in chronic experimental colitis. The concept of selective dependency of beneficial effects of a probiotic bacterial species on the species and genetic background of the host is similar to that of disease inducing bacteria.10,13,32,33
Other studies have reported primary beneficial effects of probiotics in the treatment or prevention of human or experimental intestinal inflammation. VSL#3 was able to treat established colitis in IL-10 KO mice.25 The same cocktail has also shown efficacy in an open labelled study on maintenance therapy of ulcerative colitis.34L GG can prevent relapsing Clostridium difficile diarrhoea,19 traveller’s diarrhoea35 and rotavirus infections in children.36 The non-pathogenic E coli strain Nissle was shown to be as effective as low dose mesalamine in maintaining remission of ulcerative colitis.37,38
The protective mechanisms of probiotic bacteria are still relatively unknown, with postulated effects on luminal microecology, mucosal barrier function, and immunoregulation. In our study there was no decrease in the number of luminal bacteroides spp, raising the possibility that some of the protective effects of antibiotics-L GG could be explained by immunomodulation. Not only does this therapy result in decreased caecal proinflammatory cytokines IL-1β and TNF, but it also increased mucosal IL-10. Lack of stimulation of mucosal IL-10 by L GG or antibiotics alone indicate that neither treatment is responsible for induction of IL-10. It is possible that a combination of antibiotics and probiotics alters the balance of enteric bacterial components, leading to altered mucosal cytokine profiles. IL-10 is an immunoregulatory cytokine, mainly produced by macrophages, dendritic cells, and T regulatory 1 lymphocytes. However, we recently reported that in caecal bacterial lysate stimulated MLN cells of B27 TG rats as well as their non-TG littermates, IL-10 is primarily produced by B cells.39 In vitro, lysates of VSL#3 stimulate IL-10 production and diminish TNF secretion by rat splenocytes stimulated with caecal bacteria.40 In preliminary data, it was demonstrated that DNA extracts of VSL#3 induce IL-10 and exert protective immune responses.41 IL-10 can inhibit antigen specific proliferation and cytokine secretion by Th1 lymphocytes and has downregulatory effects on macrophages and dendritic cells, such as suppression of activation and IL-12 production.42,43 IL-10 can also prevent interferon γ induced disruption of colonic epithelial barriers.44 Defective in vivo production of IL-10 results in spontaneous colitis in IL-10 KO mice in the presence of normal intestinal bacteria.7,45 Furthermore, oral administration of Lactococcus lactis, engineered to produce high colonic IL-10 levels, prevents the onset of colitis in IL-10 KO mice as well as in dextran sodium sulphate induced colitis.46 Thus stimulation of colonic IL-10 production by the combination of antibiotic use and L GG colonisation could have a variety of beneficial effects on mucosal immunoregulation and barrier function.
Increased mucosal IL-10 production and decreased levels of the proinflammatory cytokines IL-1β and TNF by the combination of antibiotics and L GG in our study are in agreement with the findings of Ulisse et al who showed increased IL-10 as well as decreased TNF and IFN-γ in pouch biopsies of refractory pouchitis patients treated with VSL#3 after antibiotic pretreatment.47 Similar to our findings, these authors found that tissue IL-10 levels were unchanged in inflamed pouches and were not affected by prior antibiotic treatment.47 However, in IL-10 KO mice the probiotic cocktail VSL#3 treated colitis with a concomitant decrease of mucosal IFN-γ and TNF secretion,21 indicating that the beneficial effects of probiotics in colitis can be mediated by factors other than IL-10 induction.
Several non-immune protective mechanisms have been described for probiotic bacterial species. These organisms can exert growth inhibitory and bactericidal activities on enterotoxigenic E coli.48 However, in our study caecal concentrations of the disease inducing bacteroides spp were not significantly decreased. Another potential protective non-immune mechanism involves inhibition of epithelial adherence of disease inducing organisms. Intestinal bacteria compete with other organisms for a limited number of epithelial receptors. Increased amounts of colonic mucosal associated bacteria were found in IL-10 KO mice28 as well as in mucosal biopsies from IBD patients.49 Madsen et al also reported a relative lack of adherent lactobacilli in IL-10 KO mice18 and restoration to normal levels after probiotic treatment.19,21 Our in vitro coculture studies demonstrated that L GG decreased adherence of B vulgatus to rat IEC-6 cells only at high concentrations of 109 CFU/ml, with no effects at lower doses. This result is in agreement with in vitro studies by Madsen and colleagues21 and Mack and colleagues25 in which probiotic bacteria prevented the epithelial adherence of Salmonella dublin or enteropathogenic E coli. However, in our study, in vivo co-association of B vulgatus and L GG did not prevent colitis in gnotobiotic TG rats. This lack of effect by L GG in the dual association model could be explained by the overwhelming colonisation capacity of the disease inducing B vulgatus versus L GG, which were 100-fold more frequent than L GG. It is possible that at this relatively low in vivo luminal concentration L GG would not be able to mediate protection by inhibiting epithelial adherence of B vulgatus, as equal concentrations of L GG were required to interfere with B vulgatus epithelial adherence. Based on these findings we postulate that protective effects by probiotics in SPF TG rats could only be achieved by pretreatment of rats with vancomycin/imipenem, which would result in a significant decrease in B vulgatus11 before L GG treatment. However, despite the prevention of colitis relapse by the combination treatment of vancomycin/imipenem followed by L GG, this treatment did not significantly reduce luminal bacteroides spp, even though increased caecal lactobacilli spp was shown by caecal culture. Taken together, reduction of luminal bacteroides spp could not explain the protective effects of broad spectrum antibiotics followed by L GG. Prevention of colitis relapse as well as immunomodulatory effects were shown only with the combination treatment of vancomycin/imipenem followed by L GG. It is possible that the combination treatment promoted synergistic interaction of L GG with other endogenous probiotic species, resulting in disease protection. The absence of these potential beneficial interactions with other intestinal protective bacteria in gnotobiotic TG rats co-associated with L GG and B vulgatus could also explain the lack of efficacy of L GG in the prevention of colitis. Alternatively, lack of exposure to lactic acid bacteria early in life in these gnotobiotic rats may prevent stimulation of protective responses by L GG at an older age.
In summary, oral L GG can prevent relapse of colitis in SPF B27 TG rats after antibiotics are stopped whereas oral administration of L plantarum did not have any effect, demonstrating the selective protective effects of two lactobacillus species. The beneficial in vivo effects of L GG following antibiotics are possibly the result of enhanced IL-10 production. This is the first study in experimental colitis in which antibiotics and probiotics display synergistic in vivo effects. Restoring the microbial balance between detrimental and protective luminal bacteria by combining antibiotic and probiotic approaches may be the most physiological approach to treat IBD and may alter the natural history of these chronic relapsing diseases.
Acknowledgments
This study was supported by NIH grants KO8 553773, RO3 558666, DK 40249, DK 34987, and by the Crohn’s and Colitis Foundation of America. The authors wish to thank Charlotte Walters of the ImmunoTechnogy Core of the Center for Gastrointestinal Biology and Disease for technical assistance.