Article Text

Abstract
Objective Serum lipase activities above the threefold upper reference limit indicate acute pancreatitis. We investigated whether high lipase activity—within the reference range and in the absence of pancreatitis—are associated with genetic single nucleotide polymorphisms (SNP), and whether these identified SNPs are also associated with clinical pancreatitis.
Methods Genome-wide association studies (GWAS) on phenotypes ‘serum lipase activity’ and ‘high serum lipase activity’ were conducted including 3966 German volunteers from the population-based Study-of-Health-in-Pomerania (SHIP). Lead SNPs associated on a genome-wide significance level were replicated in two cohorts, 1444 blood donors and 1042 pancreatitis patients.
Results Initial discovery GWAS detected SNPs within or near genes encoding the ABO blood group specifying transferases A/B (ABO), Fucosyltransferase-2 (FUT2), and Chymotrypsinogen-B2 (CTRB2), to be significantly associated with lipase activity levels in asymptomatic subjects. Replication analyses in blood donors confirmed the association of FUT-2 non-secretor status (OR=1.49; p=0.012) and ABO blood-type-B (OR=2.48; p=7.29×10−8) with high lipase activity levels. In pancreatitis patients, significant associations were found for FUT-2 non-secretor status (OR=1.53; p=8.56×10−4) and ABO-B (OR=1.69, p=1.0×10−4) with chronic pancreatitis, but not with acute pancreatitis. Conversely, carriers of blood group O were less frequently affected by chronic pancreatitis (OR=0.62; p=1.22×10−05) and less likely to have high lipase activity levels (OR=0.59; p=8.14×10−05).
Conclusions These are the first results indicating that ABO blood type-B as well as FUT2 non-secretor status are common population-wide risk factors for developing chronic pancreatitis. They also imply that, even within the reference range, elevated lipase activities may indicate subclinical pancreatic injury in asymptomatic subjects.
- Chronic Pancreatitis
- Genetic Polymorphisms
- Linkage Analysis
- Glycosyltransferases
- Pancreatic Enzymes
Statistics from Altmetric.com
Significance of this study
What is already known on this subject?
-
The definition of acute pancreatitis includes characteristic clinical symptoms in combination with elevated serum enzyme activities (amylase or lipase) above three times the upper reference limit.1 Elevated lipase levels below this cut-off are not considered diagnostic for pancreatitis, and can be observed in a number of non-pancreatic conditions.
-
No serum biomarker presently exists for chronic pancreatitis
What are the new findings?
-
In asymptomatic volunteers from a population-based study (N=3966) and healthy blood donors (N=1444), genetic polymorphisms that determine the so-called Fucosyltransferase-2 non-secretor status and the ABO blood-type B, were significantly associated with increased serum lipase activities.
-
This study is the first to identify Lewis and AB0 blood group systems as disease- relevant for the development of pancreatitis. FUT2 non-secretors and blood group B carriers displayed 1.53-fold and 1.69-fold odds of developing chronic pancreatitis, whereas in blood type 0 carriers, the odds were clearly reduced (0.62-fold).
-
FUT2 non-secretor status and blood group represent genetic cofactors in the development of alcoholic as well as idiopathic chronic pancreatitis, but apparently do not influence the risk for acute pancreatitis.
How might it impact on clinical practice in the foreseeable future?
-
Blood group B (12–15% in a Western population) and FUT2 non-secretor status (20%) are common genetic traits associated with serum lipase activities and the risk of developing chronic pancreatitis.
-
Future studies investigating genetic or environmental risk factors for pancreatitis will have to correct for these factors in their cohorts because they vary greatly between populations (eg, blood group B among Kashmiris 32%, Norwegians 8%).
-
Elevated serum lipase activities in asymptomatic subjects may indicate, even when within the reference range, subclinical pancreatic injury.
Introduction
Human pancreatic lipase (PNLIP, OMIM 246600) is secreted by the exocrine pancreas through the pancreatic duct system and into the duodenum. Here, it has a major role in breaking down dietary lipids such as triglycerides, oils and fats. In healthy individuals, only little lipase activity is found in the blood, the source of which is either basolateral exocytosis of pancreatic secretory enzymes2, or physiological turnover of acinar cells.3
Almost a century ago it was recognised that high serum levels of pancreatic secretory enzymes (initially amylase and later lipase) indicate acute injury of the pancreas.4 Today, all current guidelines require for the diagnosis of acute pancreatitis (AP), in addition to characteristic abdominal symptoms, an elevation of amylase or lipase of three times the upper limit of normal or higher.1 ,5 Conversely, for the diagnosis of chronic pancreatitis (CP) no such serum marker exists, and neither elevated nor reduced lipase activity levels are of sufficient diagnostic accuracy to be of clinical use.6 ,7 High subthreshold serum lipase activity is common in patients with critical illness of any kind, cholecystitis, gastric ulcer disease, bowel obstruction, viral gastroenteritis and renal failure, so that an underlying pancreatic disorder could so far not safely be assumed in symptomatic patients.8
In a previous population-based study, subthreshold elevated serum lipase activities were found to be associated with renal impairment, advanced age, or the use of various medications that modulate the immune system,9 all of which can occasionally be associated with subclinical pancreatic damage, or overt pancreatic disease.6 However, lipase elevations in this range are not generally regarded as diagnostic for pancreatic disease.10 The question of whether or not a genetic predisposition that involves increased lipase serum activities in asymptomatic subjects represents a risk factor for either pancreatitis or pancreatic cancer has remained unresolved.11 To test this hypothesis we have performed genome-wide association studies (GWAS) on the phenotypes ‘serum lipase activity’ and ‘high serum lipase activity’ analysing asymptomatic volunteers of the population-based Study-of-Health-in-Pomerania (SHIP). The results indicate that ABO-blood group B and FUT2 non-secretor status are associated with asymptomatic hyperlipasemia and also increase the risk of developing CP.
Material and methods
SHIP study population and sample selection
SHIP is a population-based study conducted in West Pomerania, the northeastern part of Germany, and its details have been previously reported.12 From 212 000 inhabitants of this area, a random sample was drawn via the residence registration office where all residents are registered. The SHIP-0 cohort included 4308 volunteer participants (2008 males, aged 20–81 years) with German citizenship, European ancestry and principal residence in the study area. Participants with a history of pancreatitis or renal insufficiency, due to estimated glomerular filtration rate (eGFR CKD-EPI) <30 mL/min per 1.73 m2)13 were excluded from further analysis. For our study population of 3966 participants, serum lipase activity was determined, and genome-wide SNP-typing was performed. Approval was obtained by the local ethic committee, and informed consent was given by all participants.
Blood donors and pancreatitis patients
The replication cohort consisted of 1500 blood donors (18–68 years, mean age 32.4 years, 58.2% male) who were recruited for the study in 2010 at University Medicine Greifswald. Written consent and information on alcohol and tobacco consumption, as well as a history of pancreatic disease was recorded. Subjects with a self-reported history of pancreatitis (N=9) were excluded from further analysis. To test the disease relevance of the identified SNPs we performed a second replication in 1042 German pancreatitis patients (5–97 years, mean age 46.6 years, 62.3% male) that had been enrolled in an observational study (ProZyt) between 2003 and 2011. This cohort included patients with AP (n=505) and CP ( n=537).14––16 All individuals gave their written informed consent for participation and genotyping, and the local ethics committee approved the study protocol. Definition for AP was abdominal pain in combination with a greater than three-fold elevation of serum amylase or lipase activity. Diagnosis of CP included the presence of unequivocal morphological evidence on computed tomography (CT) and/or endoscopic retrograde cholangio pancrreatography (ERCP) and was labelled idiopathic (ICP (idiopathic chronic pancreatitis); n=229) in subjects in whom no known risk factors, including gallstones, hyperlipidaemia or hypercalcaemia could be identified. Alcoholic aetiology (ACP (alcoholic chronic pancreatitis); n=214) was assumed in subjects with a history of regular alcohol consumption (more than two drinks or 20 g/day).
Phenotype determination
Serum lipase activities in the SHIP-cohort were measured using a turbidometric assay on a Hitachi 717 analyser (Roche Diagnostics GmbH, Mannheim, Germany). The ‘serum lipase activity’ phenotype in the continuous association analysis refers to the square root of serum lipase activity, and the ‘high serum lipase’ phenotype in the dichotomous analysis was defined as binary variable comparing samples with the highest 25% of serum lipase activity versus the remaining 75%. Serum lipase activity could be analysed in 1444 blood donors using the ADVIA 1800 Siemens Lipase (LIP), colorimetric test (Siemens, Germany) according to the manufacturer's protocol, and were in the range between 0.21–3.05 μmol/sl (Median 0.45). Again, the ‘high serum lipase activity’ phenotype was defined as binary variable comparing samples with the highest 25% of serum lipase activities (>0.55 µmol/sl) versus the remaining 75%.
Genome-wide SNP genotyping
SHIP samples were genotyped using the Affymetrix Human SNP Array 6.0 as previously described.12 ,17 Imputation was carried out using the HapMap v22 reference panel. For our analysis, we excluded all SNPs with a Hardy–Weinberg equilibrium p value <0.001, a minor allele frequency (MAF) <0.01 and an imputation quality score <0.4.
Genotyping of blood donor and pancreatitis cohorts
Genotyping was performed using specific TaqMan assays (Applied Biosystems) for rs632111, rs8176693, rs889512, rs17303392 and rs601338. In some individuals, also the ABO blood group was analysed or verified by genotyping analysis of rs8176719, rs8176747, rs8176750. The genotypes were analysed using TaqMan 7500 RT-System (Applied Biosystems). Samples were, in part, verified and analysed by PCR-amplification with specific oligonucleotide primers followed by Sanger sequencing using the BigDye V.3.1 terminator sequencing kit and a 3130xl genetic analyser (Applied Biosystems). All SNPs were in Hardy–Weinberg equilibrium (p value >0.001) and had a MAF >0.01.
Statistical analysis
Because lipase activity values (and corresponding residuals after adjusting for covariates age, sex, smoking and alcohol) were right-skewed, we performed a square-root transformation of the lipase measurements to normalise the distribution and to reduce false positive results. To maximise the statistical power, serum lipase activity levels were analysed as a continuous outcome using linear regression. As a complement, and to consider possible non-linear effects of the allele counts on serum lipase activity levels, we also performed logistic regression defining the outcome according to the phenotype ‘high serum lipase activity’. Linear regression is normally used to establish a relationship between dependent and independent variables, whereas logistic regression can ascertain the probability of an event (‘high lipase’).
For a total 3966 SHIP-0 samples and 1444 blood donors, SNP associations were tested using linear regression on the square root transformed serum lipase levels in µmol/s/l for the ‘serum lipase activity’ phenotype, and logistic regression for the ‘high serum lipase activity’ phenotype assuming additive genetic effects. Adjustment for age, gender, smoking and alcohol was performed. Furthermore, adjustment was performed as follows: in SHIP-0 for the smoking status (never-smoker, ex-smoker and current smoker) and the amount of alcohol usage in g day−1, in the replication cohort for the lifetime tobacco exposure in pack years, and the Alcohol Use Disorders Identification Test—C (AUDIT-C) score as a measurement for the alcohol usage. For the discovery-stage GWAS in SHIP-0, associations with a p value below 5×10−8 were considered genome-wide significant.18 The estimated genomic control was low for ‘serum lipase activity’ and ‘high serum lipase activity’ (λGC=1.0185 and λGC=0.9952, respectively), suggesting little residual confounding due to population stratification. Genomic control was applied for the ‘serum lipase activity’ analysis only. To identify possible independent associations per locus, lead SNPs were selected by the PLINK clumping algorithm (R2 > 0.01, 1 Mb distance19;) based on genotype data of the HapMapII r28 CEU samples, but no additional hits were found. Associations were considered replicated in the blood donor cohort with a p value below 0.05/4=0.0125 (Bonferroni-correction for 4 tests); χ2 tests were performed with help of the interactive calculation tool for χ2 tests of goodness-of-fit and independence, available at (http://quantpsy.org/chisq/chisq.htm).
Results
Discovery-stage GWAS
In the discovery-stage GWAS, 3966 individuals were analysed for associations of SNPs with the phenotypes ‘serum lipase activity’ and ‘high serum lipase activity’. The GWAS identified three and two loci that exhibited genome-wide significant associations for ‘serum lipase activity’ and ‘high serum lipase activity’, respectively (figure 1A,B, see online supplementary figure 1A,B and table 1). The ABO locus at chromosome 9q34 exhibited the strongest association with ‘serum lipase activity’ (p=1.29×10−30) as well as with ‘high serum lipase activity’ (p=1.28×10−22) where the minor allele of the common lead SNP of both analyses, rs8176693_T, was associated with increased lipase activity (figure 2A, B). The second locus is located in the FUT2 region at 19q13 (p=6.15×10−9 and 5.72×10−9 for ‘serum lipase activity’ and ‘high serum lipase activity’, respectively; figure 2C, D). As for the first locus, the minor allele of the common lead SNP, rs632111_G, was associated with increased lipase activities. The lead SNP rs889512 of the third locus CTRB2 at 16q23 was only genome-wide significantly associated with the ‘serum lipase activity’ phenotype (p=1.91×10−8; figure 2E, F). In this case, the minor allele rs889512_G was associated with decreased lipase activity.
Lead SNP associations from the discovery-stage Study-of-Health-in-Pomerania-Genome-wide association studies (GWAS) on the phenotypes ‘Serum lipase activity’ and ‘High serum lipase activity’
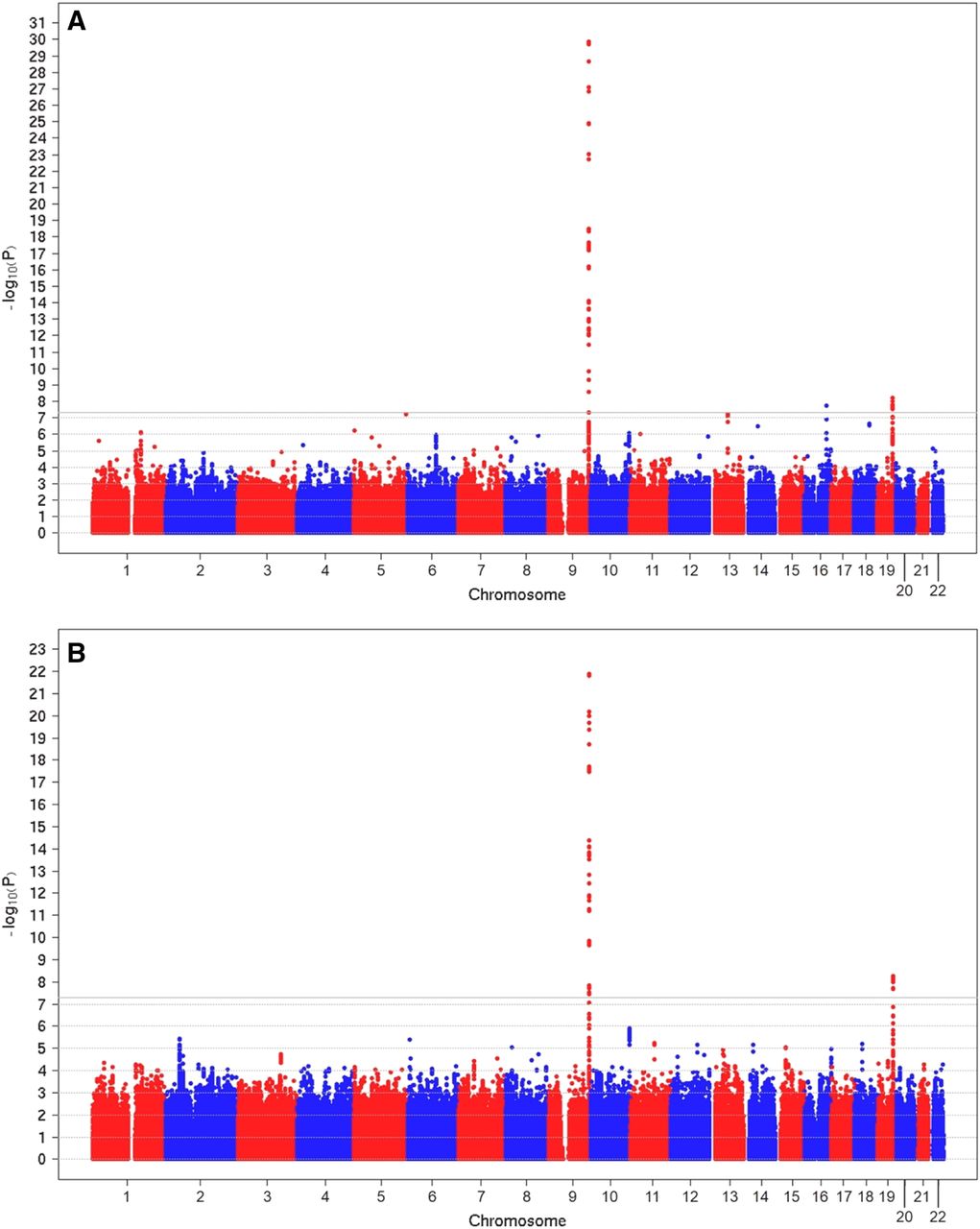
Manhattan plot showing significance of association for all SNPs with a minor allele frequency >1% in the genome-wide association analysis. The ‘serum lipase activity’ phenotype (A) refers to the square root of serum lipase concentrations and the ‘high serum lipase’ phenotype (B) was defined as binary variable comparing samples with the highest 25% of serum lipase concentrations versus the remaining 75%. SNPs are plotted on the x-axis according to their chromosomal position against the association with the phenotype (shown as -log10 p value) on the y-axis. The grey horizontal line indicates the threshold for genome-wide significance (p=5×10−8).

Regional plots of the three loci associated with ‘serum lipase activity’ and ‘high serum lipase activity’. Shown are the ABO region (A,B), the FUT2 region (C,D) and the CTRB2 region (E,F), displaying the results for ‘serum lipase activity’ on the left and for ‘high serum lipase activity’ on the right, respectively. The region's lead SNP from the ‘serum lipase activity’ Genome-wide association studies is coloured in purple while other SNPs in the region are given as colour-coded smaller circles. Red indicates high correlation with the top SNP (R2>0.8); orange indicates moderate correlation (0.6<R2≤0.8); green colour indicates weak correlation (0.4<R2≤0.6); while light and dark blue indicate no correlation with the top SNP (R2≤0.4). The y-axis on the right indicates the estimated recombination rates (based on HapMap Phase II) given in light blue, and the bottom of each panel shows the intron-exon structure of the respective annotated genes at the locus and their transcriptional direction.
Replication analysis
Serum lipase activity was determined in 1444 local blood donors who were also individually genotyped for the lead SNPs of the discovery-stage GWAS. Association analyses revealed that again the ABO locus exhibited the strongest association with the ‘serum lipase activity’ phenotype (p=2.92×10−19) as well as with the ‘high serum lipase activity’ phenotype (p=5.19×10−9; table 2). Significant association of the FUT2 and CTRB2 loci with both phenotypes was also demonstrated (p=2.05×10−6 and 9.94×10−6 for ‘serum lipase activity’ and p=1.98×10−4 and 3.31×10−4 for ‘high serum lipase activity’). In all cases, the observed effect directions were the same as in the discovery GWAS. These results confirmed the significant association of polymorphisms within the ABO, FUT2 and CTRB2 loci with serum lipase activity in asymptomatic subjects.
Replication of the discovery Genome-wide association studies (GWAS) lead SNPs in a validation cohort of 1444 local blood donors
Analysis of the correlation between ABO locus association and ABO blood group
The GWAS lead SNP rs8176693 is located in the first intron of ABO, which raised the question whether it is associated with a distinct ABO blood group. As blood groups were not determined in SHIP, we analysed the correlation between rs8176693 and ABO blood group subtypes in our blood donor replication cohort (n=1491) where blood groups were typed serologically and verified, in specific cases, using TaqMan or sequencing analysis of ABO (see online supplementary table S1). We found complete co-occurrence of rs8176693_T and blood group B, with a single exception in one individual, in which rs8176693_T was associated with blood group A1/0. Obviously the minor allele of rs8176693 belongs to the ABO haplotype that determines blood group B. A high linkage disequilibrium (LD) is indicated by a 2.5-fold over-representation of individuals with blood group B (p=7.3×10−08) in the blood-donor subgroup exhibiting the 25% highest lipase activities (figure 3). By contrast, blood group 0 was significantly under-represented (OR=0.59; p=8.1×10−05) in this subgroup, and carriers of blood group A were equally frequent in both groups. After stratification for blood groups, we could confirm that in the blood donor cohort, the mean lipase activity was highest in carriers of the blood groups B and AB. These results indicate significant associations of blood groups B and 0 with high and low serum lipase activities, respectively.

Association of blood group and serum lipase activity in the blood donor replication cohort. ABO blood groups were typed in 1444 donors by serological tests. Blood group prevalences and mean lipase activities (µmol/sl) were calculated for the blood donor cohort as well as for subcohorts of high lipase (25%) and low lipase (75%) phenotypes (A). A forest plot illustrates the different ORs (and 95% CI) of specific blood groups in the 25% of blood donors with highest lipase activities. Results of Pearson's chi-square tests are also shown (B).
Analysis of the correlation between FUT2 locus association and secretor status
Fucosyltransferase 2 (FUT2) determines an individual's aibility to secret ABO blood type antigens into body fluids. This so-called secretor (Se) or a non-secretor status (se) is completely independent of the ABO blood type. Twenty percent of individuals with European ancestry are ‘non-secretors’ because they represent homozygous carriers of the FUT2 W134X allele (SNP rs601338_A) resulting in a FUT2 null phenotype.20 The observation that the GWAS lead SNP rs632111 is located in the 3′-untranslated region (3′-UTR) of FUT2 raised the question whether it is also associated with the non-secretor (W134X) allele. Therefore, rs601338 was additionally genotyped in the 1444 individuals of our blood donor replication cohort (table 3, see online supplementary table S3). The corresponding R2-value was 0.88, indeed demonstrating a shared underlying haplotype for rs601338_A and rs632111_G. Similar results (R2=0.87) were obtained for the pancreatitis patients cohort subsequently analysed (N=973, see below). In accordance with these results, we identified a significant overpresentation of the non-secretor rs601338_A allele (OR=1.42; p=1.88×10−4) in the blood-donor subgroup exhibiting 25% highest lipase activities (table 3). Homozygous carriers of the rs601338_A allele (non-secretors) were significantly over-represented (OR=1.49; p=0.012) in this group.
Association of rs601338 (W134X SNP) and secretor status with lipase activity in blood donor cohort
Replication of associations in pancreatitis patients
In order to test whether the loci identified as associated with increased serum lipase activities are also associated with an increased risk for pancreatitis, we typed the FUT2 locus lead SNP rs632111, the linked W134X SNP rs601338, and the CTRB2 locus lead SNP rs889512 in a further cohort consisting of patients with acute and CP. Genotype distributions for all SNPs were in Hardy–Weinberg equilibrium (see online supplementary table S3). In addition, ABO blood groups, if not available from medical records, were also typed (see online supplementary table S2). The obtained results were compared with those determined in the blood donor replication cohort. These analyses revealed an over-representation of blood group B prevalence in patients with CP (OR=1.69, p=0.0001) but not in those with AP (figure 4). This association was found in CP of either idiopathic (ICP) or alcoholic (ACP) aetiology (OR=1.57, p=0.0189 and OR=1.65, p=0.0097). Further nominal significant associations were found between A and AB blood groups and ICP and AP, respectively (OR=1.36; p=0.032 and OR=1.71; p=0.008). Carriers of blood group O tended to be less frequent among pancreatitis patients, and their proportion was significantly decreased in patients with ICP (OR=0.51, p=2.51×10−05).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Association results of blood groups in pancreatitis cohort versus healthy blood donors. ABO blood groups were typed by TaqMan analysis or serologic testing in 1042 pancreatitis patient and compared with 1444 blood donors. Pie charts (A) show the blood group distribution in BD, blood donor cohort; Pan, total pancreatitis cohort; CP, chronic pancreatitis; AP, acute pancreatitis; ICP, idiopathic chronic pancreatitis; ACP, alcoholic chronic pancreatitis. Pearson's χ2 tests were calculated of blood type frequencies in patient cohorts vs. blood donors (B). PVAL, p value (χ2 test), 95% CI, 95% CI of OR, grey shaded cells are nominal significant (PVAL<0.05) and red shaded cells are significant after Bonferroni correction for 15 tests (PVAL<0.05/15 = 0.003).
We further identified a significant allele imbalance for the two FUT2 SNPs typed in the pancreatitis cohort (table 4). Homozygosity for the minor alleles (table 4, Recessive Model) of rs632111 (GG) as well as of rs601338 (AA, non-secretors) was significantly associated with CP (OR=1.58, p=2.4×10−4 and OR=1.57, p=0.001) but not with AP. Interestingly, FUT2 non-secretors are over-represented in idiopathic (OR1.7, p=0.003) but not in alcoholic CP. Taken together, these results suggest that the FUT2 and ABO glycosyltransferases, both cooperating in the synthesis of Lewis antigens Leb and LeY, contribute to the risk of developing pancreatitis. The CTRB2 locus lead SNPs rs889512 was not significantly associated with pancreatitis of any aetiology in the analysed patient cohort.
Association results of Genome-wide association studies (GWAS) top SNPs in the pancreatitis cohort vs healthy blood donors
Co-occurrence of blood group B and non-secretor phenotype
While the non-secretor phenotype (se/se) was not significantly different distributed between the individual ABO blood groups in the blood donor replication cohort, this was found to be the case in the pancreatitis cohort, where carriers of blood groups AB and O less frequently and more frequently, respectively, represented non-secretors. Subsequently, we analysed whether blood group B and non-secretor status (se/se) represent independent or combined/synergistic risk factors for CP. As shown in table 5 no change in combined risk was detected in the pancreatitis cohort, neither in ICP nor in ACP. However, it has to be considered that the corresponding double genotype/phenotype (se/se-blood plus group B) is quite rare, as B represents a low-frequency blood group in the general population. Therefore, the statistical power of this analysis is rather limited.
Association results (A) and comparison (B) of extreme groups in the pancreatitis cohort vs healthy blood donors
Discussion
Relationships between blood groups and disease phenotypes have been debated for almost a century, when variations in blood group frequencies between different populations were identified. This study is the first to identify components of the Lewis and ABO blood group systems as genetic risk factors for the development of CP. Our findings suggest that non-secretor status and blood group B both predispose to this chronic disease in the absence of other known precipitating risk factors. While blood group B was associated with an increased disease risk, group O was significantly under-represented and appears to have a protective effect. In the initial GWAS and the replication study, we identified FUT2 non-secretor status and the blood group B as being significantly associated with elevated serum lipase activity in asymptomatic volunteers. The fact that pancreatic injury is associated with increased serum lipase activity has long been established for AP. In CP, on the other hand, increased activities due to acute inflammatory episodes, as well as decreased lipase levels due to reduced exocrine tissue have been observed, and serum lipase measurements were, therefore, considered to be of little diagnostic value. Elevated lipase activity in asymptomatic subjects has been correlated in previous studies with increased serum creatinine levels, advanced age, use of steroids or antihypertensive medication as well as self-reported CP, but was not considered to be a risk factor for CP in itself. The current data raise the possibility that hyperlipasemia—even below the 3× upper reference limit and, therefore, within the range defined as normal—may be a marker of subclinical and continuous pancreatic injury in asymptomatic subjects. The common denominator for the observed clear associations of the ABO and FUT2 loci with high lipase activity as well as with pancreatitis might be that, on a population-based level rather than an individual level, high lipase activity indicates subclinical pancreatic damage. An alternative explanation is that a common underlying mechanism causes pancreatitis and elevated serum lipase activity, and that this mechanism involves specific glycosylation patterns mediated by the haplotypes underlying blood group B and the non-secretor status. Under the latter scenario, serum lipase activity would simply represent a surrogate marker for other, not yet identified disease-relevant proteins involved in pancreatitis pathogenesis.
The third locus that was significantly associated with high lipase activity in the population-based GWAS, CTRB2, was not associated with pancreatitis in the patient cohort. There are, again, different explanations for this finding, the most simple of which is that the association of this locus with pancreatitis simply is much weaker than the associations between pancreatitis and the ABO and FUT2 loci. In this case, a larger patient cohort would be necessary to reach a comparable level of statistical significance for CTRB2.
A, B and H antigens, the antigenic determinants of the ABO blood group system, are formed by the sequential addition of specific monosaccharides to the carbohydrate side chains of glycolipids and glycoproteins. These specific glycans decorate the surface of erythrocytes. The biosynthesis of A and B antigens involves glycosyltransferases that catalyse the addition of N-acetyl-D-galactosamine or D-galactose to core H antigens. A single base pair deletion resulting in the inactivation of these glycosyltransferases is the reason why in individuals with blood group O the H antigen remains unmodified. The common denominator of the H antigen, the Fucα1.2-glycan structure, is formed by specific α (l.2) fucosyltransferases which play a regulatory role in the tissue expression of the ABH antigens. In the bone marrow (from where erythrocytes originate) fucosylation of ABH antigens is performed by the common H-(fucosyl)-transferase (FUT1). However, ABH antigens are not confined to erythrocytes but are expressed on other tissue cells, body fluids or along the gastrointestinal mucosal lining in individuals of ‘positive secretor status’ (secretor phenotype, Se).21 This is because Se individuals carry at least one functional allele (Se/Se or Se/se) for the Secretor-(fucosyl)-transferase (FUT2), which produces the Fucα1.2-glycan structure in secreted body fluids, such as saliva, tears, breast milk, gastric juice and gastrointestinal mucus secretions. Individuals who are ‘secretors’ express blood group O antigens (Lewis b (Leb) and H antigens), which can be further converted by the ABO glycosyltransferases into A or B antigens, respectively.22 By contrast, individuals of non-secretor phenotype, Se0, lack the secretor-transferase altogether, and synthesise the shorter Lewis a antigen (Lea).20 Approximately 20% of the world population are non-secretors, but geographical and ethnical differences do exist.20
The functional significance of ABO antigen expression outside the blood has not been defined, but several studies have found correlations between disease susceptibilities and ABO blood group or secretor status. One long-established association is that between blood group O and peptic ulcer disease.23 Reilly et al recently reported an association between ABO polymorphisms and myocardial infarction in patients with coronary artery disease. The ABO association was attributable to the glycotransferase deficiency specified by the ABO blood group O genotype previously proposed to protect against myocardial infarction.24 Of particular importance in the context of our study are the recently reported associations between ABO blood group and the risk and outcome of pancreatic cancer.25––27 Cohort studies in independent populations confirmed a significant higher risk of developing sporadic pancreatic cancer in carriers of the blood group B. Carriers of blood groups A and AB exhibited an intermediate risk, and carriers of blood group O demonstrated the lowest risk.28––30 Compared with the OO genotype, addition of each non-O allele increased the pancreatic cancer risk. Subjects with AO and AA genotype had ORs of 1.33 and 1.61, while individuals with BO and BB genotypes had ORs of 1.45 and 2.42.29 Strikingly, according to the HapMap (release 22) database, the ABO lead SNP rs8176693 identified in our study to be associated with ‘serum lipase activity’ as well as with ‘high serum lipase activity’ is in perfect LD (D′ value: 1.0) with rs505922, the lead SNP of the initial GWAS on pancreatic cancer.24 The minor alleles of both SNPs were associated with increased serum lipase activity and increased risk for pancreatic cancer, respectively, indicating a shared haplotype that apparently also carries the polymorphisms specifying blood group B. Indeed, the D′ values between blood group B defining SNPs rs8176743, rs8176746 and rs8176747 on the one hand, and rs8176693 on the other hand also consistently amounted to 1.0. The corresponding D′ values between these three ABO-B SNPs and rs505922 were 1.0, 1.0, and 0.56. Interestingly, while the r2 values between the three ABO-B SNPs and rs8176693 were also consistently high (1.0, 1.0, 0.69), this was not the case between the ABO-B SNPs and rs505922 (0.16, 0.16, 0.073) as well as between rs505922 and rs8176693 (0.16). This reflects the higher MAF (according to HapMap data) of the pancreatic cancer risk lead SNP rs505922 (0.363) as compared to the ABO-B SNPs (0.08, 0.075, 0.123) and the lead SNP rs8176693 (0.083) in individuals with European ancestry. We therefore hypothesise that rs505922 represents a phylogenetic older SNP which has already been present on an ancestral haplotype and is only associated with increased pancreatic cancer risk via its LD with the more recently arisen haplotype that is associated with blood group B and increased serum lipase activity. According to this model, the increased pancreatic cancer risk would be caused by the presence of the latter haplotype detected in our study. Inactivation of the ABO-encoded blood group specifying glycosyltransferases mediates a protective effect against pancreatic cancer and, according to the results of our study, mediates a decreased risk for developing CP. Today, we have only a limited understanding of the initiating molecular events in pancreatic cancer and the potential role of ABH blood groups in pancreatic tumorigenesis. It is, however, well established that persisting CP increases the risk of developing pancreatic cancer.31 ,32 One possible mechanism explaining our findings is a 25% increased level of von-Willebrandt-Factor (VWF) in individuals who have a blood group other than O.33 The glycosylation of VWF by ABO A and B glycosyltransferases affects circulating VWF levels and function through several mechanisms.34 These may include the rate of VWF synthesis and secretion, its proteolysis by metalloprotease ADAMTS13, and the modulation of VWF clearance.35 VWF is a negative modulator of angiogenesis, and displays proinflammatory activity which could promote a chronic inflammatory state.
A recent GWAS demonstrated that the FUT2 non-secretor status is a risk factor for Crohn's disease,36 but apparently is no risk modifier in the association of ABO antigens with pancreatic cancer.37 Being a non-secretor has several potential health disadvantages. Secreting blood-type antigen into saliva, mucus, and other body fluids may add a degree of protection against environmental factors, particularly with respect to micro-organisms. Secretors have a better capacity to stabilise the blood-type-dependent intestinal bacterial ecosystem. A part of the intestinal microbiome that is considered beneficial (probiotics) preferentially feed on the glycosylation products that define the blood type system. Bacteria in the digestive tract of secretors may benefit from a constant nutritional supply by the secretion of blood group antigens in the mucus that lines the intestinal mucosa. The composition of this commensal flora is a contributing mechanism of the ABO and Lewis blood group-dependent risk for inflammatory bowel disease,36 various gastrointestinal infections38––40 and could influence the development of chronic inflammation in the pancreas.
The identification of blood group B and non-secretor status as inherited risk factors for pancreatitis advance our understanding of the complex interactions between environmental and genetic risk factors in the development of the disease. By contrast with dominant risk factors like the catonic trypsinogen (PRSS1) mutations R122H and N29I, several sequence variations in SPINK1 (PSTI), Chymotrypsinogen C (CTRC) or CFTR are now recognised as predisposing factors, which are probably not disease-causing by themselves. It is assumed that ‘weak’ genetic risk factors and other precipitating factors are adding up to a complex disease risk for CP in affected individuals. Blood group and non-secretor status appear to belong to this category. However, the question remains why no ABO-association was found in previous studies,41 and why the prevalence of CP varies so widely (from 1.6 to 23/100.000) across populations.42––44 One reason for this variation may involve the genetic background of different ethnicities which includes, among other traits, a vastly different distribution of ABO blood types ranging from, for example, 32% for blood type B among Kashmiris and 8% in Norwegians. Future investigations into the environmental and genetic risk factors of pancreatitis should, therefore, regard the genetic background of their cohorts as a confounding factor that needs to be corrected for, and also the ABO blood type and FUT-2 secretors status.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Data supplement 1 - Online figures
- Data supplement 2 - Online tables
Footnotes
-
FUW and CS are equally contributing first authors. GH and MML are equally contributing senior authors.
-
Contributors Planning and concept of study: FUW, HV, AG, UV, MML. Acquisition of data: FUW, CS, AG, FE, PS. Statistical analysis: FUW, CS, AT. Data interpretation and manuscript revision: FUW, CS, AG, AT, JM, PS, HV, AG, MZ, J-PK, UV, GH, MML. Writing committee: FUW, CS, GH and MML.
-
Funding Funding for this project was received from Alfried-Krupp-von-Bohlen-und-Hahlbach-Foundation (Graduate Schools Tumor Biology and Free Radical Biology), the Deutsche Krebshilfe/Dr. Mildred-Scheel-Stiftung (109102), the Deutsche Forschungsgemeinschaft (DFG GRK840-E3/E4, MA 4115/1-2/3), the Federal Ministry of Education and Research (BMBF GANI-MED 03152061A, BMBF 0314107), the European Union (EU-FP-7: EPC-TM and EU-FP-7-REGPOT-2010-1) and EFRE-State Ministry of Economics MV (V-630-S-150-2012/132/133). SHIP is part of the Community Medicine Research net of the University Medicine Greifswald, Germany, which is funded by the Federal Ministry of Education and Research (grants no. 01ZZ9603, 01ZZ0103, and 01ZZ0403), the Ministry of Cultural Affairs and the Social Ministry of the Federal State of Mecklenburg-West Pomerania. Genome-wide data have been supported by the Federal Ministry of Education and Research (grant no. 03ZIK012) and a joint grant from Siemens Healthcare, Erlangen, Germany and the Federal State of Mecklenburg-West Pomerania. The University of Greifswald is a member of the ‘Center of Knowledge Interchange’ programme of the Siemens AG and the Caché Campus programme of the InterSystems GmbH.
-
Competing interests None.
-
Patient consent Obtained.
-
Ethics approval Local ethics committee at University Greifswald.
-
Provenance and peer review Not commissioned; externally peer reviewed.