Article Text

Abstract
Objective Liver tumour-initiating cells (T-ICs) are critical for hepatocarcinogenesis. However, the underlying mechanism regulating the function of liver T-ICs remains unclear.
Methods Tissue microarrays containing 242 hepatocellular carcinoma (HCC) samples were used for prognostic analysis. Magnetically activated cell sorting was used to isolate epithelial cell adhesion molecule (EPCAM)-positive cells. The gene expressions affected by miR-429 were determined by arrays. Co-immunoprecipitation was used to study interactions among retinoblastoma protein (RB1), Rb binding protein 4 (RBBP4) and E2F transcription factor 1 (E2F1). The DNA methylation status in CpG islands was detected by quantitative methylation analysis. miRNAs in microvesicles were isolated by a syringe filter system.
Results The significant prognosis factor miR-429 was upregulated in HCC tissues and also in primary liver T-ICs isolated from clinical samples. The enrichment of miR-429 in EPCAM+ T-ICs contributed to hepatocyte self-renewal, malignant proliferation, chemoresistance and tumorigenicity. A novel functional axis involving miR-429, RBBP4, E2F1 and POU class 5 homeobox 1 (POU5F1 or OCT4) governing the regulation of liver EPCAM+ T-ICs was established in vitro and in vivo. The molecular mechanism regulating miR-429 expression, involving four abnormal hypomethylated sites upstream of the miR-200b/miR-200a/miR-429 cluster, was first defined in both EPCAM+ liver T-ICs and very early-stage HCC tissues. miR-429 secreted by high-expressing cells has the potential to become a proactive signalling molecule to mediate intercellular communication.
Conclusions Epigenetic modification of miR-429 can manipulate liver T-ICs by targeting the RBBP4/E2F1/OCT4 axis. This miRNA might be targeted to inactivate T-ICs, thus providing a novel strategy for HCC prevention and treatment.
- Hepatocellular Carcinoma
- Stem Cells
- Signal Transduction
Statistics from Altmetric.com
Significance of this study
What is already known about this subject?
-
Liver tumour-initiating cells are critical to hepatocarcinogenesis.
-
Liver tumour-initiating cells can be identified by several cell-surface antigens such as epithelial cell adhesion molecule (EPCAM) and PROM1 (CD133).
-
RBBP4 and E2F1 are physically associated in the presence of RB1.
-
miR-429, miR-200b and miR-200a are clustered on Chr1 and have similar seed sequences.
What are the new findings?
-
The significant prognosis factor miR-429 was upregulated in hepatocellular carcinoma (HCC) tissue and also in primary liver tumour-initiating cells (T-ICs) isolated from clinical HCC samples. The enrichment and internalisation of miR-429 in EPCAM+ T-ICs were shown, contributing to hepatocyte self-renewal, malignant proliferation, chemoresistance and tumorigenicity.
-
A novel functional axis involving miR-429/Rb binding protein 4 (RBBP4)/E2F transcription factor 1 (E2F1)/POU class 5 homeobox 1 (POU5F1 or OCT4) for the manipulation or conversion of the characteristics of liver EPCAM+ T-ICs was established and verified in vitro and in vivo. Stemness gene OCT4 was identified as the new candidate gene downstream of the RBBP4/E2F1 signal pathway which was directly regulated by miR-429.
-
The molecular mechanism of abnormal miR-429 expression involving four hypomethylated sites upstream of the miR-200b/miR-200a/miR-429 cluster were first defined in both EPCAM+ liver T-ICs and very early-stage clinical HCC tissue.
-
Secreted miR-429, packaged in microvesicles (MVs), has the potential to become a proactive signalling molecule that mediates intercellular communication.
How might it impact on clinical practice in the foreseeable future?
-
Our findings provide a new insight into the molecular mechanism of hepatocarcinogenesis which may help to establish a promising strategy for the prevention and treatment of HCC.
Introduction
Our growing understanding of cancer has made it clear that tumour-initiating cells (T-ICs) play a critical role in the development of tumours. The existence of T-ICs was first observed in acute myeloid leukaemia.1 Human epithelial cell-derived cancers have highly heterogeneous morphologies, suggesting that T-ICs, which are believed to give rise to tumour heterogeneity, may be especially important for the tumorigenesis of epithelial cell-derived cancers. T-ICs have also been detected in several human epithelial cell-derived tumours including colon, breast, prostate and pancreas cancers.2–5 Eradicating the T-ICs of a tumour is essential to achieve more stable remission or to completely eliminate the cancer.6 ,7 Further investigation of T-ICs is needed to achieve better clinical outcomes for patients with tumours.
Hepatocellular carcinoma (HCC) is among the 10 most deadly cancers worldwide and is particularly prevalent in China. Increasing evidence indicates that liver T-ICs are critical to hepatocarcinogenesis.8–11 Liver T-ICs can be identified by several cell-surface antigens such as PROM1 (CD133),8 THY1 (CD90),9 epithelial cell adhesion molecule (EPCAM)10 and CD24.11 Advances in our knowledge of HCC and liver T-ICs have generated considerable interest in studying the underlying mechanisms regulating the function of liver T-ICs.
miRNAs are an abundant class of small non-coding RNAs that inhibit translation or induce mRNA degradation in general by binding to the 3′UTR of target mRNAs. miRNAs have been identified as tumour promoters or suppressors, regulating the progression of cancers and T-ICs.12 ,13 miR-429, a member of an evolutionarily conserved family of miRNAs that includes miR-200c, miR-141, miR-200b and miR-200a, is expressed in various epithelial tissues.14 miR-429 plays contrasting roles in a wide range of common epithelial cancers. It is downregulated in human gastric carcinoma tissue15 and malignant myoepithelioma of the breast.16 Overexpression of miR-429 induces the mesenchymal-to-epithelial transition in metastatic ovarian cancer cells, and low-level expression of miR-200b-429 predicts poor outcomes.17 ,18 In contrast, miR-429 is upregulated in human colorectal cancer tissues and is significantly associated with tumour size, lymph node metastasis and poor prognosis.19 The increased expression of miR-429 has also been identified in endometrial adenocarcinoma and infiltrating bladder urothelial carcinoma, and plays an important role in tumorigenesis.20 ,21 A significant induction of miR-429 has been found in highly metastatic pancreatic ductal adenocarcinoma cell lines.22 In addition to being specifically expressed during the differentiation of human embryonic stem cells,23 miR-429 is differentially expressed in tumour stem cells such as pancreatic cancer stem cells.24 However, the expression and role of miR-429 in HCC, one of the most common epithelial cancers, and in liver T-ICs have not been clearly demonstrated.
In this study we identified a novel mechanism regulating the function of liver T-ICs. We found that epigenetic modification of miR-429 can promote the properties of liver T-ICs by targeting the Rb binding protein 4 (RBBP4)/ E2F transcription factor 1 (E2F1)/POU class 5 homeobox 1 (POU5F1 or OCT4) axis, implying a novel strategy for the prevention and treatment of HCC based on targeting miR-429 and its downstream genes.
Methods
Patients, specimens and tissue microarrays
All samples used in this study were obtained from patients undergoing surgery for HCC or hepatic haemangioma at the Eastern Hepatobiliary Surgery Hospital (Shanghai, China). Patient samples were obtained following informed consent according to an established protocol approved by the Ethics Committee of Eastern Hepatobiliary Surgery Hospital. A total of 161 HCC tissue specimens and 48 normal liver tissue specimens were collected immediately after resection for RNA isolation and miR-429 quantification. Normal liver tissue was collected from the distal normal liver section of patients undergoing surgery for liver haemangioma who had no evidence of chronic liver disease. The use of normal liver tissue as a negative control can exclude the possible impact of HCC mass on adjacent liver tissues. Chronic hepatitis type B (n=24) and liver cirrhosis tissues (n=22) were obtained from the distal liver tissue of patients undergoing surgery for liver haemangioma who showed evidence of chronic hepatitis type B or HBV-related cirrhosis, respectively. No patients received any preoperative anticancer treatment. Another tissue microarray containing 242 HCC cancer tissues was used to analyse the correlation between miR-429 expression and the clinicopathological features of patients with HCC. Overall survival was the interval between the dates of surgery and death. Disease-free survival was the interval between the dates of surgery and recurrence; if recurrence was not diagnosed, patients were classified on the date of death or the last follow-up.
Further details of the materials and methods used in this study can be found in the online supplement.
Results
miR-429 is enriched in HCC tissues and is a significant prognosis factor
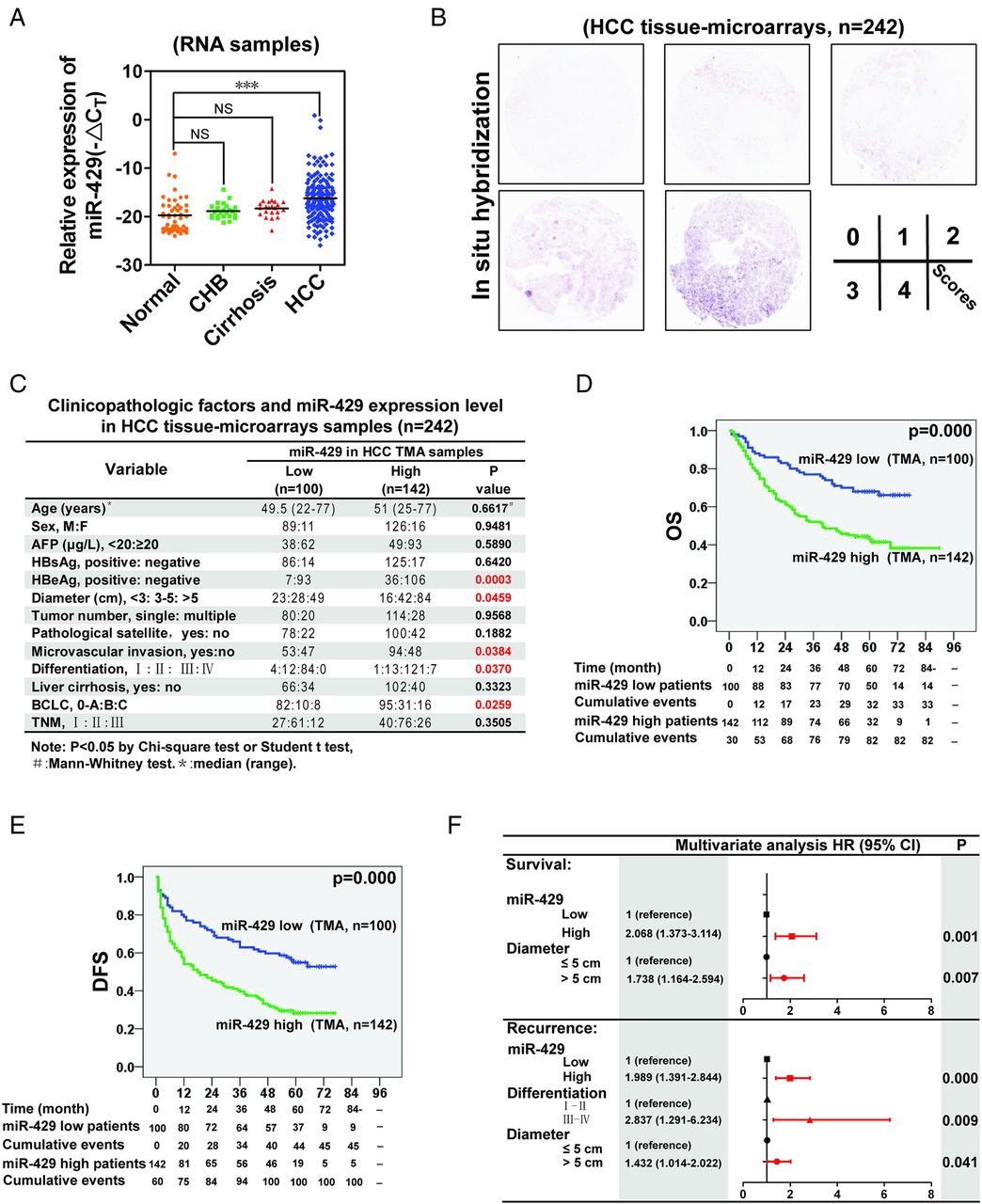
First, we investigated the expression of miR-429 in fresh RNA samples of normal liver, chronic hepatitis type B, liver cirrhosis and HCC tissues. We observed miR-429 enrichment in clinical tumour tissues compared with other liver sections (figure 1A and online supplementary table S1). To determine the relationship between miR-429 and HCC clinicopathological characteristics, we next performed in situ hybridisation of miR-429 in HCC tissue microarrays (n=242; figure 1B and online supplementary table S2). Note that the expression of miR-429 in fresh RNA samples detected by quantitative RT-PCR is positively correlated with that in paraffin-embedded samples determined by in situ hybridisation (n=161), which underlines the fact that the classification of miR-429 by these two methods in the same patients achieves similar results (see online supplementary table S3). Correlation regression analysis indicated that high miR-429 expression was associated with multiple malignant characteristics of HCC (n=242 for tissue microarrays and n=161 for RNA samples; figure 1C and online supplementary table S4). Kaplan–Meier survival analysis showed that miR-429 expression levels were negatively correlated with overall survival and disease-free survival (figure 1D,E). Thirteen survival- and recurrence-related clinicopathological variables were analysed by univariate analysis (see online supplementary table S5), which showed that HBeAg, tumour diameter, tumour number, microvascular invasion, differentiation and miR-429 expression level were statistically correlated with both survival and recurrence (see online supplementary figures S1A,B). Each individual parameter was further subjected to multivariate Cox proportional hazards analysis, which indicated that tumour diameter and the miR-429 expression level were independent and significant factors affecting the survival of patients with HCC (figure 1F). Importantly, miR-429 had the greater HR for survival.

miR-429 is a prognostic factor of hepatocellular carcinoma (HCC). (A) The relative expression of miR-429 was determined in fresh RNA samples of normal liver (distal non-cancerous tissues away from liver haemangioma, n=48), chronic hepatitis type B (CHB, n=24), liver cirrhosis tissue (n=22) and HCC tissue (n=161). (B) Representative staining for miR-429. (C) Associations between miR-429 expression and clinicopathological characteristics. Tissue microarrays (n=242). Scores 0 and 1 defined as low expression, scores 2–4 defined as high expression. (D,E) Kaplan–Meier analysis of the correlation between miR-429 expression levels and overall survival (OS) or tumour recurrence in patients with HCC. (F) Multivariate analysis of HRs for overall survival and tumour recurrence. ***p<0.001; NS, not significant. DFS, disease-free survival.
Enrichment of miR-429 in liver T-ICs
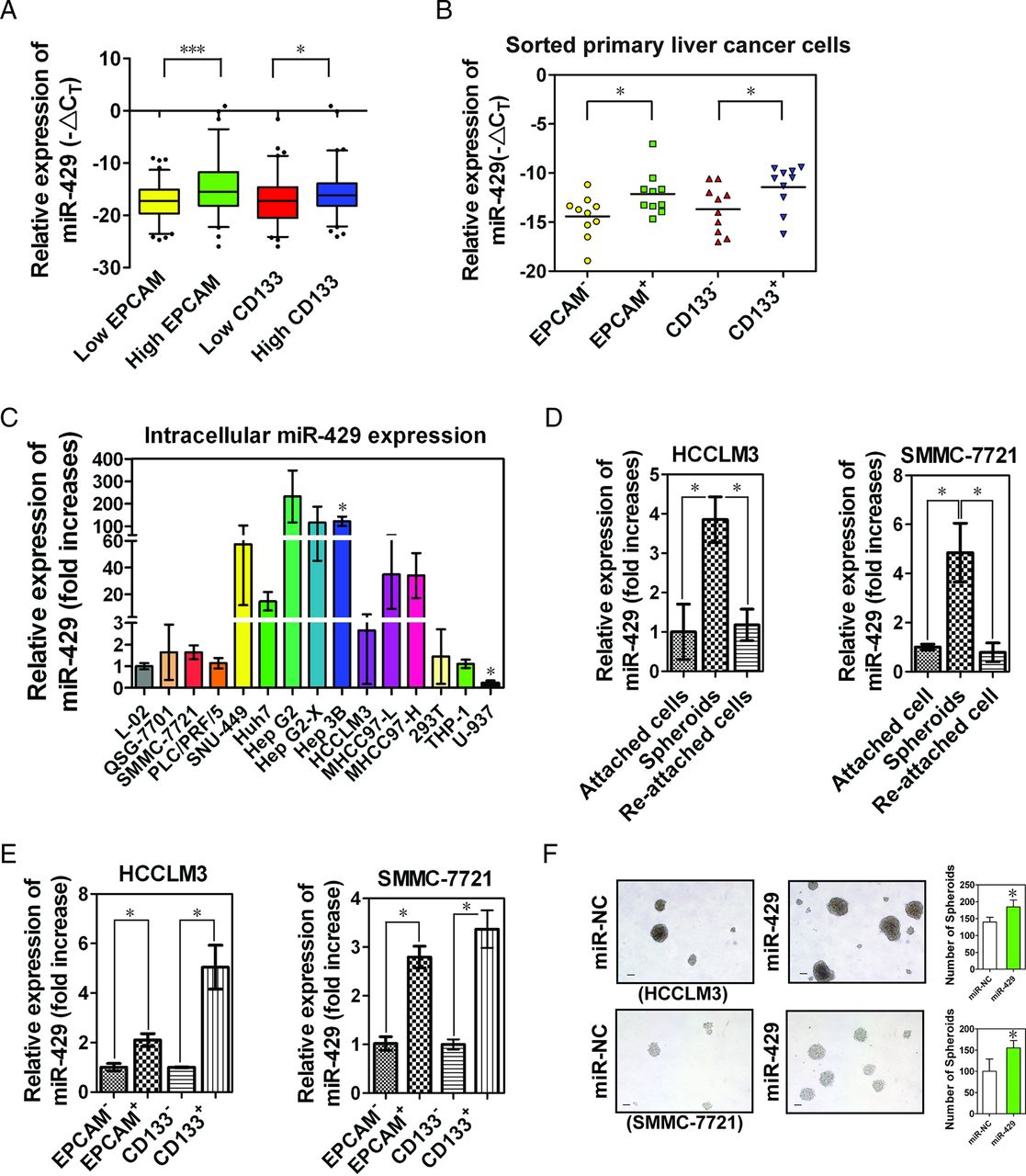
Stem-like cells within tumours (ie, T-ICs) are probably a unique subpopulation of tumour cells responsible for the initiation and pathogenesis of cancer.25 Liver T-ICs are characterised by several cell marker molecules such as EPCAM and PROM1 (CD133).8 ,10 We further examined the expression of miR-429 in high EPCAM- or high CD133-expressing HCC tissues. As shown in figure 2A, miR-429 showed significantly elevated expression in high EPCAM and high CD133 HCC tissues. Furthermore, we observed increased miR-429 levels in sorted primary EPCAM+ or CD133+ liver cancer cells relative to matched EPCAM− or CD133− cells (figure 2B and see online supplementary figure S2A).

miR-429 is enriched in liver tumour-initiating cells (T-ICs). (A) Box-and-whisker plots of miR-429 expression data were constructed with different expression levels of epithelial cell adhesion molecule (EPCAM) or CD133 in fresh hepatocellular carcinoma (HCC) RNA specimens (n=161). The end of the whiskers was set to 5–95 percentiles. Outliers were plotted as individual points. The average expression level was used as the cut-off to define the low and high expression group. (B) The expression of miR-429 was determined in magnetically sorted primary EPCAM+ or CD133+ cells in clinical specimens of HCC (n=10). (C) Intracellular expression of miR-429 was determined by TaqMan quantitative RT-PCR. The results are shown as mean±SD (n=3). *p<0.05 vs intracellular miR-429 expression in L02 cells. (D) Expression of miR-429. A total of 3000 cells were seeded into low-adhesion plates to collect undifferentiated spheroids. (E) Expression of miR-429 in magnetically sorted HCC cells. (F) Comparison of spheroid formation. After transfected with 100 nM miR-429 mimics or negative control mimics, a total of 3000 cells were seeded into low-adhesion plates and the number and size of the spheroids was examined 10 days after incubation. *p<0.05, ***p<0.001.
As expected, the enhanced expression of miR-429 was observed in most HCC cell lines compared with normal liver cell lines (L-02, QSG-7701), 293 T cells and immune cell lines (THP-1, U-937) (figure 2C). We chose the lower miR-429-expressing HCC cell lines HCCLM3 and SMMC-7721 to perform gain of function experiments. T-ICs are enriched in non-attached spheroids and will differentiate after reseeding under normal adherent conditions.26 As expected, the expression of miR-429 was increased together with most of the stem cell-associated genes (see online supplementary figure S2B) in undifferentiated spheroids (see online supplementary figure S2C; differentiation status was determined by the expression of hepatocyte differentiation-related genes including ALB, G-6-P, ALDOB, KRT8, AFP, KRT19 and FN1), and decreased after reseeding into adherence-promoting plates (figure 2D). Similar results were also obtained in magnetically sorted EPCAM+ or CD133+ HCC cells (figure 2E), suggesting a putative role of miR-429 in liver T-ICs.
As shown in online supplementary figure S3A and figure 2F, miR-429-transduced HCC cells formed larger and more numerous spheroids than control cells treated with negative-control (NC) mimics alone. miR-429-transduced HCC cells also showed higher levels of stem cell-associated genes (see online supplementary figure S3B) and an increased percentage of the EPCAM+ but not the CD133+ T-IC subpopulation (see online supplementary figure S3C). In addition, we observed a positive correlation between EPCAM (but not CD133) and miR-429 expression in both HCC cell lines and clinical samples (see online supplementary figures S3D,E), suggesting that miR-429 may promote the properties of EPCAM+ liver T-ICs.
miR-429 promotes the properties of EPCAM+ liver T-ICs
To determine the effect of miR-429 on the properties of liver T-ICs, magnetically sorted EPCAM− or EPCAM+ HCCLM3 cells were treated with miR-429 mimics or an antisense inhibitor of miR-429 (miR-429 antagomir). The introduction of antagomir-429 into the EPCAM+ HCCLM3 cells (see online supplementary figure S4A) led to a significantly decreased proportion of EPCAM+ but not CD133+ cells (figure 3A and see online supplementary figure S4B), blockage of malignant cell proliferation/cell cycle (figure 3B), inhibition of tumour spheroid growth (figure 3C), reduction of T-ICs (figure 3D and see online supplementary figure S4C) and amelioration of chemotherapy resistance (figure 3E) compared with NC or mutant antagomir-429-treated cells. Downregulation of miR-429 in HepG2 cells, which highly express miR-429, also resulted in decreased spheroid formation and impaired expression of stem cell-associated genes (see online supplementary figures S4D,E). Conversely, exogenous expression of miR-429 in EPCAM− HCCLM3 cells promoted malignant proliferation, self-renewal, chemoresistance and increased the proportion of T-ICs in vitro compared with miR-NC or miR-429mut mimic groups (figures 3A–E and online supplementary figures S4A–C). Subcutaneous injection of miR-429-expressing EPCAM− HCCLM3 cells resulted in an increased incidence of tumour formation, even when only 5000 cells were implanted (figure 3F). Moreover, blockage of miR-429 expression by miR-429 antagomir significantly retarded tumorigenesis and decreased the tumour volume in EPCAM+ HCCLM3 cells (figure 3F and online supplementary figure S4F). We then used Huh7 cells, which have intermediate levels of miR-429 (figure 2C), to perform gain and loss of function experiments. As expected, similar results were obtained in the presence or absence of miR-429 in Huh7 cell lines (see online supplementary figures S5A–D), which further verified the universal effect of miR-429 on the properties of liver T-ICs.
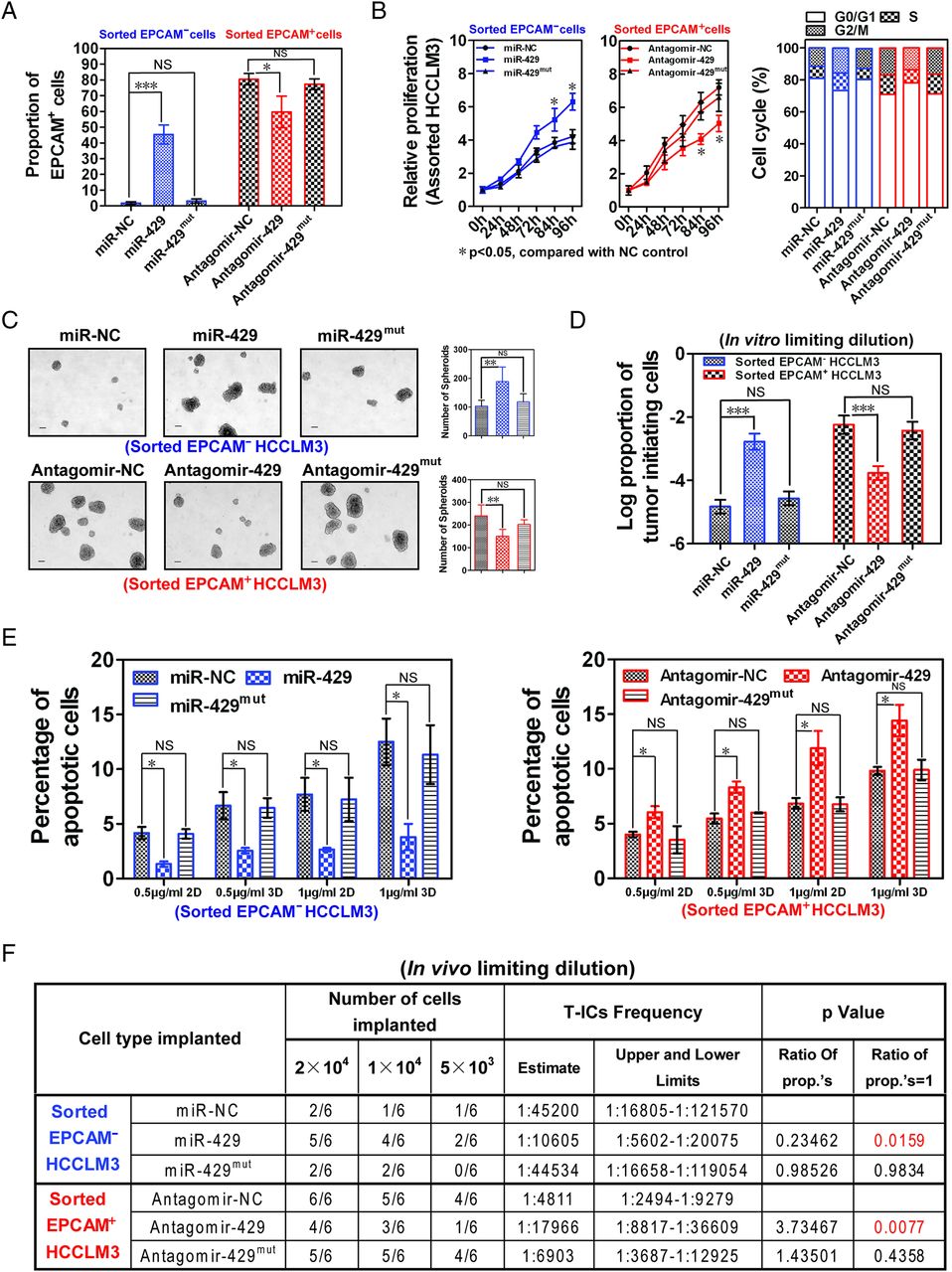
miR-429 manipulates the properties of epithelial cell adhesion molecule-positive (EPCAM+) liver tumour-initiating cells (T-ICs). (A) The percentage of EPCAM+ cells was determined by flow xytometry 4 days after magnetic sorting of EPCAM− or EPCAM+ HCCLM3 cells in which miR-429 was restored (miR-429 mimics: 100 nM) or inhibited (antagomir-429: 40 μM), respectively. Data are shown from three independent experiments. Blue represents treated EPCAM− cell group; red represents treated EPCAM+ cell group. (B) Proliferation was evaluated by a CCK8 assay. The cell cycle was calculated by flow cytometry 4 days after the indicated treatment. (C) Determination of tumour spheroid formation. A total of 3000 cells were seeded into low-adhesion plates and incubated for 10 days after the indicated treatment. (D) The proportion of T-ICs was evaluated by a limiting dilution assay 10 days after the indicated treatment. (E) Apoptosis was determined by flow cytometry 2 or 3 days after treatment with 0.5 μg/mL or 1 μg/mL cisplatin. (F) Male non-obese, diabetic, severe combined immunodeficient (NOD-SCID) mice (n=6) were subcutaneously injected with the indicated number of EPCAM− or EPCAM+ HCCLM3 cells in which miR-429 was restored (miR-429 mimics: 100 nM) or inhibited (antagomir-429: 40 μM) before being implanted and the proportion of T-ICs was evaluated 6 weeks after injection by a limiting dilution assay. ***p<0.001, **p<0.01, *p<0.05; NS, not significant.
miR-429 directly targets RBBP4
As miRNAs function through inhibition of downstream target genes, we further analysed potential target genes of miR-429 by identifying the shared genes among three groups: (1) downregulated genes in EPCAM− HCCLM3 cells treated with miR-429 mimics; (2) upregulated genes in EPCAM+ HCCLM3 cells treated with antagomir-429; and (3) genes predicted as potential miR-429 targets (http://www.targetscan.org). Figure 4A shows the 13 genes shared by these three groups. We then examined the expression of these 13 potential target genes in four HCC cell lines in the presence of miR-429 mimic or antagomir. As shown in online supplementary figure S6A, a negative correlation between miR-429 and SLC38A4, RBBP4, ACVR1C, EIF5B, EDEM3 or RASSF8 was observed in at least three cell lines, suggesting that microarrays presented a believable result, and these six genes are verified target genes of miR-429 in multi-HCC cell lines. In comparison with the other five target genes, only the restraint (see online supplementary figure S6B) of RBBP4 resulted in a significant upregulation of the proportion of EPCAM+ T-ICs, which suggests that miR-429 manipulates liver T-ICs mainly through targeting RBBP4 but not other genes (see online supplementary figure S6C). RBBP4, also known as RbAp48 or NURF55, is a tumour suppressor protein. It mediates the function of the RB1 by forming an RB1/RBBP4-associated histone deacetylase complex and is important for human stem cell maintenance.27–29 However, little is known about its potential role during the pathogenesis of HCC. We found lower levels of RBBP4 in tumour tissues compared with normal tissues, and an inverse correlation between the expression of miR-429 and that of RBBP4 mRNA in HCC samples and most HCC cell lines (see online supplementary figures S7A–C). As expected, we found a markedly lower expression of RBBP4 in undifferentiated spheroids and EPCAM+ HCC cells (figure 4B,C and online supplementary figure S7D). Western blot and luciferase reporter assays containing a wild-type or mutant 3′UTR of RBBP4 further indicated the direct regulation of RBBP4 by miR-429 (figure 4D,E).
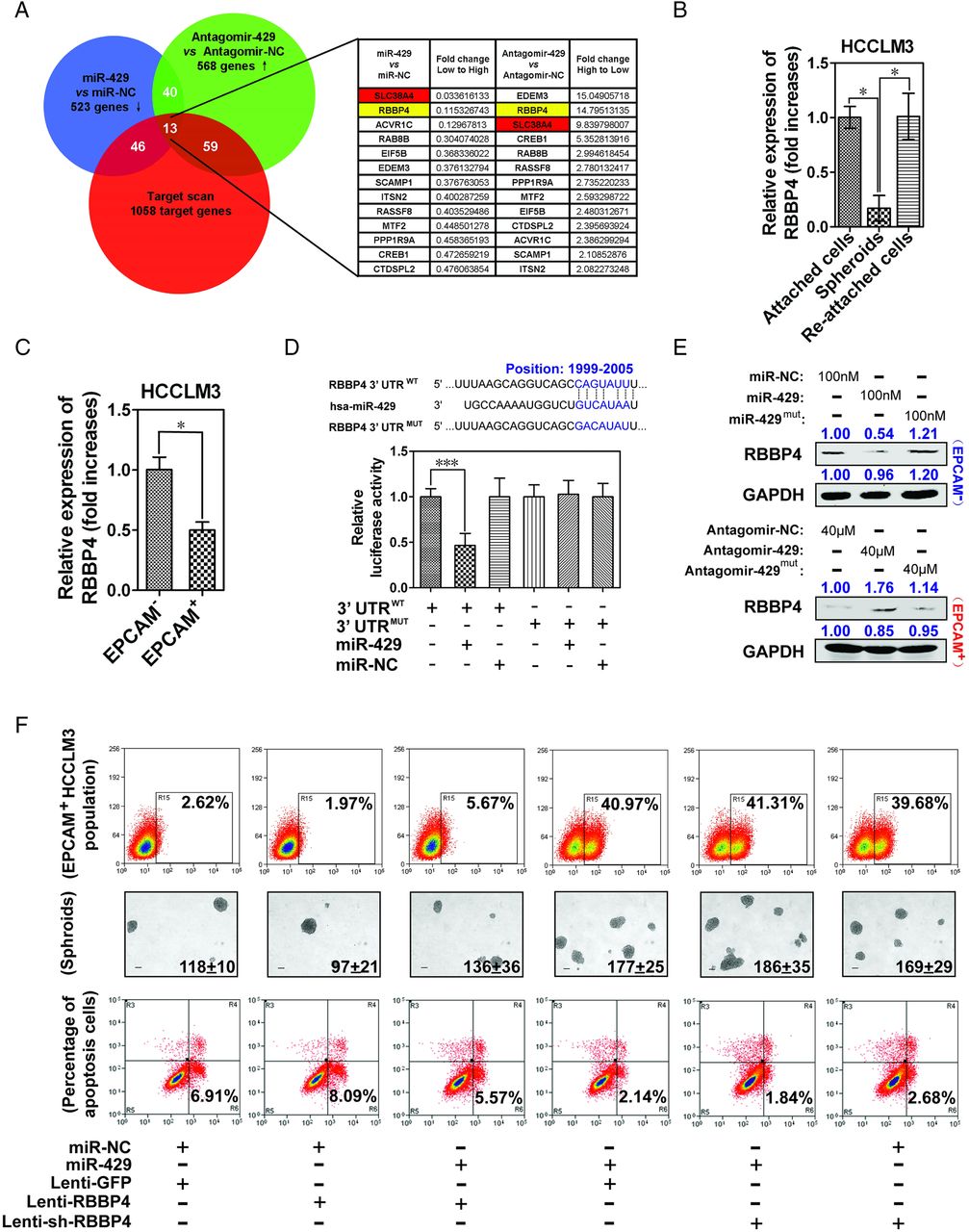
RBBP4 is a direct target gene of miR-429. (A) Magnetically sorted epithelial cell adhesion molecule-negative (EPCAM−) and positive (EPCAM+) HCCLM3 cells were treated with 100 nM miR-429 mimics or 40 μM antagomir-429, respectively, for 4 days. The gene expression profiles were determined by an Affymetrix Human U133 plus2.0 array. (B) Levels of RBBP4 mRNA in attached cells, spheroids and re-attached HCCLM3 cells. (C) Levels of RBBP4 mRNA in magnetically sorted EPCAM+ and EPCAM− HCCLM3 cells. (D) Putative miR-429-binding site in the RBBP4 3′UTR. Luciferase assays with reporter constructs comprising the wild-type or mutant RBBP4 3′UTR downstream of a luciferase gene were performed after co-transfection with miR-429 mimics in HEK293 T cells. The results are shown as mean±SD of three independent experiments. (E) Levels of RBBP4 protein determined by western blot 48 h after the restoration or inhibition of miR-429 in EPCAM− or EPCAM+ HCCLM3 cells. (F) The population of EPCAM+ cells was determined 4 days after the indicated treatment. A total of 3000 cells were seeded into low-adhesion plates and incubated for 10 days to count the number of spheroids. Apoptosis was determined after treatment with 0.5 μg/mL cisplatin for 3 days. Stable overexpression or knockdown of RBBP4 cell lines was generated using a lentivirus system. *p<0.05, ***p<0.001.
To further verify the functional role of RBBP4 in the characteristics of liver T-ICs in vitro, we examined the EPCAM+ cell population, spheroid formation and chemotherapy resistance upon treatment with RBBP4. Figure 4F shows that the knockdown of RBBP4 (see online supplementary figure S7E) increased the percentage of EPCAM+ cells, number of spheroids and chemotherapy resistance. Moreover, the introduction of RBBP4 dramatically reversed miR-429-provoked characteristics of liver T-ICs, suggesting that miR-429 regulates liver T-ICs at least in part by restraining RBBP4 expression. As expected, a higher tumour incidence and greater tumour volume were observed in xenograft mice after they were inoculated with lenti-sh-RBBP4-infected EPCAM− HCCLM3 cells in comparison with lenti-sh-NC-infected cells (see online supplementary figure S7F).
miR-429 decreases RBBP4 expression and results in the activation of POU5F1 (OCT4) transcription by E2F1
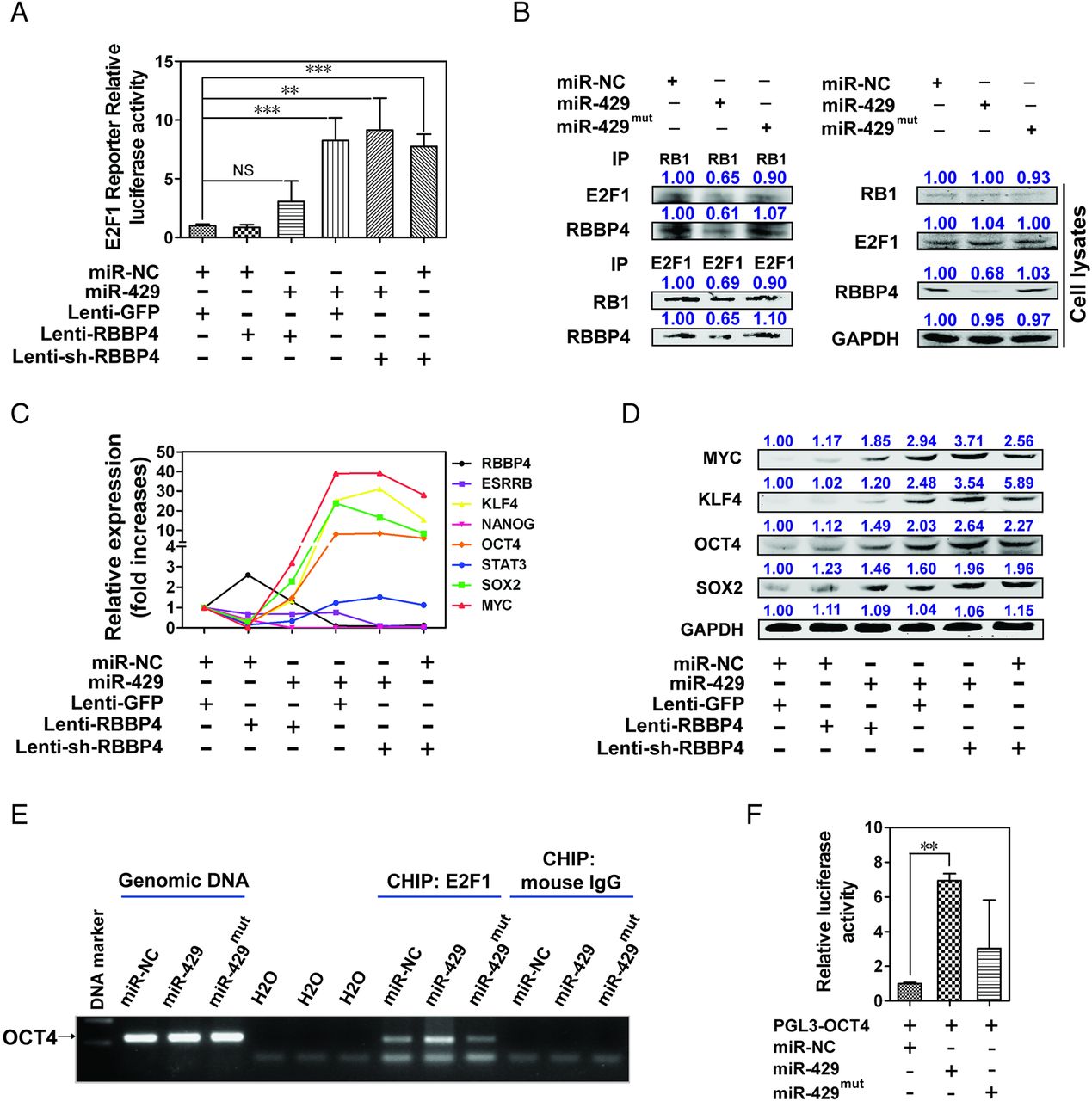
The RBBP4/RB1-associated histone deacetylase complex is involved in the transcriptional repression of E2F1-responsive genes and epigenetic modification.30 ,31 We examined whether miR-429 could affect the transcriptional activity of E2F1 through RBBP4 and found that overexpression of miR-429 or knockdown of RBBP4 alone dramatically enhanced the transcriptional activity of E2F1 (figure 5A). The overexpression of RBBP4 inhibited miR-429-induced E2F1 transcriptional activity, and the exogenous expression of miR-429 did not further enhance the E2F1 activity after RBBP4 knockdown, suggesting that miR-429 regulates the transcriptional activity of E2F1 mainly by directly targeting RBBP4 (figure 5A). Importantly, immunoprecipitation assays showed that the protein–protein interaction between RB1 and E2F1 was significantly weakened upon miR-429 transfection, indicating that miR-429-induced transcriptional activity of E2F1 may result from dissociation of the E2F/RB1 complex caused by miR-429 targeting RBBP4 directly (figure 5B).

miR-429 inhibits the formation of RBBP4/RB1-associated histone deacetylase complex and enhances E2F1 transcriptional activity. (A) Transcriptional activity of E2F1 was determined by a luciferase reporter gene assay in epithelial cell adhesion molecule-negative (EPCAM−) HCCLM3 cells. miR-429 mimics: 100 nM. (B) EPCAM− HCCLM3 cells were treated with 100 nM miR-429 mimics for 4 days. Whole-cell extracts were immunoprecipitated using an anti-RB1 antibody or anti-E2F1 antibody. (C) The mRNA expression of RBBP4 and E2F1- associated genes was determined. 18S was used as an internal control. (D) The protein expression of E2F1 response-genes was estimated 4 days after the indicated treatment. The signal intensity of the target protein was compared with the control group and shown above the band. (E) Chromatin immunoprecipitation from 100 nM miR-429 mimic-treated EPCAM− HCCLM3 cells using E2F1 antibody. Analysis was conducted using specific primers for the promoter region of OCT4. (F) Luciferase assays with reporter constructs containing the promoter region of OCT4 were performed after co-transfection with 100 nM miR-429 mimics in EPCAM− HCCLM3 cells for 2 days. **p<0.01, ***p<0.001, NS, not significant.
E2F1 is a novel regulator of global signalling in pluripotent stem cells and participates in the key transcriptional regulatory network in embryonic stem cells.32 ,33 Figure 5C,D and online supplementary figure S8A show that overexpression of miR-429 or knockdown of RBBP4 markedly upregulated the mRNA and protein expression of the E2F1-responsive genes KLF4, POU5F1 (OCT4), MYC and SOX2. In line with these, the blockage of E2F1 expression with siRNA decreased the expression of these four genes (see online supplementary figures S8B,C). Importantly, chromatin immunoprecipitation (ChIP) showed that more E2F1 was recruited to the promoter regions of the OCT4 gene—a critical transcriptional factor for the maintenance of stem cells and for the chemoresistance of HCC34 ,35—in miR-429-infected cells than in NC mimic-infected cells (figure 5E). Notably, no significant recruitment of E2F1 to the promoter region of KLF4, MYC or SOX2 occurred (data not shown), suggesting that the overexpression of these genes resulted from another indirect mechanism, including but not limited to transcriptional activation of OCT4.33 Figure 5F further confirms that the introduction of miR-429 strongly promoted the effect of E2F1 on OCT4 promoter reporter activity.
To further establish whether miR-429 could regulate EPCAM expression directly through the RBBP4/E2F1/OCT4 axis, we introduced exogenous OCT4 into HCCLM3 cells and examined the level of EPCAM. Interestingly, we found significantly increased expression of EPCAM (see online supplementary figure S8D). Conversely, OCT4 interference further resulted in a kind of downregulation of EPCAM compared with the miR-429-treated group (see online supplementary figure S8E). In addition, EPCAM promoter luciferase activity was upregulated in the presence of OCT4 (see online supplementary figure S8F), suggesting that miR-429-RBBP4 dependent regulation of EPCAM could mediated directly through their downstream factor OCT4. Note that OCT4 interference cannot completely block miR-429-increased EPCAM expression, which suggests that some unknown mechanism such as the network regulation between OCT4, SOX2, KLF4 and MYC may be involved in the regulation of EPCAM (see online supplementary figure S8E).
As expected, we also observed low expression of RBBP4 and high expression of OCT4 in subcutaneous tumour masses in the miR-429 mimic-treated group. Conversely, high expression of RBBP4 and low expression of OCT4 were observed in those of the antagomir-429-treated group (see online supplementary figures S9A,B).
Aberrant CpG methylation enhances the expression of the miR-200b/miR-200a/miR-429 cluster in liver T-ICs
Three miRNAs (miR-429, miR-200b and miR-200a) are clustered on Chr1 and have similar seed sequences (figure 6A). miR-200b, miR-200a and miR-429 are overexpressed in various cancers21 ,36 ,37 and are hypomethylated regulated in pancreatic cancer.37 We thus hypothesised that the increase of miR-429 in HCC resulted from the aberrant CpG methylation status in its putative promoter region.36 Mass array-based quantitative methylation analysis was applied to examine the methylation status of the region upstream of the 5′ miRNA (miR-200b) of the miR-200b/miR-200a/miR-429 cluster38 in HCC cell lines and non-neoplastic cell lines. In total, 12 CpG sequences on Chr 1-6 (10 were detected) and 23 on Chr 1-7 (11 were detected) were analysed (see online supplementary figure S10A and figure 6B). We found that several regions (CpG 8, 9/10, 11 and 12 on Chr 1-6 and CpG 13, 19/20/21, 22 and 23 on Chr 1-7) were significantly hypomethylated in highly miR-429-expressing HCC cell lines (figure 6B), of which four regions (CpG 8, 9/10, 11) on Chr 1-6 showed similar hypomethylation in EPCAM+ HCCLM3 and SMMC-7721 cells, but not in their counterpart EPCAM− cells (figure 6C). Furthermore, the lower methylation status of these four regions was confirmed in very early-stage HCC in comparison with normal liver tissue (figure 6D). To validate the role of DNA methylation in the control of miR-200b/miR-200a/miR-429 cluster expression, we treated HCCLM3 cells (hypermethylated) with 5-Aza-CdR for 2 days and found upregulated expression of miR-429 and miR-200a/miR-200b (figure 6E), indicating a negative correlation between DNA methylation and the expression of these miRNAs. Consistent with this result, miR-200b/miR-200a/miR-429 showed a similar trend of expression in a panel of HCC cell lines and very early-stage HCC tissues (figure 6F and online supplementary figure S10B). To determine whether the miR-200b/miR-200a/miR-429 cluster presents synergetic regulation or competitive inhibition, functional experiments were performed with these miRNAs individually and mixed. To exclude the saturation effects which may cover the additive roles, a final concentration of 20, 60, 100, 140 and 180 nM miR-429 mimics was introduced into HCCLM3 cells, respectively. As shown in online supplementary figure S10C, a gradual increase in the percentage of EPCAM+ HCCLM3 cells was detected in response to miR-429 induction in a dose-dependent manner (from 20 nM to 60 nM and 100 nM), whereas treatment with a higher dose of miR-429 (>100 nM) did not achieve further upregulation of the proportion of EPCAM+ cells, suggesting that such effect may reach a ceiling with an overdose of miRNA mimics. Based on this, final concentrations of 60 and 100 nM miRNA mimics were used in further experiments. Online supplementary figure S10D shows that, like miR-429, miR-200b increased the proportion of EPCAM+ cells and facilitated chemotherapy resistance, spheroid formation and malignant proliferation at both 60 and 100 nM concentrations, but miR-200a had no such significant effect. At the same time, miR-200b/miR-429 mixed group (30 nM miR-200b+30 nM miR-429) produced additive effects compared with 60 nM miR-429 alone, suggesting that miR-200b and miR-429 have a synergetic role rather than a competitive role in carcinogenesis. Given that miR-200b and miR-429 share the same seed sequence, it is possible that these two miRNAs may share some intracellular target genes such as RBBP4 (see online supplementary figure S10E) to regulate synergetically the characteristics of liver T-ICs.

DNA methylation regulates miR-429 expression in liver tumour-initiating cells (T-ICs). (A) Schematic representation of the miR-200b/miR-200a/miR-429 clusters located on Chr1p36. miRNAs sharing the same seed sequence (2–7 base pairs) are marked in the same colour. (B) Coloured bars represent the methylation status of indicated CpG islands detected by quantitative DNA methylation analysis. p Values in red are <0.01. p Values are based on a comparison with the low-miR-429 expression control. (C) Methylation status was analysed in sorted epithelial cell adhesion molecule-positive (EPCAM+) hepatocellular carcinoma (HCC) cells. (D) CpG islands located upstream of the miR-200b/miR-200a/miR-429 cluster were hypomethylated in BCLC stage 0 HCC (n=29) compared with normal liver tissue (n=26). CpG islands located on Chr 1-6 were further investigated. (E) HCCLM3 cells were treated with 10 μM 5-Aza-CdR for 2 days. (F) Arrays were mean-centred, median-centred and normalised using GENE CLUSTER 3.0. Average linkage clustering was performed. ***p<0.001, **p<0.01, *p<0.05.
Secreted miR-429 has the potential to become a proactive signalling molecule mediating intercellular communication
Microvesicles (MVs, including exosomes and microparticles), particularly exosomes, are carriers of miRNAs secreted from cells.39 We examined whether miR-429 can be secreted from HCC cells and packed in MVs. Total RNA and protein in microparticles or exosomes were enriched from the culture supernatant of HCC cell lines (see online supplementary figure S11A) and the presence of exosomes was confirmed by their marker protein CD63 (see online supplementary figure S11B). We then analysed the correlation of the mature miRNA levels in cells with those in microparticles and exosomes. As shown in figure 7A and online supplementary figure S11C, the expression of miR-429 in the intracellular compartment was positively correlated with its level in MVs (especially exosomes). Interestingly, the expression of miR-200a, but not miR-200b, in the intracellular compartment was positively correlated with its level in exosomes, suggesting that distinct mechanisms may be responsible for the extracellular release of miRNAs (see online supplementary figure S11D). Moreover, we found that the expression of miR-429 in the serum of patients with very early-stage HCC was positively correlated with its level in tissues, suggesting that the miR-429 driven from high-expressed HCC tissues may be packed in exosomes and secreted into the serum (see online supplementary figure S11E).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Intracellular and intercellular distribution of miR-429 in cultured hepatocellular carcinoma (HCC) cell lines. (A) Clustering analysis of miR-429 expression in cell lines, microparticles and exosomes. The tree displays their average –△CT values (normalised to the expression of U6). Arrays were mean-centred, median-centred and normalised using GENE CLUSTER 3.0. (B) Quantitative RT-PCR analysis of miR-429 expression in exosomes. The culture medium of HepG2 cells was divided into three equal portions. These samples were treated with RNase, Triton X-100 or RNase plus 0.3% Triton X-100 for 30 min. (C) The percentage of epithelial cell adhesion molecule-positive (EPCAM+) cells was determined 4 days after the indicated treatment. Apoptosis was determined 3 days after treatment with 0.5 μg/mL cisplatin. (D) The proliferation rate was evaluated by a CCK8 assay. Spheroid formation was determined after 3000 cells were seeded in low-adhesion plates for 10 days. (E) White arrows indicate the decreased expression of Cy3-miR-429 in GFP-HCCLM3 cells and white circles indicate the newly Cy3-miR-429-staining HCCLM3 cells that initially lacked Cy3-miR-429. (F) A model showing miR-429 action in manipulating the properties of liver tumour-initiating cells. *p<0.05, **p<0.01, NS, not significant.
We then determined whether miR-429 was indeed present within the MVs, which are surrounded by a double-layer membrane, and are not simply being directly released into the cell culture supernatant, as the miR-429 expression in exosomes treated with RNase was similar to that in untreated media while the miR-429 level in exosomes treated with both RNase and Triton X-100 significantly decreased (figure 7B). A recent study showed that abundantly expressed miR-150 in THP-1 cells could be packaged into MVs and delivered into recipient HMEC-1 cells to exert a proactive role.40 To investigate whether miR-429 could enter into recipient cells via exosomes, the culture supernatant of HepG2 cells, which were transfected with Cy3-labelled miR-429, was collected and incubated with HCCLM3 cells. After incubation for 48 h, we observed red fluorescent dots and an increase of about 200-fold in miR-429 expression in HCCLM3 cells compared with its basal expression (see online supplementary figure S11F). Moreover, the proportion of EPCAM+ cells, chemotherapy resistance ability, cell proliferation and tumour spheroid formation were markedly induced by miR-429-containing MVs (figure 7C,D). In addition, a mixed co-culture experiment also reinforced the conclusion that miR-429 was secreted from high-expressing HCC cells and relocated into the surrounded target cells via MVs. Twenty-four hours after the transfection of Cy3-labelled miR-429, the GFP-HCCLM3 cells were co-cultured with blank HCCLM3 cells in a ratio of 1 : 4. Real-time confocal microscopy showed that Cy3-miR-429 (but not the non-secreted GFP) was released from the GFP-HCCLM3 cells and re-entered the HCCLM3 cells (figure 7E).
Discussion
We report here, for the first time, that the increased expression of miR-429 is an independent and significant factor affecting the survival and tumour recurrence of patients with HCC. miR-429 was preferentially expressed in EPCAM+ cells as well as spheroid-enriched liver T-ICs. Functional experiments showed that miR-429 played important roles in T-IC function. The overexpression of miR-429 endowed EPCAM− cells with increased expression of stem cell-associated genes, greater potential for self-renewal, enhanced chemotherapeutic resistance and greater tumorigenicity. Conversely, the antagonist of miR-429 in EPCAM+ cells resulted in the opposite effect. Interestingly, we also observed the enrichment of miR-429 in CD133+ HCC tissues and sorted HCC cells, whereas the reinforced expression of miR-429 had no obvious effects on the characteristics of CD133+ or CD133− cells, implying that each type of T-IC may have a distinct mechanism for regulating its functions and self-expansion.
Through an integrated analysis of mRNA expression profiles, software predictions and functional studies, we identified RBBP4 as a functional downstream target of miR-429. RBBP4 is a tumour suppressor protein that suppresses the growth of several types of cancer cells in vitro and in vivo. E2F1 and RBBP4 are physically associated in the presence of RB1 and histone deacetylase,30 ,31 suggesting that miR-429 can modulate the transcriptional activity of E2F1 by directly targeting RBBP4 expression. Moreover, the stemness-related OCT4 gene was identified as a novel E2F1-responsive gene that was upregulated upon miR-429 overexpression or RBBP4 knockdown. Expression analysis further documented a negative correlation between miR-429 and RBBP4 in liver T-ICs, implying that RBBP4 may be crucial for liver tumorigenesis. To our knowledge, the potential role of RBBP4 in the regulation of liver T-ICs has not previously been reported. We found that knockdown of RBBP4 promoted the self-renewal and tumorigenicity of EPCAM− T-ICs.
Although several genes have been implicated in T-IC-dependent initiation, progression, metastasis and resistance to therapy, the molecular mechanisms involved in the reciprocal regulation of T-IC functions and their niche or other surrounding normal cells during tumour development are largely unknown. Based on our results, we hypothesise that the secreted miR-429 can enter surrounding cells through MV-mediated intercellular exchange, which provides a permissive niche for cell proliferation, self-renewal, tumorigenicity, chemoresistance and malignant transformation.
The four abnormal hypomethylated sites upstream of the miR-200b/miR-200a/miR-429 cluster in the Chr1p36 region, one of the most unstable chromosome regions documented in various tumours, results in the high expression of these miRNAs in most HCC cell lines and HCC clinical tissues. miR-429 and miR-200b share the same seed sequence and show synergetic regulation on the target gene RBBP4 and the characteristics of liver T-ICs. Because of the deficient function of miR-200a and low intracellular secretion rate of miR-200b, we speculate that miR-429 may be the most effective regulator for liver T-ICs in the miR-200b/miR-200a/miR-429 cluster.
In summary (figure 7F), the miR-200b/miR-200a/miR-429 cluster was hypomethylated in EPCAM+ liver T-ICs and very early-stage HCC samples. miR-429 exerted the most dramatic effects on the properties of liver T-ICs by directly targeting RBBP4 in vitro and in vivo and further promoted the transcriptional activity of E2F1 and the expression of OCT4. Due to the interaction between OCT4 and SOX233 and the potential effect of SOX2 on miR-429 transcription,41 there may be a positive feedback loop to manipulate the properties of liver T-ICs.
Acknowledgments
The authors thank Professor GS Feng for helpful and valuable discussions and Dongping Hu, Dandan Huang, Shanhua Tang, Shanna Huang, Huanlin Sun and Congli Hu for technical assistance.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Data supplement 1 - Online supplement
- Data supplement 2 - Online figures
- Data supplement 3 - Online tables
Footnotes
-
LL, JT and BZ contributed equally.
-
Contributors LL, JT and BZ contributed to study concept, design and acquisition of data. LC and WY contributed to study design and acquisition of data. RW, YT, CC, JF, YC, MLG, JF, FJ, YY, JG, DW, GC, LG and DC contributed to acquisition of data. HL and MW contributed to analysis and interpretation of data. MQZ contributed to statistical analysis. HW contributed to study concept and design, drafting of the manuscript and critical revision of the manuscript.
-
Funding This study was supported by the following grants: The State Key Project for Liver Cancer (2012ZX10002-009), the National Basic Research Program of China (2012CB316503), National Natural Science Foundation of China (81372674, 81300306, 81221061, 91029723 and 81272212) and the Science Foundation of Shanghai (XYQ2013076 and 11ZR1449400).
-
Competing interests None.
-
Ethics approval Ethics approval was obtained from the Ethics Committee of the Eastern Hepatobiliary Surgery Hospital, Shanghai, China.
-
Provenance and peer review Not commissioned; externally peer reviewed.