Article Text

Abstract
Objective A hallmark of chronic HBV (cHBV) infection is the presence of impaired HBV-specific CD8+ T cell responses. Functional T cell exhaustion induced by persistent antigen stimulation is considered a major mechanism underlying this impairment. However, due to their low frequencies in chronic infection, it is currently unknown whether HBV-specific CD8+ T cells targeting different epitopes are similarly impaired and share molecular profiles indicative of T cell exhaustion.
Design By applying peptide-loaded MHC I tetramer-based enrichment, we could detect HBV-specific CD8+ T cells targeting epitopes in the HBV core and the polymerase proteins in the majority of 85 tested cHBV patients with low viral loads. Lower detection rates were obtained for envelope-specific CD8+ T cells. Subsequently, we performed phenotypic and functional in-depth analyses.
Results HBV-specific CD8+ T cells are not terminally exhausted but rather exhibit a memory-like phenotype in patients with low viral load possibly reflecting weak ongoing cognate antigen recognition. Moreover, HBV-specific CD8+ T cells targeting core versus polymerase epitopes significantly differed in frequency, phenotype and function. In particular, in comparison with core-specific CD8+ T cells, a higher frequency of polymerase-specific CD8+ T cells expressed CD38, KLRG1 and Eomes accompanied by low T-bet expression and downregulated CD127 indicative of a more severe T cell exhaustion. In addition, polymerase-specific CD8+ T cells exhibited a reduced expansion capacity that was linked to a dysbalanced TCF1/BCL2 expression.
Conclusions Overall, the molecular mechanisms underlying impaired T cell responses differ with respect to the targeted HBV antigens. These results have potential implications for immunotherapeutic approaches in HBV cure.
- T lymphocytes
- BCL-2 family proteins
- hepatitis B
- chronic viral hepatitis
- immune response
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
Significance of this study
What is already known on this subject?
HBV-specific CD8+ T cells play a central role in the elimination of HBV infection.
Frequencies of HBV-specific CD8+ T cells in chronic HBV infection are very low reflected by low detection rates applying ex vivo peptide-loaded MHC I (pMHC I) tetramer staining.
Detectable HBV-specific CD8+ T cells are functionally impaired, exhibit mitochondrial alterations, express inhibitory receptors and display dysregulation of T-bet.
What are the new findings?
By applying a pMHC I tetramer-based enrichment strategy, we could detect HBV-specific CD8+ T cells in the majority of chronically HBV-infected patients.
Env183-specific CD8+ T cells are hardly detectable by pMHC I tetramer-based enrichment in chronically HBV-infected patients.
HBV-specific CD8+ T cells targeting core18/141and pol455/173 epitopes are not terminally exhausted but predominantly exhibit a memory-like phenotype in chronic infection.
Core18/141 and pol455/173-specific CD8+ T cells differ in frequency, phenotype and function in chronic HBV infection but not in resolved HBV infection.
Dysregulated TCF1/BCL2 expression limits the expansion capacity of pol455/173-specific CD8+ T cells in chronic HBV infection.
How might it impact on clinical practice in the foreseeable future?
These findings have potential implications for the design of T cell-targeted therapeutic approaches in HBV cure.
Introduction
Virus-specific CD8+ T cells play a central role in the elimination of HBV infection and are thus considered promising targets for immunotherapeutic approaches to cure chronic HBV (cHBV) infection. However, for the design of efficient immunotherapies, a better understanding of HBV-specific CD8+ T cell biology, especially of the dominant antigen targets and their distinct functional characteristics coupled to the respective underlying molecular mechanisms and the corresponding phenotypic profile, is required.
Insights into HBV-specific CD8+ T cell responses were mostly gained by conventional ex vivo peptide-loaded MHC I (pMHC I) tetramer staining and after in vitro expansion. These analyses revealed that the frequencies of HBV-specific CD8+ T cells in cHBV infection are overall very low and associated with the stage of disease.1 Indeed, ex vivo detectability of HBV-specific CD8+ T cells in patients with cHBV is mainly limited to individuals with low viral loads1–7 with detection rates of only 20% of patients even in non-replicative inactive HBsAg carriers.8 The reason for the low detection rate of HBV-specific CD8+ T cells is largely unknown. One possibility might be deletion of HBV-specific CD8+ T cells in patients with cHBV as supported by the finding of elevated expression of the proapoptotic molecule Bim.9 Nevertheless, it has been shown that the few detectable HBV-specific CD8+ T cells are functionally impaired, express several inhibitory receptors5 8 10–14 and display a dysregulation of the transcription factor T-bet.7 These findings are indicative of T cell exhaustion in cHBV infection. Recently, ex vivo transcriptome analyses of HBV-specific CD8+ T cells revealed that mitochondrial alterations contribute to this functional exhaustion.15 However, due to the aforementioned low frequency of HBV-specific CD8+ T cells present in patients with cHBV and the technical limitations of classical immune assays, a comparative knowledge about ex vivo frequencies, molecular profiles and functional characteristics of HBV-specific CD8+ T cells targeting different epitopes is still lacking.
Newly established, highly sensitive technologies have introduced the possibility of comprehensive in-depth analyses of such rare cell populations. Indeed, by applying a pMHC I tetramer-based enrichment strategy, we have recently been able to identify heterogeneous memory-like and terminally exhausted subsets of low frequency HCV-specific CD8+ T cell populations targeting the same epitope.16 Importantly, CD127+PD1+ memory-like HCV-specific CD8+ T cells are maintained after cessation of persistent antigen recognition and harbour the proliferative potential that is determined by the transcription factor TCF1. Here, by adapting the pMHC I tetramer-based enrichment strategy, we were able to detect HBV-specific CD8+ T cells targeting HLA-A*02-restricted polymerase455-463 (pol455) and core18-27 (core18) epitopes in the majority of cHBV patients with low viral load. However, the detection rate of envelope183-191 (env183)-specific CD8+ T cells was low. Subsequent in-depth analyses revealed that HBV-specific CD8+ T cells are not terminally exhausted but rather exhibit a less differentiated, TCF1+ memory-like phenotype in these patients possibly reflecting very weak ongoing cognate antigen recognition. Furthermore, we demonstrate that pol455-specific compared with core18-specific CD8+ T cells are present in different frequencies and exhibit phenotypically and functionally different characteristics that were reproducible also for non-HLA-A*02-restricted core141-160 (core141) and pol173-181 (pol173) epitopes. Specifically, limited expansion of pol455/173-specific but not core18/141-specific CD8+ T cells was linked to dysregulated TCF1/BCL2 expression demonstrating different molecular mechanisms underlying core-specific and pol-specific CD8+ T cell impairment.
Experimental procedures
Study cohort
Eighty-eight patients with cHBV, 10 patients with resolved HBV infection (rHBV) and 6 chronically HCV-infected (cHCV) patients were recruited at the Department of Medicine II of the University Hospital Freiburg, Germany. Viral load and transaminases were determined as part of the clinical diagnostics at the University Hospital Freiburg. Patients with liver cirrhosis were excluded from the study. Detailed patient characteristics are listed in table 1 and online supplementary SI tables 1–4. Twelve healthy donors (HD) served as controls. Human leukocyte antigen (HLA)-typing was performed by next-generation sequencing. Written informed consent was obtained in all cases.
Supplemental material
Study cohort of HLA-A*02-positive cHBV patients with low viral load
Peptides and tetramers
Peptides of HLA-A*02-restricted, HLA-A*11-restricted and HLA-B*35-restricted HBV-derived epitopes (core18: FLPSDFFPSV, pol455: GLSRYVARL, env183: FLLTRILTI, core141: STLPETVVRR, pol173: SPYSWEQEL), HLA-A*02-restricted HCV-derived epitopes (NS31073: CINGVCWTV, NS31406: KLVALGINAV) and HLA-A*02-restricted cytomegalovirus-derived epitope pp65495 (NLVPMVATV) were obtained from Genaxxon, Germany, and dissolved in DMSO (Sigma, Germany). pMHC I tetramers were generated by conjugation of biotinylated pMHC I monomers with phycoerythrin (PE)-conjugated or allophycocyanin (APC)-conjugated streptavidin.
pMHC I tetramer-based magnetic bead enrichment of virus-specific CD8+ T cells
Enrichment of virus-specific CD8+ T cells was performed as previously described.17 Briefly, 2−3×107 peripheral blood mononuclear cells (PBMCs) were labelled for 30 min with PE-coupled or APC-coupled pMHC I tetramers. Subsequent enrichment was performed with anti-PE/APC beads applying MACS technology (Miltenyi Biotec, Germany) according to the manufacturer’s protocol. Frequencies of virus-specific CD8+ T cells were calculated as described previously.17
Expansion of virus-specific CD8+ T cells and assessment of effector function
PBMCs (1−2×106) were stimulated with epitope-specific peptides (10 µg/mL) and anti-CD28 mAb (0.5 µg/mL, BD) and expanded for 14 days in rIL2-containing (20 IU/mL, Miltenyi Biotec) complete RPMI culture medium. The expansion factor was calculated based on pMHC I tetramer staining as previously described.16 Cytokine production and degranulation were assessed after 6-hour restimulation with epitope-specific peptides as described before.16
Statistics
Statistical analysis was performed with GraphPad Prism 6 (USA) and is indicated in the figure legends. Levels of significance are indicated as follows: *p<0.05; **p<0.01; ***p<0.001; ****p<0.0001.
Results
pMHC I tetramer-based enrichment reveals presence of core18-specific and pol455-specific CD8+ T cells in the majority of cHBV patients with low viral load
HBV-specific CD8+ T cells targeting three well-described HLA-A*02-restricted epitopes, core18, pol455, and env183, were analysed in a large cohort of 49 patients with cHBV (HBsAg+, HBeAg−) with low viral load (table 1). Previously, we were only able to detect HBV-specific CD8+ T cells in about 20% of these patients by ex vivo analysis.8 To increase the detection rate and to allow a subsequent comprehensive phenotypic and functional analysis of these rather very low frequency populations, we performed pMHC I tetramer-based enrichment (figure 1A). Indeed, by using this approach, we were able to detect HBV-specific CD8+ T cells in about 80% (39/49) of analysed patients (figure 1B). Specifically, core18-specific and pol455-specific CD8+ T cells were detectable in 87% (20/23) and 66.7% (22/33) of analysed patients, respectively (figure 1C). The higher detection rate of core18-specific CD8+ T cells was also associated with a higher frequency (figure 1D). However, env183-specific CD8+ T cells were only detectable in 25% (8/32) of patients with cHBV (figure 1C), so that further comparative analyses were focused on core18-specific versus pol455-specific CD8+ T cells.

pMHC I tetramer-based magnetic bead enrichment of HLA*A02-restricted HBV-specific CD8+ T cells in patients with low viral load. (A) Representative flow cytometry analyses showing core18-specific, pol455-specific and env183-specific CD8+ T cells obtained from patients with cHBV before and after pMHC I tetramer-based enrichment. Frequency of core18, pol455 and env183 epitope-specific CD8+ T cells within the total CD8+ T cell population is indicated. Proportion of analysed cHBV patients with (depicted in black) and without (depicted in grey) detectable virus-specific (B) and epitope-specific (C) CD8+ T cells after performing pMHC I tetramer-based enrichment. Numbers of analysed patients are indicated. (D) Frequencies of enriched HBV epitope-specific among bulk CD8+ T cells in patients with cHBV. (E) Frequencies of enriched HBV epitope-specific CD8+ T cells with an antigen-experienced (CD45RA+CCR7-, CD45RA-CCR7- or CD45RA-CCR7+) phenotype in patients with cHBV. Dotted line indicates the cut-off value of >80% antigen-experienced cells used for further analyses. Each dot represents one epitope-specific CD8+ T cell population enriched from one individual patient. Median with IQR is indicated. Statistical analyses was performed via Mann-Whitney test (D and E) (*p<0.05; ****p<0.0001). cHBV, chronic hepatatis B virus; pMHC I, peptide-loaded MHC I.
Next, we performed a phenotypical analysis of the enriched core18-specific and pol455-specific CD8+ T cells. Of note, all core18-specific CD8+ T cells showed a phenotype of antigen-experienced cells (being either CD45RA+CCR7−; CD45RA-CCR7−; or CD45RA-CCR7+; figure 1E). In contrast, a subset of pol455-specific CD8+ T cells showed a naive-like phenotype characterised by the coexpression of CD45RA and CCR7. Importantly, the naive-like phenotype of these virus-specific CD8+ T cells is not associated with viral escape mutations and consequent loss of antigen recognition since we only included T cell responses of patients with matched autologous viral sequences in this study. These results indicate that all core18-specific CD8+ T cells have been primed in vivo, while a subset of pol455-specific CD8+ T cells might either not have been primed or may exhibit a naive-like phenotype due to abortive activation. Thus, for comparability reasons, we only included patients in our further analyses that harboured pol455-specific CD8+ T cell responses with >80% antigen-experienced cells.
Increased expansion capacity of core18-specific versus pol455-specific CD8+ T cells
Next, we analysed core18-specific and pol455-specific CD8+ T cells for their expansion capacity and cytokine production. For this, HBV-specific CD8+ T cells were expanded for 2 weeks after stimulation with the respective peptides. As shown in figure 2A,B, core18-specific CD8+ T cells displayed a significantly higher interferon γ (IFNγ) production compared with pol455-specific CD8+ T cells after in vitro expansion. This was also reflected by a better expansion capacity of core18-specific CD8+ T cells (figure 2C). Of note, the expansion capacity of core18- and pol455-specific CD8+ T cells did not correlate with HBsAg levels (online supplementary SI figure 1A). To determine whether the higher IFNγ production was only due to the better expansion capacity or also to increased effector functions of single cells, we determined the multifunctionality, assessed by coexpression of IFNγ, tumour necrosis factor (TNF) and CD107a in relation to the frequency of core18-specific and pol455-specific CD8+ T cells. However, we did not find a statistically significant better multifunctionality of core18-specific CD8+ T cells (figure 2D), indicating that the higher IFNγ production after expansion is most likely reflecting the higher frequency and not increased effector functions. This was further supported by similar antiviral efficacies assessed in coculture experiments of HBV-infected hepatoma cells (HepG2-hNTCP) with either HBV-specific CD8+ T cell clones or patient-derived in vitro expanded HBV-specific CD8+ T cells (online supplementary SI figure 2). Of note, we were not able to assess corresponding antiviral efficacies of HBV-specific CD8+ T cells ex vivo since frequencies were too low for the required effector cell numbers in coculture experiments.18
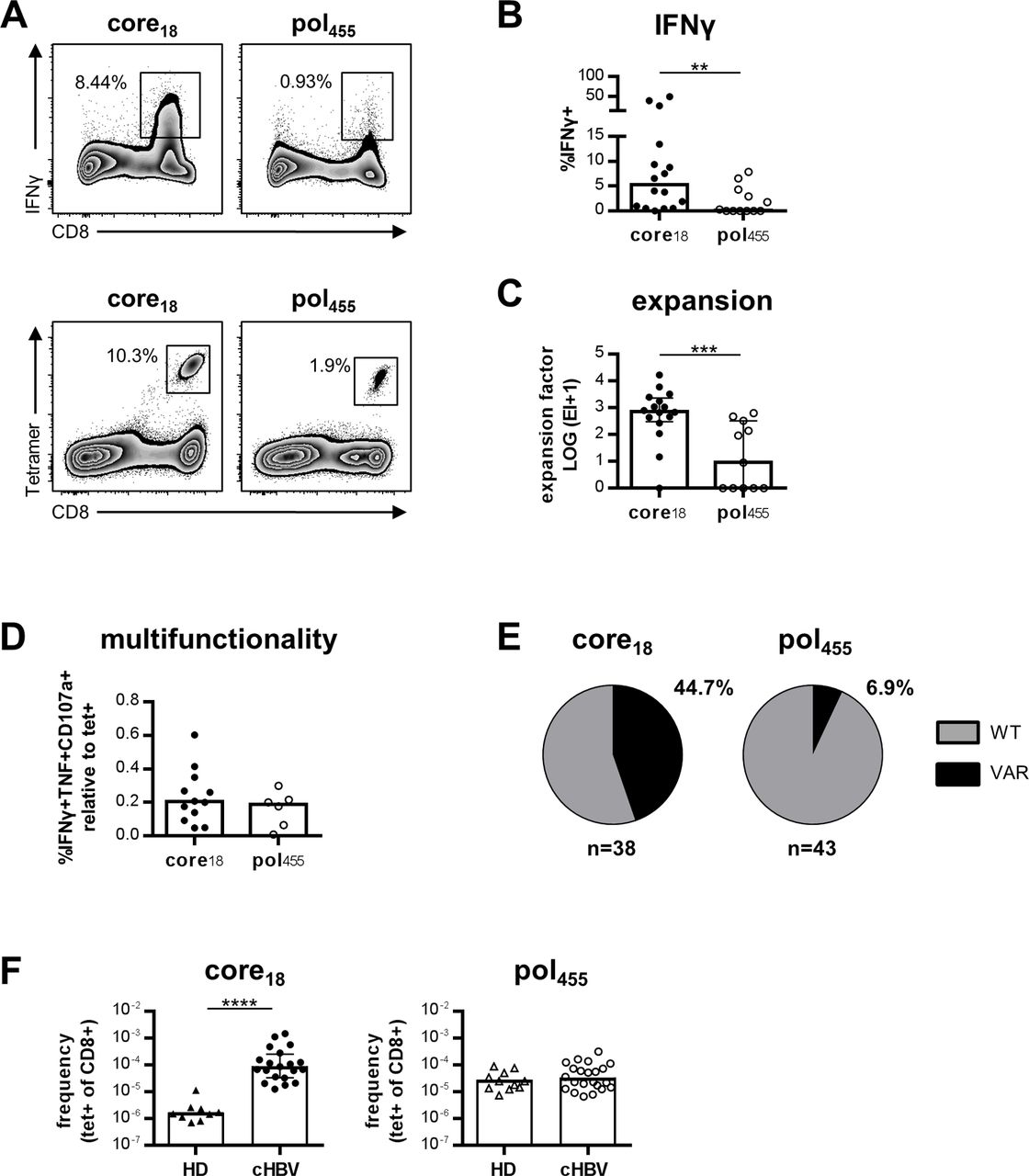
Increased expansion capacity of core18-specific versus pol455-specific CD8+ T cells. (A) Representative flow cytometry analyses showing IFNγ production and tetramer staining of core18-specific and pol455-specific CD8+ T cells obtained from patients with cHBV after peptide-specific in vitro expansion for 14 days. (B) IFNγ production was analysed after restimulation for 5 hours. Frequency of IFNγ+ cells within the CD8+ T cell population is depicted. (C) Calculated expansion factor of core18-specific and pol455-specific CD8+ T cells. (D) Analysis of multifunctionality was performed by assessing the frequency of IFNγ+TNF+CD107a+CD8+ T cells of expanded peptide-specific populations relative to the pMHC I tetramer+ population. (E) Viral sequences of the epitopes targeted by the enriched HBV-specific CD8+ T cell populations were analysed by nested PCR. Proportions of patients harbouring wild type (WT; in grey) and variant (VAR; in black) viral sequences are depicted for HBV core18 and pol455 epitopes, respectively. (F) Frequencies of enriched HBV epitope-specific among bulk CD8+ T cells in HD compared with patients with cHBV. Each dot represents one epitope-specific CD8+ T cell population from one individual patient. Bars indicate the median with IQR. Statistical analysis was performed via Mann-Whitney test (B–D, F) (**p<0.05; ***p<0.001; ****p<0.0001). cHBV, chronic HBV; HD, healthy donors; pMHC I, peptide-loaded MHC I; TNF, tumour necrosis factor.
As a possible indicator of the strength of HBV-specific CD8+ T cell responses in vivo, we analysed the occurrence of viral sequence variations within the respective epitope. Sequence variations were more frequent in the core18 epitope compared with the pol455 epitope (figure 2E) potentially suggesting stronger immune pressure mediated by core18-specific CD8+ T cells. Additionally, to estimate the in vivo expansion of HBV-specific CD8+ T cells after activation, we measured the precursor frequency of HBV-specific CD8+ T cells in HD, compared with the frequencies detectable in patients with cHBV (figure 2F). Of note, we only observed an increased frequency of core18-specific but pol455-specific CD8+ T cells in patients with cHBV compared with HD. This finding indicates a stronger in vivo expansion of core18-specific versus pol455-specific CD8+ T cells in cHBV infection. In sum, these data demonstrate that core18-specific compared with pol455-specific CD8+ T cells are superior in their functional capacity in vitro and in vivo.
Higher KLRG1 and Eomes expression in pol455-specific versus core18-specific CD8+ T cells
Next, we asked whether the functional differences between core18-specific and pol455-specific CD8+ T cells are linked to a differential phenotypical expression of markers indicative of exhaustion in a chronic infection (PD1, 2B4, TIGIT, KLRG1 and CD39). We could detect a higher frequency of KLRG1+ cells in pol455-specific compared with core18-specific CD8+ T cell populations, while the proportion of HBV epitope-specific CD8+ T cells expressing PD1, TIGIT, 2B4 and CD39 was similar (figure 3A and online supplementary SI figure 3A). Interestingly, the expression level of PD1 was increased on core18-specific CD8+ T cells (online supplementary SI figure 3C). However, this was not evident for any other inhibitory receptor. Furthermore, we also analysed expression of the transcriptions factors Eomes and T-bet since high expression of Eomes accompanied by low levels of T-bet has been linked to T cell exhaustion. Here, we found no difference in T-bet but a significantly higher fraction of Eomes+ cells among pol455-specific versus core18-specific CD8+ T cells (figure 3B). The fractions of Eomes+ core18-specific and pol455-specific CD8+ T cells were increased in cHBV compared with patients with rHBV (online supplementary SI table 2, SI figure 3D). Of note, T-bet levels were intermediate in HBV-specific CD8+ T cells targeting core18 and pol455 epitopes (figure 3B; online supplementary SI figure 3B). Thus, a higher fraction of pol455-specific compared with core18-specific CD8+ T cells obtained from patients with cHBV exhibited a typical profile of severe Eomes+T-betint T cell exhaustion that is also in agreement with the lower expansion capacity of these cells (online supplementary SI figure 3E). The profile indicative of T cell exhaustion did not correlate with serum HBsAg levels of analysed patients with cHBV (online supplementary SI figure 1B). Importantly, the phenotypic characteristics of HBV epitope-specific CD8+ T cells were similar when extending our analyses on CD8+ T cell populations targeting the HLA-A*11-restricted core141 and the HLA-B*35-restricted pol173 epitope (online supplementary SI table 1, SI figure 4 and figure 3). Thus, these data clearly suggest that phenotypic profiles of HBV-specific CD8+ T cells in cHBV infection differ with respect to the targeted antigen.

Decreased expression of KLRG1 and Eomes in core18-specific versus pol455-specific CD8+ T cells. Ex vivo expression analyses of inhibitory receptors PD1, TIGIT, 2B4 and KLRG1 (A) and transcription factors Eomes and T-bet (B) in enriched HBV epitope-specific CD8+ T cells obtained from patients with cHBV. (C) Expression of PD1, KLRG1, Eomes and T-bet of HLA-A*02-restricted core18 and pol455-specific CD8+ T cells compared with HLA-A*11-restricted core141 and HLA-B*35-restricted pol173-specific CD8+ T cell populations, respectively. Frequencies refer to cells positive for the respective marker within the indicated HBV-specific CD8+ T cell populations. Naive-like (CD45RA+CCR7+) HBV epitope-specific CD8+ T cells were excluded from marker expression analyses. Each dot represents one epitope-specific CD8+ T cell population enriched from one individual patient. Bars indicate the median with IQR. The following statistical analyses were performed: Mann-Whitney test (A and B: T-bet), unpaired t-test with Welch’s correction (A: KLRG1, B: Eomes) and two-way analysis of variance with Sidak’s multiple comparison test (C) (*p<0.05; **p<0.01).
Core18/141-specific and pol455/173-specific CD8+ T cells differ in their T cell subset distribution in chronic infection
Based on CD127/PD1 coexpression, we could recently define two distinct epitope-specific CD8+ T cell subsets in chronic HCV (cHCV) infection that significantly differ in their functional and phenotypical characteristics: a CD127+PD1+ memory-like population and a more severely exhausted CD127-PD1+ subset (figure 4A). Thus, we asked whether core18/141-specific and pol455/173-specific CD8+ T cells differ in their T cell subset distribution. Interestingly, in comparison with HCV (figure 4A), HBV-specific CD8+ T cells display a different T cell subset distribution during chronic infection with a higher proportion of cells within the memory-like subset (figure 4B). However, within the HBV-specific population, we found significant differences between core18/141-specific and pol455/173-specific CD8+ T cells. Indeed, as shown in figure 4B, core18/141-specific CD8+ T cells were more frequently found in the memory-like T cell subset compared with pol455/173-specific CD8+ T cells. Based on previous findings from our group in cHCV infection showing that antigen stimulation is the main driver of terminal T cell exhaustion and thus contributing to T cell subset distribution, we hypothesised that differential antigen stimulation may also explain the differences between core18/141-specific and pol455/173-specific CD8+ T cells. To address this issue, we analysed the epitope-specific populations for their expression of CD38 as a marker for T cell activation and recent T cell receptor (TCR) triggering. As shown in figure 4C, we found a higher expression of CD38 on pol455/173-specific versus core18/141-specific CD8+ T cells indicating that pol455/173-specific CD8+ T cells are most likely more activated in vivo and that this may lead to a reduced frequency of memory-like CD127+PD1+CD8+ T cells compared with core18/141-specific CD8+ T cells. The overall low expression of CD38 on core18/141-specific and pol455/173-specific CD8+ T cells during cHBV infection cannot be explained by intrinsic T cell defects since HBV epitope-specific TCR stimulation in vitro led to rapid CD38 upregulation (figure 4D). Thus, the low CD38 expression, also in comparison with HCV-specific CD8+ T cells (online supplementary SI table 3, SI figure 5A) is probably due to the low viral load present in our cohort of patients with cHBV. This assumption is further supported by the finding that pol455/173-specific and core18/141-specific CD8+ T cells from patients with cHBV undergoing nucleos(t)ide analogue (NUC) treatment exhibited reduced CD38 expression after NUC-mediated viral control (online supplementary SI table 4 and figure 4). Importantly, reduced CD38 expression after NUC treatment was associated with a higher fraction of memory-like pol455/173-specific CD8+ T cells. Based on the fact that core18/141specific CD8+ T cells have an exclusively memory-like phenotype before NUC treatment, no further increase could be observed (figure 4E). In this context, it is also noteworthy that we did not observe differences in CD38 expression and memory-like CD127+PD1+ subset distribution within our cohort of patients with low viral load when we compared inactive HBsAg carriers and NUC-treated patients (online supplementary SI figure 5B). Hence, antigen recognition in cHBV patients with low viral load is linked to the predominance of memory-like HBV-specific CD8+ T cells but probably differs with respect to the targeted antigen.
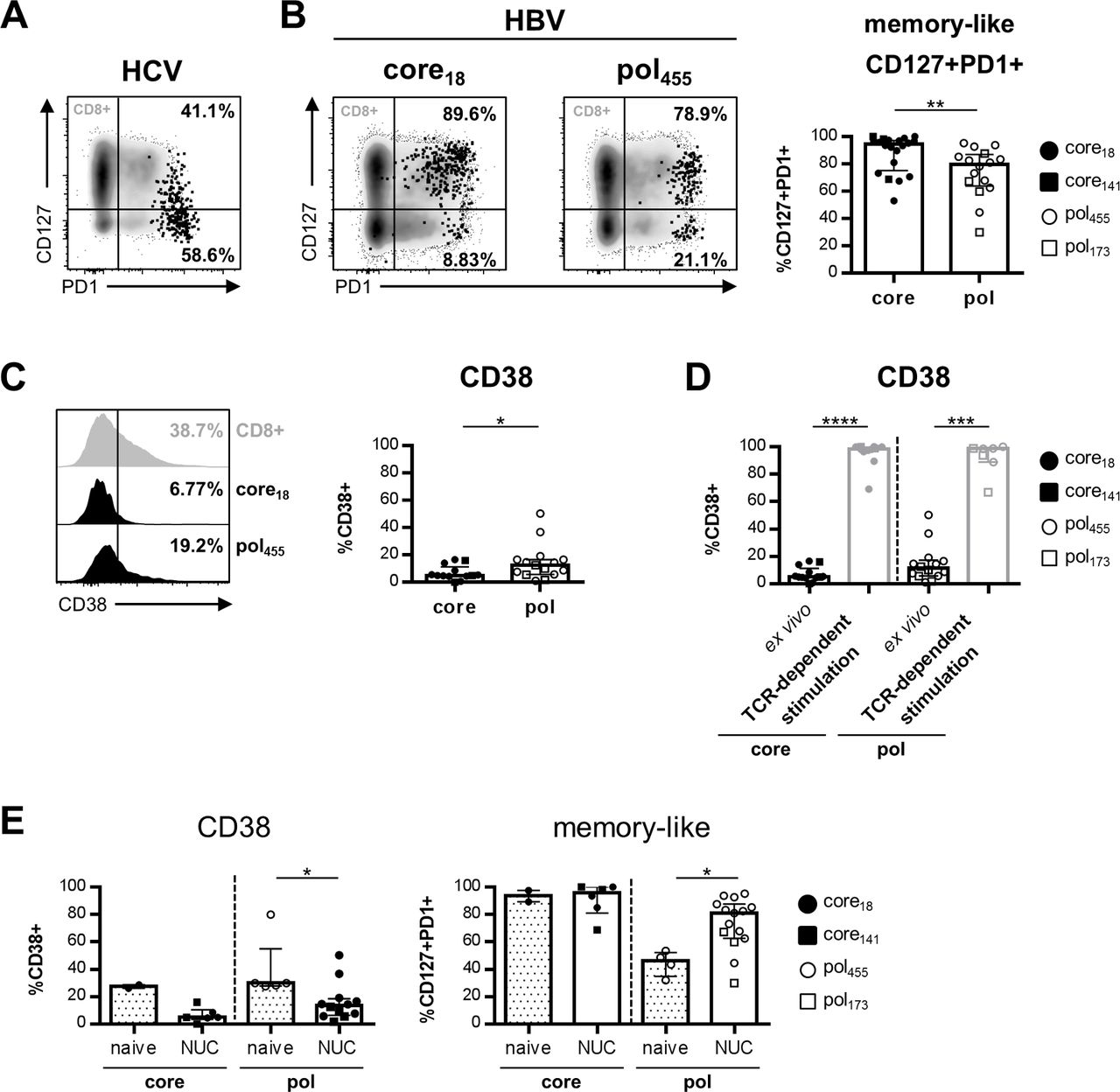
Core18/141-specific and pol455/173-specific CD8+ T cells differ in their CD127/PD1-based subset distribution in low viral load patients. CD127/PD1 coexpression analysis of enriched HCV NS31073-specific (A) and HBV epitope-specific (B) CD8+ T cells obtained from cHCV and cHBV patients, respectively. Frequencies refer to CD127+PD1+ and CD127-PD1+ cells within the epitope-specific CD8+ T cell population. CD38 expression of enriched core18/141-specific versus pol455/173-specific CD8+ T cells from patients with cHBV ex vivo (C) and in comparison with CD38 expression after peptide-specific in vitro expansion (D). (E) Expression of CD38 (left) and proportion of memory-like CD127+PD1+ cells (right) within the HBV epitope-specific CD8+ T cell population in treatment-naive (median viral load: 2.96×106 IU/mL) compared with nucleos(t)ide analogue (NUC)-treated patients (median viral load: 12 IU/mL). Naive-like (CD45RA+CCR7+) epitope-specific CD8+ T cells were excluded from marker expression analyses. Each dot represents one epitope-specific CD8+ T cell population from one individual patient. Bars indicate the median with IQR. Statistical analysis was performed via Mann-Whitney test (B–D) (*p<0.05; **p<0.01; ***p<0.001; ****p<0.0001). cHBV, chronic HBV; cHCV, chronic HCV.
Dysregulated BCL2 expression in pol455/173-specific CD8+ T cells results in defective expansion capacity in cHBV infection
Next, we asked whether the different subset distribution of core18/141-specific versus pol455/173-specific CD8+ T cells may explain the differential expansion capacity of these cells. As shown in figure 5A, we indeed found a clear correlation between the expansion factor and frequency of memory-like core18/141-specific CD8+ T cells. Surprisingly, however, such a correlation was not observed for pol455/173-specific CD8+ T cells (figure 5A). Since the transcription factor TCF1 has been shown to best determine the memory-like phenotype and since it has been linked to the proliferative capacity of this T cell subset,19 20 we next stained both HBV epitope-specific T cell populations for TCF1 expression. We did not find a differential expression of TCF1 within the overall and the memory-like core18/141-specific and pol455/173-specific CD8+ T cell populations (figure 5B) suggesting that the differential expansion capacity cannot be attributed to distinct TCF1-associated proliferative capacities of the respective T cell populations. Next, we asked whether core18/141-specific and pol455/173-specific CD8+ T cells differ in their survival potential. To address this question, we stained HBV epitope-specific CD8+ T cells for the expression of the antiapoptotic molecule BCL2. Importantly, we found a significantly lower expression of BCL2 in pol455/173-specific compared with core18/141-specific CD8+ T cells. The different expression was evident in both the overall antigen-specific CD8+ T cell population and the memory-like subset (figure 5C). We also found a correlation between the expansion factor and the presence of cells highly positive for BCL2 (figure 5D).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Dysregulated TCF1/BCL2 expression determines impaired expansion capacity of pol455/173-specific CD8+ T cells in cHBV patients with low viral load. (A) Correlation analyses of the expansion and the frequency of memory-like CD127+PD1+ core18/141-specific and pol455/173-specific CD8+ T cells. Expression of TCF1 (B) and BCL2 (C) were assessed on enriched core18/141-specific and pol455/173-specific CD8+ T cells (left panel) and the respective memory-like CD127+PD1+ subsets (right panel). (D) Correlation analyses of the expansion and the frequency of BCL2high-expressing HBV-specific CD8+ T cells. HBV-specific CD8+ T cells obtained from rHBV patients were analysed for the expression of TCF1 (left) and BCL2 (middle) and their expansion capacity (right). Naive-like (CD45RA+CCR7+) HBV epitope-specific CD8+ T cells were excluded from marker expression analyses. Each dot represents one epitope-specific CD8+ T cell population from one individual patient. Bars indicate the median with IQR. The following statistical analyses were performed: linear regression analyses (A, D), Mann-Whitney test (B: left panel, D) and unpaired t-test with Welch’s correction (B: right panel, C) (*p<0.05; ***p<0.001). cHBV, chronic HBV; rHBV, resolved HBV.
To dissect whether the dysregulated TCF1/BCL2 expression in pol455/173-specific CD8+ T cells is a hallmark of viral persistence, we further analysed HBV-specific CD8+ T cells obtained from rHBV patients (online supplementary SI table 2). Importantly, we did neither find a dysregulated TCF1/BCL2 expression nor different expansion capacities of pol455-specific versus core18-specific CD8+ T cells after rHBV infection (figure 5E).
These results suggest that the different expansion of core18/141-specific versus pol455/173-specific CD8+ T cells in chronic infection is not primarily due to a differential proliferative capacity but rather to different survival characteristics.
Discussion
By using pMHC I tetramer-based enrichment and thereby increasing the detection sensitivity of HBV-specific CD8+ T cells in the peripheral blood of patients with cHBV, our study gives novel insights into the immunodominance, function and phenotypical properties of the HBV-specific CD8+ T cell response.
The first important finding of our study is that HBV-specific CD8+ T cells are present in the majority of cHBV patients with low viral load. Indeed, we were able to detect HBV-specific CD8+ T cells ex vivo in about 80% of patients clearly suggesting that the previously reported low detection rate of core18-specific and pol455-specific CD8+ T cells1–6 8 cannot be explained by a complete deletion of virus-specific CD8+ T cells. Our finding is in agreement with a recent study from the group of Bertoletti also reporting the common existence of HBV-specific CD8+ T cells in a patient cohort with low or undetectable viral load due to antiviral therapy with NUCs.21 Importantly, env183-specific CD8+ T cells were only detectable in a minor fraction of tested cHBV patients with low viral loads suggesting that limited presence of env183-specific CD8+ T cells may impair efficient responses in cHBV infection.
The higher detection rate of antigen-specific CD8+ T cells by pMHC I tetramer-based enrichment revealed previously unappreciated differences between core18-specific and pol455-specific CD8+ T cells. In particular, core18-specific compared with pol455-specific CD8+ T cells were present in a higher frequency and displayed a predominant antigen-experienced phenotype in all tested patients indicating a higher immunogenicity of the HBV core protein. In contrast, some pol455-specific CD8+ T cells displayed a naive-like phenotype despite the presence of cognate antigen. Thus, as has also been shown in HCV infection,22 these results reveal the presence of phenotypically diverse HBV-specific CD8+ T cells in cHBV infection. They also suggest that some pol455-specific CD8+ T cells are inefficiently recruited to the effector T cell pool and present a naive-like phenotype due to abortive activation. This is further supported by the reduced in vivo expansion of pol455-specific CD8+ T cells that result in low frequencies hardly differing from naive precursor frequencies. The mechanisms responsible for the different frequencies and phenotypes of core18-specific and pol455-specific CD8+ T cells remain unclear. The overall cytokine milieu in patients with cHBV was most likely not responsible for the occurrence of naive-like HBV-specific CD8+ T cells since we were able to detect HBV epitope-specific CD8+ T cell populations with distinct phenotypes, more specifically antigen-experienced core18-specific and naive-like pol455-specific CD8+ T cells, in the same cHBV donor. Noteworthy, HBV core and pol proteins are produced in different quantities in HBV-infected cells with significantly higher levels of core.23 Since the level of antigen presented on hepatocytes may dictate T cell frequencies23 as well as T cell function,24 it is tempting to speculate that this contributes to the higher priming capacity and subsequent higher frequency of core18-specific versus pol455-specific CD8+ T cells. In addition, differences in antigen processing and presentation as well as in HLA binding and TCR affinity may also influence the frequencies, phenotype and function of HBV-specific CD8+ T cells. Further studies are definitely required to clarify this point, for example, by analysing T cell responses towards HBV-infected target cells. In this context, it is interesting to note that in comparison with the pol455 peptide, the core18 peptide has been reported to have a higher HLA binding affinity and an increased cross-reactive behaviour.25 In line with this observation, core18-specific CD8+ T cell responses can be detected in the context of different HLA-A*02 alleles25 26 supporting a highly cross-reactive priming capacity of core18. Clearly, future studies have to address how the finding of different frequencies and phenotypes of circulating core18-specific versus pol455-specific CD8+ T cells translates into the intrahepatic compartment. In this regard, it will be important to dissect antigen-specific and tissue-specific influences27 on intrahepatic HBV-specific CD8+ T cells.
We also observed functional differences between the two HBV epitope-specific CD8+ T cell populations with core18-specific CD8+ T cells showing a significantly better expansion capacity. This was not reflected by the fraction of PD1+ cells since PD1 was detectable on nearly all HBV-specific CD8+ T cells irrespective of their antigen specificity. Thus, this finding supports the assumption that PD1 is not a definitive marker for complete functional exhaustion but can also be expressed on at least partially functional T cells in the context of a chronic infection.16 19–21 28 29 Furthermore, in the lymphocytic choriomeningitis virus mouse model, PD1 has even been shown to prevent virus-specific CD8+ T cells from terminal exhaustion and to contribute to the survival of memory T cell populations.30 In line with this, elevated PD1 expression levels on core18-specific CD8+ T cells may be linked to their otherwise less severely exhausted characteristics compared with pol455-specific CD8+ T cells.
In addition to PD1, we also detected expression of inhibitory receptors such as 2B4, KLRG1 and TIGIT on a large fraction of both HBV epitope-specific CD8+ T cell populations. These results are in agreement with several previous studies reporting that HBV-specific CD8+ T cells in cHBV infection are characterised by expression of PD1 and other inhibitory receptors indicative of CD8+ T cell exhaustion.5 8 10–14 Interestingly, in comparison with core18-specific CD8+ T cells, significantly more pol455-specific CD8+ T cells expressed KLRG1 suggesting further antigen-mediated differentiation.31 32 In addition, we also found differences between core18-specific and pol455-specific CD8+ T cells on the transcription factor level. In fact, we observed a higher Eomes expression in pol455-specific CD8+ T cells accompanied by an equally low T-bet expression. Hence, in cHBV infection pol455-specific compared with core18-specific CD8+ T cells exhibit dysregulated Eomes/T-bet expression ratios that more closely resemble the pattern that has been associated with terminal exhaustion.29 Of note, in rHBV infection, Eomes expression of HBV-specific CD8+ T cells was reduced compared with cHBV patients with an even lower fraction of Eomes+ pol455-specific CD8+ T cells. The fraction of Eomes-expressing HBV-specific CD8+ T cells correlated with their expansion capacity, further supporting the association of dysregulated Eomes expression and T cell dysfunction in cHBV infection. Importantly, the phenotypic and functional differences of HBV core-specific and pol-specific CD8+ T cells were similar comparing HLA-A*02 and non-HLA-A*02-restricted epitopes in cHBV infection and are therefore most probably determined by the targeted antigen.
Exhausted virus-specific CD8+ T cell populations in chronic infections are not homogeneous but rather consist of heterogeneous subsets of less differentiated progenitor/memory-like and further differentiated/terminally exhausted cells.16 19 20 28 29 33 Interestingly, memory-like cells predominated in all tested HBV epitope-specific CD8+ T cell populations of our cohort with low viral loads probably reflecting low ongoing antigen stimulation. However, in line with the advanced differentiation towards terminal exhaustion reflected by a higher proportion of KLRG1 and Eomes expressing cells, we also found a decreased frequency of CD127+PD1+ memory-like cells among CD8+ T cells targeting pol455/173 compared with core18/141. In HCV infection, we have previously reported that this memory-like subset is maintained after antigen removal on treatment with direct-acting antivirals but that ongoing antigen triggering diminishes the frequency of this subset.16 Thus, the decreased frequency of memory-like pol455/173-specific CD8+ T cells indicates stronger ongoing antigen stimulation of these cells compared with core18/141-specific CD8+ T cells. The higher antigen triggering of pol455/173-specific CD8+ T cells is further supported by the higher CD38 expression observed in our study. In sum, based on the distinct expression of KLRG1, Eomes and CD38 and reflected by the differential CD127/PD1-based T cell subset distribution, our results indicate that pol455/173-specific CD8+ T cells probably receive more sustained stimulation and are thus more severely exhausted in vivo compared with core18/141-specific CD8+ T cells. In this context, it is also important to note that we observed, in line with previous studies,34 a higher mutation rate within the core18 epitope versus the pol455 epitope further indicating a higher pol455-specific T cell exhaustion and thus reduced viral evolution pressure by pol455-specific CD8+ T cells also in patients with cHBV.
The frequency of the memory-like CD127+PD1+ subset clearly correlated with the expansion capacity of core18/141-specific CD8+ T cells. However, no correlation was evident with regard to pol455/173-specific CD8+ T cells. This was a surprising finding since expansion capacity has been previously associated with the frequency of memory-like virus-specific CD8+ T cells in HCV infection.16 In HCV infection, the memory-like HCV-specific CD8+ T cell subset is determined by the expression of TCF1 that has been linked to the proliferative capacity of these cells.16 Interestingly, core18/141-specific and pol455/173-specific CD8+ T cells and the respective memory-like CD127+PD1+ subsets expressed TCF1 at equal frequencies. Thus, TCF1 does probably not exclusively determine the differential expansion capacities of HBV epitope-specific CD8+ T cell populations. Next to proliferation, virus-specific CD8+ T cell expansion requires T cell survival. Of note, we observed lower expression of the antiapoptotic molecule BCL2 in pol455/173-specific versus core18/141-specific CD8+ T cells and the expression of BCL2 correlated with the expansion of the two HBV-specific CD8+ T cells. These results therefore clearly suggest that the differential BCL2 expression is linked to the different expansion capacities of core18/141-specific versus pol455/173-specific CD8+ T cells and pinpoint to distinct molecular mechanisms of HBV-specific CD8+ T cell failure targeting cognate core and pol antigens. Furthermore, these data also reveal that a balanced expression of TCF1 and BCL2 is essential for the superior expansion characteristics of memory-like subsets among exhausted virus-specific CD8+ T cells in general. Thus, the combined capacity to proliferate and to survive determines memory-like characteristics of T cells in chronic infection.
In sum, the results of our study reveal on the one hand that, in cHBV infection, the molecular mechanisms underlying impaired circulating T cell responses differ with respect to the targeted HBV antigens core versus pol. However, regarding chronic infection in general, we specified key characteristics of targetable virus-specific CD8+ T cell subsets in greater depth, namely proliferation accompanied by survival. Both aspects have relevance for the design of future immunotherapeutic approaches in HBV cure that aim to expand CD8+ T cell responses within the peripheral blood, the best accessible site for intervention.
Acknowledgments
We would like to thank all patients and donors for participating in the current study.
References
Footnotes
ESA and KH contributed equally.
MH and RT contributed equally.
Contributors AS designed, performed and analysed experiments and wrote the manuscript; ESA, KH, DW, MMK, JK and OS performed experiments; YN and PZ participated in developing experimental procedures; SL-L, SU, MN and DAP provided reagents; BB, CN-H and HL contributed to data interpretation; FE conducted HLA genotyping; MH and RT designed the study, contributed to experimental planning, interpreted data and wrote the manuscript.
Funding This work was supported by the SFB 1160/IMPATH (Project 08) of the German Research Foundation (DFG) to RT, by the SFB 1160/IMPATH (Project 10) of the DFG to CN-H, by the SFB/TRR179 (TP16) of the DFG to MN and by the SFB/TRR179 (TP15) of the DFG to SU. MH was supported by a DZIF maternity leave stipend (TI 07.005_Hofmann) and by the SFB/TRR179 (TP01) of the DFG.
Competing interests None declared.
Patient consent Not required.
Ethics approval The study was conducted according to federal guidelines, local ethics committee regulations (Albert-Ludwigs-University, Freiburg, Germany, HBUF 474/14 and 299/01) and the Declaration of Helsinki (1975).
Provenance and peer review Not commissioned; externally peer reviewed.
Data sharing statement All data are published in the manuscript.