Article Text

Abstract
Objective To study the role of α4β7 integrin for gut homing of monocytes and to explore the biological consequences of therapeutic α4β7 inhibition with regard to intestinal wound healing.
Design We studied the expression of homing markers on monocyte subsets in the peripheral blood and on macrophage subsets in the gut of patients with IBD and controls with flow cytometry and immunohistochemistry. Integrin function was addressed with dynamic adhesion assays and in vivo gut homing assays. In vivo wound healing was studied in mice deficient for or depleted of α4β7 integrin.
Results Classical and non-classical monocytes were clearly dichotomous regarding homing marker expression including relevant expression of α4β7 integrin on human and mouse non-classical monocytes but not on classical monocytes. Monocyte-expressed α4β7 integrin was functionally important for dynamic adhesion to mucosal vascular addressin cell adhesion molecule 1 and in vivo gut homing. Impaired α4β7-dependent gut homing was associated with reduced (effect size about 20%) and delayed wound healing and suppressed perilesional presence of wound healing macrophages. Non-classical monocytes in the peripheral blood were increased in patients with IBD under clinical treatment with vedolizumab.
Conclusion In addition to reported effects on lymphocytes, anti-α4β7 therapy in IBD also targets non-classical monocytes. Impaired gut homing of such monocytes might lead to a reduction of wound healing macrophages and could potentially explain increased rates of postoperative complications in vedolizumab-treated patients, which have been observed in some studies.
- gut immunology
- ulcerative colitis
- surgery for IBD
- inflammatory bowel disease
- Crohn’s disease
Statistics from Altmetric.com
Significance of this study
What is already known on this subject?
The anti-α4β7 integrin antibody vedolizumab is successfully used for clinical treatment of Crohn’s disease and ulcerative colitis.
Mechanistically, anti-α4β7 integrin antibodies are considered to prevent gut homing of disease-driving lymphocytes.
Wound healing macrophages support tissue remodelling in intestinal wound healing.
What are the new findings?
A relevant fraction of non-classical monocytes expresses α4β7 integrin.
Monocyte-expressed α4β7 integrin is functionally important for dynamic adhesion and gut homing.
Impaired α4β7-dependent gut homing leads to delayed experimental wound healing and reduces perilesional presence of wound healing macrophages.
How might it impact on clinical practice in the foreseeable future?
As α4β7 blockade may affect intestinal wound healing, careful counselling of vedolizumab-treated patients and timing of surgical procedures might be required.
Introduction
The pathogenesis of the IBDs, Crohn’s disease (CD) and ulcerative colitis (UC) is still far from being understood. However, multiple factors contributing to their development have been identified, including an aberrant response of the intestinal immune system.1 2 Elucidation of communication and trafficking of intestinal immune cells has led to the development of novel biological therapies for the treatment of IBD.3–5
In particular, the anti-α4β7 integrin antibody vedolizumab is routinely used for the clinical treatment of IBD since 2014.6 7 On a mechanistic level, it is well established that vedolizumab impedes adhesion of α4β7 integrin-expressing lymphocytes to mucosal vascular addressin cell adhesion molecule 1 (MAdCAM-1) expressed on high endothelial venules in the gut.8 9 This prevents so-called ‘gut homing’—the process of extravasation of cells from the blood to the intestinal tissue10—eventually leading to decreased lymphocyte infiltration to the gut and, thus, reduced inflammation.
Yet, beyond this, several aspects of the mechanism of action of vedolizumab are still unclear. For example, the functional effects on other immune cells expressing α4β7 that crucially contribute to inflammation in CD and UC are not well understood.11 Particularly, macrophages are considered to prominently contribute to the inflammatory network in IBD.12 13 Intestinal macrophages arise from monocytes circulating through the peripheral blood and homing to the gut tissue.14 15 Depending on the local environment such macrophage differentiation can lead to development of M1-like macrophages secreting proinflammatory cytokines like tumour necrosis factor alpha (TNF-α), interleukin (IL)-12 or IL-23 or to development of alternatively activated M2-like macrophages producing anti-inflammatory mediators like transforming growth factor beta (TGF-β) or IL-10.16 While the former have been shown to contribute to active inflammation, the latter are deemed important for tissue homeostasis, remodelling and wound healing.17 This dichotomous model has recently been expanded to a spectrum model of a whole variety of macrophage activation states, of which M1-like inflammatory macrophages and M2-like alternatively activated macrophages are the extremes.18
Monocyte homing to the gut has not been characterised in detail so far. In general, the integrins αLβ2 and αMβ2 are considered as important adhesion molecules for monocyte homing.19 However, whether α4β7 integrin might also play a role in monocyte gut homing has never been investigated. Moreover, it is known that distinct subsets of peripheral blood monocytes—classical, intermediate and non-classical monocytes—exist,20 but whether these subsets behave differently with regard to gut homing has not been explored to date. Finally, the relation between peripheral blood monocyte subsets and macrophage subsets in the gut has not been addressed so far.
Here, we show with human cells and tissues as well as in in vivo mouse models, that a relevant fraction of non-classical but not classical monocytes expresses α4β7 integrin, which is functionally important for dynamic adhesion to MAdCAM-1 and in vivo gut homing. Furthermore, we demonstrate that non-classical monocytes are progenitors of wound healing macrophages upregulated in the proliferative phase of wound healing, but decreased following anti-α4β7 integrin antibody treatment. Taken together, our data uncover previously unrecognised effects of anti-α4β7 therapy in IBD and experimental wound healing with potential clinical relevance in perioperative settings.
Methods
Methods are described in the online supplementary methods. Clinical characteristics of patients contributing material to the study are summarised in table 1.
Supplemental material
Patient characteristics
Results
α4β7 integrin is expressed on a portion of intermediate and non-classical monocytes
The expression of α4β7 integrin on human monocytes in general and human monocyte subsets in particular is unclear. We therefore quantified the expression in CD14++CD16− classical, CD14++CD16+ intermediate and CD14+CD16++ non-classical monocytes in the peripheral blood of patients with IBD and controls.
The classical subset was clearly dominating (online supplementary figure 1A), but it was reduced in patients with CD and UC compared with healthy controls. Conversely, the intermediate subset was significantly increased in UC (figure 1A). While classical monocytes expressed virtually no α4β7 integrin, coexpression of α4 and β7 integrin could be demonstrated on around 5% of intermediate and non-classical monocytes (figure 1B). This was comparable in patients with CD and UC and controls. We confirmed our results in a further set of analyses, in which we excluded CD56+natural killer cells, which can also express CD16, to exclude an impact of these cells on our results (data not shown). Finally, we performed further experiments with additional dye-based exclusion of dead cells and exclusion of CD3 and CD19-expressing cells within the monocyte gate (online supplementary figure 1B). Consistently, all data demonstrated increased expression of α4β7 on a portion of non-classical and intermediate monocytes, but not classical monocytes.
Supplemental material

α4β7 integrin is expressed on intermediate and non-classical monocytes in the peripheral blood. (A) Left panel: representative gating of CD14++CD16− classical (CLM), CD14++CD16+ intermediate (INM) and CD14+CD16++ non-classical monocytes (NCM) after previous exclusion of doublets and selection of monocytes based on forward and sideward scatter. Right panels: proportion of classical, intermediate and non-classical monocyte subsets among overall monocytes in control donors (CON, n=23) and patients with Crohn’s disease (CD, n=37) and ulcerative colitis (UC, n=22). (B) Expression of α4β7 integrin on classical, intermediate and non-classical monocytes in control donors and patients with IBD (n=18–26) as indicated. Left: quantitative flow cytometry. Right panels: representative flow cytometry.
Since coexpression of α4 and β7 could potentially also be due to concomitant expression of α4β1 and αEβ7 in the absence of α4β7 integrin heterodimers, we performed an additional series of experiments with directly labelled vedolizumab to detect the α4β7 dimer (online supplementary figure 2A). Indeed, heterodimer expression was increased on intermediate and even more on non-classical monocytes compared with absent expression on classical monocytes, supporting the view that a fraction of non-classical but not classical monocytes expresses α4β7 integrin and might thus be a target of clinical therapy with vedolizumab.

Different homing marker profiles in non-classical versus classical monocytes. Expression of αLβ2 (A), GPR15 (B), αMβ2 (C) and α4β1 (D) integrins on classical, intermediate and non-classical monocytes in control donors and patients with IBD (n=11–31) as indicated. Left: quantitative flow cytometry. Right panels: representative flow cytometry. CD, Crohn’s disease; CLM, classical monocyte; CON, control; INM, intermediate monocyte; NCM, non-classical monocyte; UC, ulcerative colitis.
Different homing marker profiles on non-classical versus classical monocytes
Next, we expanded our analyses to further evaluate homing markers on peripheral blood monocyte subsets. It appeared that non-classical and classical monocytes seem to be dichotomous with regard to homing marker expression, since ‘traditional’ monocyte homing markers like αLβ2 integrin and αMβ2 integrin were expressed on virtually all classical monocytes but only on a fraction of non-classical monocytes. Indeed, markers important for intestinal lymphocyte homing or adhesion like α4β1 and GPR15 were expressed on considerably more non-classical than classical monocytes (figure 2, online supplementary figure 2B).
Taken together, these expression data suggested that distinct homing marker profiles in different peripheral blood monocyte subsets might potentially be associated with differential functional gut homing strategies.
Monocyte-expressed α4β7 integrin is functional for dynamic adhesion and in vivo homing to the gut
While the homing molecules predominantly expressed on classical monocytes have an established functional role for monocyte homing,19 a relevance of α4β7 integrin in this context has not been demonstrated so far. Thus, we assessed, whether these integrins, which have an important function in lymphocyte gut homing, might have similar implications for monocytes.
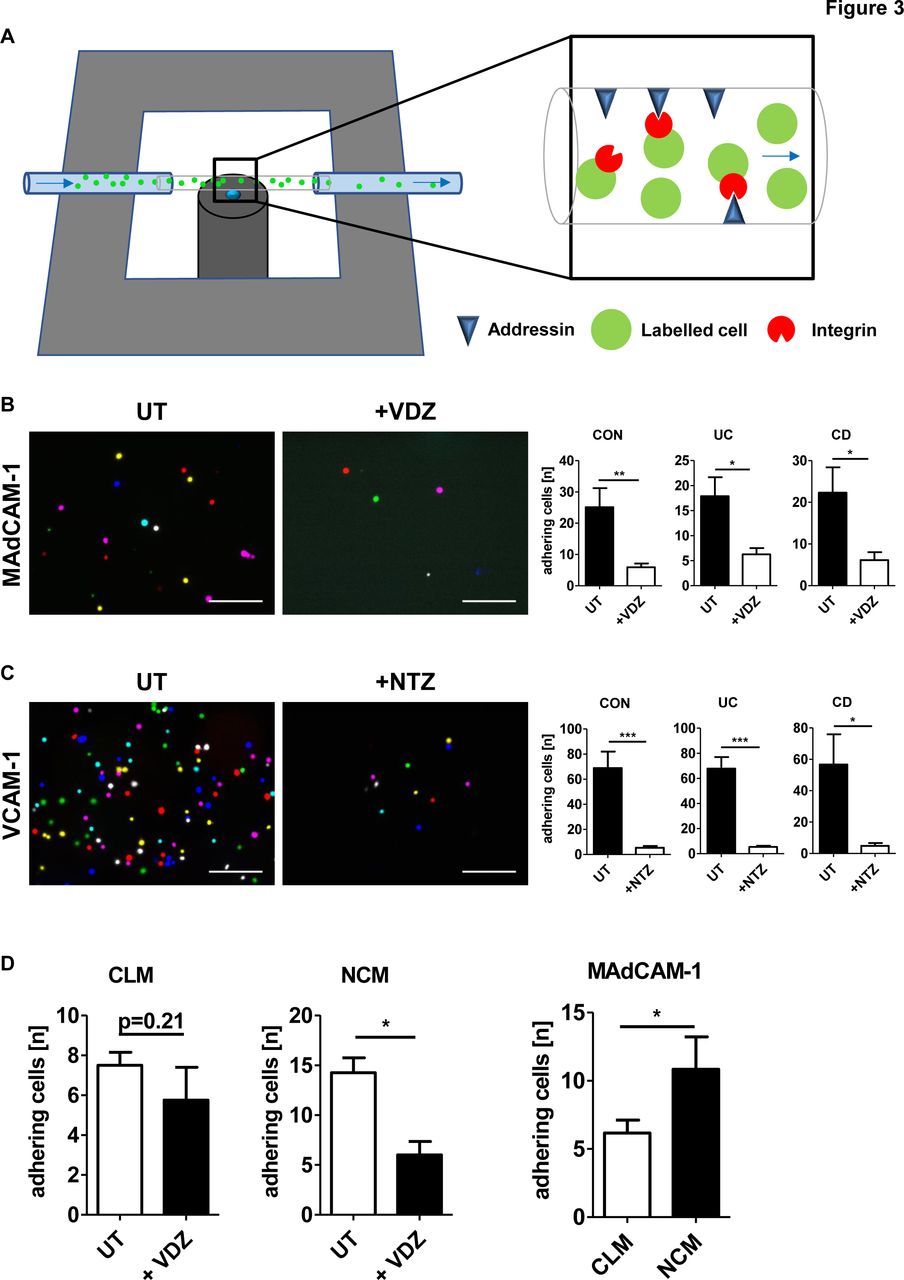
To this end, we performed functional dynamic adhesion assays (figure 3A). We purified peripheral blood monocytes from patients with IBD as well as healthy donors, labelled them with carboxyfluorescein succinimidyl ester (CFSE) and perfused them through ultrathin glass capillaries coated with or without MAdCAM-1 and vascular cell adhesion molecule 1 (VCAM-1) after treatment with or without the anti-α4β7 antibody vedolizumab and the anti-α4 antibody natalizumab, respectively.

Monocyte-expressed α4β7 integrin is functional for dynamic adhesion. (A) Schematic depiction of the principle of dynamic adhesion assays. Addressin-coated ultrathin glass capillaries are perfused with a labelled cell suspension and integrin-dependent dynamic adhesion to addressins is analysed by microscopy. (B, C) Quantitative data from experiments with magnetic activated cell sorting (MACS)-isolated CD14+ monocytes treated without (UT) or with vedolizumab (VDZ, B) and without (UT) or with natalizumab (NTZ, C) and perfused through capillaries coated with mucosal vascular addressin cell adhesion molecule 1 (MAdCAM-1) (B) and vascular cell adhesion molecule 1 (VCAM-1) (C), respectively. Left panels: representative merged images from eight high-power fields; scale bar 100 µm. Right panels: quantification of dynamic adhesion of cells from control donors and patients with IBD (n=5–11). (D) Quantitative data from experiments with FACS-sorted CD14++CD16− classical (CLM) and CD14+CD16++ non-classical (NCM) monocytes treated without (UT) or with VDZ. Left panels: adhesion of CLM and NCM with and without treatment. Right panel: quantification of adhering untreated cells (n=4–6). CD, Crohn’s disease; CON, control; UC, ulcerative colitis.
Dynamic adhesion of monocytes from patients with IBD as well as control patients to both MAdCAM-1 and VCAM-1 could be noted in adhesion molecule-coated but not in control capillaries indicating that monocytes may indeed adhere to these ligands. Treatment with vedolizumab reduced monocyte adhesion to MAdCAM-1 to background adhesion as did natalizumab regarding adhesion to VCAM-1 (figure 3B,C). Of note, vedolizumab and natalizumab did not increase apoptosis or necrosis of monocytes (data not shown). These data suggested that monocyte adhesion to MAdCAM-1 and VCAM-1 is specifically mediated by α4 integrins and that the effects of vedolizumab and natalizumab extend beyond lymphocytes and also include monocytes expressing α4β7 and α4, respectively.
Next, we aimed to confirm the functional relevance of differential α4β7 expression on monocyte subsets. For this purpose, dynamic adhesion assays with FACS-sorted CD14++CD16− classical and CD14+CD16++ non-classical monocytes perfused through MAdCAM-1 coated capillaries were performed. Dynamic adhesion to MAdCAM-1 was significantly higher with non-classical compared with classical monocytes. Further, treatment with the anti-α4β7 antibody vedolizumab led to a significant reduction of dynamic adhesion of non-classical monocytes, while the adhesion of classical monocytes was not affected (figure 3D). Taken together, these data suggested functional implications of differential homing marker profiles on monocyte subsets.
We also aimed to evaluate the role of α4β7 for gut homing in vivo and with specific regard to monocyte subsets. In order to analyse appropriate quantities of cells, we used mouse CX3CR1high non-classical and CX3CR1low classical monocytes, which are considered to functionally correspond to the human non-classical and classical monocyte subsets.21 Thus, we analysed the expression of α4 and β7 integrin on these cells in mice and found an increased coexpression on non-classical compared with classical cells (figure 4A, online supplementary figure 3A), suggesting that differential α4β7 integrin expression on monocyte subsets is shared between humans and mice.

Monocyte-expressed α4β7 integrin is functional for in vivo homing. (A) Expression of α4β7 integrin on mouse CX3CR1low classical (CLM) and CX3CR1high non-classical monocytes (NCM). Left: quantitative flow cytometry data (n=16). Right: representative flow cytometry including fluorescence minus one (FMO) controls. (B) In vivo gut homing of NCM after adoptive transfer to the ileocolic artery. Representative light sheet fluorescence microscopy. Red: anti-CD31, green: CellTrace dyes, grey: autofluorescence. Left panels: overview in blend mode (left) and in maximum intensity projection (second from left); scale bar 300 µm. Right panels: detailed images of the indicated 3D cube as maximum intensity projection (second from right) and in blend mode (right); scale bar 50 µm. Green arrows indicate green extravasated cells, yellow arrows indicate cells within the vessels (yellow overlay with red vasculature). (C) In vivo homing of CLM and NCM after adoptive transfer to the ileocolic artery together with anti-α4β7 or isotype antibody treatment. Left upper panels: representative in vivo confocal microscopy. White arrows highlight extravasated cells, orange arrows highlight cells within the vessels. Right upper panels: representative flow cytometry of CFSE+ monocytes in the lamina propria of adoptively transferred mice after 1 hour. Left lower panel: quantitative flow cytometry of the accumulation of untreated NCM versus CLM. Right lower panels: quantitative flow cytometry of the accumulation of treated versus untreated CLM and NCM (n=5). CFSE, carboxyfluorescein succinimidyl ester; SSC, sideward scatter.
For the analysis of in vivo homing, we adapted a previously established in vivo gut homing model.22 23 In a first step, we cultured bone marrow cells to macrophage colony-stimulating factor-differentiated monocytes, which were subsequently sorted to classical and non-classical monocytes (online supplementary figure 3B,C). These cells were fluorescently labelled and transferred to the ileocolic artery of recipient mice together with anti-α4β7 integrin antibody or isotype control antibody treatment and in addition to recipient vessel staining with Texas Red Dextran or anti-CD31 antibodies. Gut homing of transferred monocyte subsets was then analysed by light sheet fluorescence microscopy,23 intravital confocal microscopy and flow cytometry of lamina propria mononuclear cells (LPMC) (figure 4B,C, online supplementary figures 3D and 4). Our microscopic data indicated extravasation of non-classical monocytes to the gut tissue and, by flow cytometry, we found that significantly more isotype-treated non-classical than classical monocytes homed to the gut. Furthermore, anti-α4β7 integrin treatment led to significantly reduced colonic accumulation of non-classical monocytes, while effects in classical monocytes were less marked and not statistically significant. These results supported the notion that α4β7 expressed on non-classical monocytes is functionally important for mediating dynamic adhesion and in vivo gut homing.
Non-classical monocytes are skewed towards wound healing macrophage commitment in the gut
In view of these data, we wondered what implication the inhibition of α4β7 integrin-mediated gut homing of non-classical monocytes might have for macrophage composition and function in the gut.
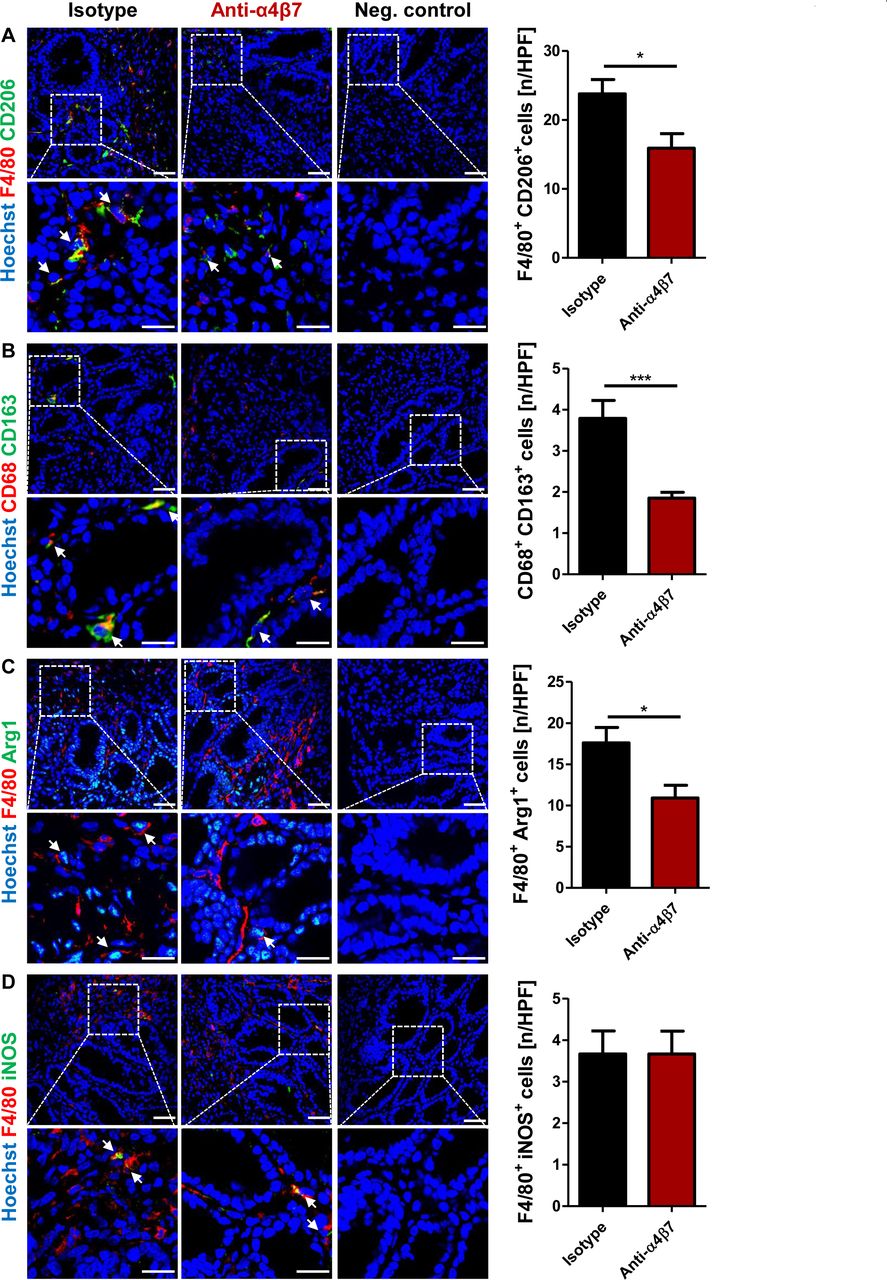
Several pieces of evidence from non-intestinal tissues suggest that non-classical monocytes preferentially develop into wound healing macrophages in peripheral tissues,24 25 which have an important role in the proliferative phase of wound healing.26 Thus, we hypothesised that this might also be the case in the gut. Accordingly, we used a previously reported technique to label non-classical monocytes in mice with polychromatic red (PR)-coupled latex beads.24 Subsequently, colonic wounds were inflicted in these mice and LPMCs from wounded areas were collected 5 days later and stained for macrophage markers (figure 5A). While the specificity of the labelling for non-classical monocytes was very high (figure 5B, online supplementary figure 5A), we could demonstrate that PR+ intestinal macrophages expressed significantly higher levels of CD206, CD163 and arginase, but not inducible nitric oxide synthase (iNOS) than PR− intestinal macrophages (figure 5C). These data suggested that non-classical monocytes preferentially develop into wound healing macrophages.

Preferential development of non-classical monocytes to wound healing macrophages in the intestine. (A) Schematic depiction of experimental set-up. Mice were injected intravenously with polychromatic red (PR)-coupled latex beads (LTX-PR) on day −1. Twenty-four hours later, blood was collected to analyse monocyte labelling and wounds were inflicted to their colon. On day 5, lamina propria mononuclear cells (LPMC) from wounded areas were isolated and stained for flow cytometry (FC) to address the fate of non-classical monocytes in experimental intestinal wound healing. (B) Monocyte subset composition among PR− and PR+ monocytes on day 0. Representative flow cytometry. (C) Expression of markers for inflammatory and wound healing macrophages among PR− and PR+ intestinal F4/80+ macrophages from wounded areas on day 5. Representative and quantitative flow cytometry of CD206 (upper left panels), CD163 (upper right panels), arginase 1 (bottom left panels) and inducible nitric oxide synthase (iNOS; bottom right panels). n=9. (D, E) Expression of β7 integrin on CD14+ and CD163+ cells in the colon of patients with IBD and controls. (D) Representative immunohistochemistry, arrows highlight double positive cells, (E) quantification of the fraction of β7+ cells among CD14+ and CD163+ cells (n=4–8); scale bars 50 µm (upper panels), 25 µm (lower panels). (F) Expression of α4 integrin on CD14+ and CD163+ cells in the colon of patients with IBD and controls. Quantitative immunohistochemistry (n=6). CLM, classical monocyte; CON, control; MFI, mean fluorescence intensity; NCM, non-classical monocyte.
To explore whether this relation of non-classical monocytes and wound healing macrophages might also hold true in the human gut, we analysed the expression of the integrins α4, β7 and αL on macrophages in the colon by immunohistochemistry (figure 5D–F, online supplementary figure 5B,C). αL integrin was clearly upregulated on CD14+ macrophages, which are considered to be proinflammatory in IBD,27 as compared with macrophages expressing the marker CD163.28 On the contrary, β7 and α4 were preferentially expressed on CD163+ cells. Since we had found a similar integrin expression profile on classical and non-classical monocytes, respectively, as detailed above, these data suggested a close relation of classical monocytes with CD14+ inflammatory intestinal macrophages and of non-classical monocytes with CD163+ wound healing intestinal macrophages.
Anti-α4β7 treatment delays intestinal wound healing via a lymphocyte-independent mechanism
As M2 macrophages have been associated with important functions in the proliferative phase of wound healing,17 we next aimed to investigate whether intestinal wound healing is affected by anti-α4β7 integrin treatment in mice. We therefore used a previously established in vivo wound healing assay29 and followed the course of wounds created with a biopsy forceps in the colon of wild-type mice treated with neutralising anti-α4β7 or isotype control antibody.
Indeed, a small, but significant delay in wound healing was observed in anti-α4β7-treated compared with isotype-treated wild-type mice. This resulted in a wound size difference of 20% related to the initial size and almost four times larger remaining wounds in anti-α4β7- versus isotype-treated mice after 7 days (figure 6A). This was confirmed by a series of experiments with β7 integrin-deficient (β7−/−) mice in comparison with wild-type mice, in which wound healing in β7−/− mice was similarly delayed (figure 6C). This was consistent with the idea that deficiency or depletion of α4β7 integrin results in impaired intestinal wound healing.

Delayed wound healing on treatment with anti-α4β7 integrin antibody. (A–C) Endoscopic surveillance of wound healing in C57BL/6 mice treated with anti-α4β7 integrin antibodies or isotype control (A), β7−/− versus wild-type (WT) mice (B) and Rag1−/− mice treated with anti-α4β7 integrin antibodies or isotype control (C). Left panels: representative endoscopic images at the indicated time points. Right panels: quantitative analysis of relative wound diameters over time (n=12–17). (D) Representative H&E staining of wound areas at the indicated time points showing abolished crypt structure and immune cell infiltration. (E) Perilesional expression of CD206 in intestinal wound areas of untreated WT mice over time. Left: representative immunohistochemistry at the indicated time points. Right: quantitative analysis (n=6); scale bars 50 µm (upper panels), 25 µm (lower panels). HPF, high-power field.
Since lymphocytes are considered a major target of anti-α4β7 treatment, we aimed to address whether these effects are accountable to lymphocytes and performed similar assays with Rag1-deficient (Rag1−/−) mice lacking native lymphocytes. However, again, a significant delay in wound healing in anti-α4β7 integrin-treated Rag1−/− mice was observed compared with isotype-treated animals (figure 6B).
Collectively, these findings argued for a lymphocyte-independent impact of anti-α4β7 integrin treatment on intestinal wound healing, and thus supported the hypothesis that wound healing macrophages might be implicated in these effects.
Inhibition of α4β7 in intestinal wound healing leads to decreased perilesional numbers of wound healing macrophages
To analyse a potential role of macrophages in this context, we performed immunohistochemistry on wound areas (figure 6D) at specific time points after wounding.
In first experiments, we characterised the presence of overall macrophages with the pan-macrophage markers F4/80 and CD68, of proinflammatory macrophages with the marker CD86 and of alternatively activated macrophages with the marker CD20614 30 31 on days 1, 3, 5 and 7 after wounding. While both overall macrophage numbers and inflammatory macrophage numbers sharply increased in the early phase of wound healing and peaked on day 3, the abundance of CD206+ wound healing macrophages gradually rose to a peak in the late phase on days 5 and 7 (figure 6E, online supplementary figure 6). This was well in line with current concepts on the contribution of inflammatory macrophages to the early inflammatory phase of wound healing marked by phagocytic and proinflammatory activity and the importance of alternatively activated wound healing macrophages for later phases requiring tissue proliferation and remodelling.17
We therefore investigated the infiltration of macrophages to wound areas from wild-type mice treated with anti-α4β7 or isotype control antibodies on days 1 and day 5. While no relevant differences were noted on day 1, overall F4/80+ macrophage numbers in the perilesional area were numerically and CD206+ wound healing macrophage numbers markedly and significantly reduced in anti-α4β7-treated compared with isotype-treated animals. No such difference was observed for CD86+ inflammatory macrophages. In addition, cells positive for the wound healing macrophage marker CD163 were also decreased (online supplementary figure 7). Of note, no differences in the expression of these markers were observed in unwounded mice treated with or without anti-α4β7 antibodies (online supplementary figure 8). However, consistent differences were observed with wounded tissues from wild type in comparison with β7−/− mice and with sections from isotype-treated versus anti-α4β7 antibody-treated Rag1−/− mice (online supplementary figures 9 and 10). Finally, our findings were confirmed by double staining of day 5 wound areas from wild-type mice treated with anti-α4β7 or isotype control antibodies. These indicated reduced numbers of macrophages expressing CD206, CD163 or the functional wound healing macrophage marker Arginase together with the pan-macrophage markers F4/80 or CD68, whereas the number of iNOS-expressing inflammatory macrophages was unaffected (figure 7).

Reduced perilesional presence of wound healing macrophages under treatment with anti-α4β7 integrin antibody. Perilesional coexpression of F4/80 and CD206 (A), CD68 and CD163 (B), F4/80 and Arg1 (C), and F4/80 and inducible nitric oxide synthase (iNOS) (D) in intestinal wound areas of wild-type mice treated with anti-α4β7 integrin or isotype control antibodies. Tissue was collected 5 days after wounding. Representative (left) and quantitative immunohistochemistry (right, n=9–11); scale bars 50 µm (upper panels), 25 µm (lower panels). HPF, high-power field.

Enrichment of CD16+ monocytes in the peripheral blood of patients with IBD treated with vedolizumab. (A) Schematic depiction of vedolizumab treatment and flow cytometric analyses of monocytes in the peripheral blood. (B) Frequency of CD16+ intermediate and non-classical monocytes and CD16− classical monocytes in the peripheral blood of patients with IBD directly prior to vedolizumab therapy (T1) and after 6 weeks of treatment (T3). Representative (left) and quantitative flow cytometry (right, n=14). (C, D) Frequency of CD14+ (C) and CD163+ (D) cells in the colonic lamina propria of patients with IBD prior to and under treatment with vedolizumab (VDZ). Representative (left) and quantitative immunohistochemistry (right, n=10). Biopsies for immunohistochemistry were obtained after a mean follow-up of 8 months±2.25 months (SEM) after initiation of vedolizumab therapy; scale bars 50 µm (upper panels), 25 µm (lower panels). HPF, high-power field; T, treatment.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Putative model of monocyte subset gut homing and macrophage differentiation. (A) While classical monocytes use ‘traditional’ monocyte homing markers like αLβ2 integrin to home to the gut, non-classical monocytes may also home via α4β7. Once homed to the intestinal tissue, non-classical monocytes preferentially develop to wound healing macrophages and importantly contribute to the late proliferative phase of intestinal wound healing, while inflammatory macrophages are predominantly implicated in the early phase. (B) On anti-α4β7 treatment, non-classical monocyte homing is impaired, leading to reduced intestinal numbers of wound healing macrophages and protracted intestinal wound healing. CLM, classical monocyte; I, inflammatory macrophage; NCM, non-classical monocyte; W, wound healing macrophage.
Taken together, these data suggested that delayed wound healing in α4β7-deficient or -depleted mice is associated with reduced numbers of wound healing macrophages in wounded areas, thus providing a potential mechanism for the macroscopic findings.
Enrichment of non-classical monocytes in the peripheral blood of patients treated with vedolizumab is associated with a decrease in CD163+ intestinal cells
Finally, we attempted to find further evidence for impaired gut homing of α4β7-expressing monocytes in vivo using blood samples from patients undergoing vedolizumab therapy. In view of recent reports indicating that cell populations targeted by anti-α4β7 enrich in the peripheral blood due to impaired gut homing,32 we quantified the abundance of CD16+ monocytes in paired samples of patients undergoing vedolizumab therapy prior to treatment initiation and after 6 weeks of treatment (figure 8A). In the vast majority of patients the frequency of CD16+ non-classical and intermediate monocytes rose leading to a statistically significant increase in this monocyte subset, whereas the frequency of CD16− monocytes was unaffected (figure 8B). Furthermore, we analysed the frequency of CD14+ and CD163+ cells in paired intestinal samples from patients prior to and under treatment with vedolizumab. While CD14+ cell numbers were unaffected by vedolizumab therapy, the number of cells expressing CD163 substantially decreased (figure 8C,D).
Thus, α4β7-dependent gut homing of non-classical monocytes seems to be impaired by vedolizumab leading to a reduction of alternatively activated M2-like macrophages in the gut. These data suggested that the above observations in mice might also be relevant for patients with IBD.
Discussion
In our current study, we show for the first time that α4β7 integrin has an essential function for homing of non-classical monocytes to the gut. In synopsis, our data and the existing literature discussed below support a model, in which classical and non-classical monocytes may use different homing pathways to reach the gut and preferentially differentiate into inflammatory and wound healing macrophages, respectively, in the tissue (figure 9A). This implies that therapeutically interfering with gut homing differentially impacts on monocyte subset homing. In the case of clinically implemented anti-α4β7 integrin antibody therapy, this seems to suppress a non-classical monocyte-wound healing macrophage axis with functional relevance for intestinal wound healing (figure 9B).
The groundwork for our understanding of monocyte homing to peripheral tissues has been laid by research focusing on non-intestinal sites. In particular, studies addressing monocyte homing to atherosclerotic lesions have uncovered several important mechanistic aspects including a crucial role of P-selectin-dependent monocyte rolling, CCR2-mediated cell activation and firm adhesion to intercellular cell adhesion molecule 1.33–35 With regard to gut homing, two independent studies had proposed an important role of CCR9 for monocyte gut homing in the context of IBD.36 37 β7 and α4β7 expression of circulating monocytes had been reported to be negligible,11 36 but these studies had only looked at overall monocytes and not addressed monocyte subsets. Indeed, with regard to the overall monocyte population our data confirm these findings, but add essential further details regarding the distribution of α4β7 on monocyte subsets and indicating some expression on non-classical and intermediate monocytes. Moreover, our data are the first to show direct evidence for a functional effect of vedolizumab on monocytes.
CD14++CD16− classical and CD14+CD16++ non-classical monocytes in humans and Ly6ChighCX3CR1low classical and Ly6ClowCX3CR1high non-classical monocytes in mice have been described as distinct subsets of circulating peripheral blood monocytes in the 1980s and 2003, respectively.21 38 Already at that point, differential migratory behaviour had been associated with these subsets, since classical monocytes were described to preferentially home to inflamed sites in contrast to non-classical monocytes homing to non-inflamed tissues.21 Further research showed that these differences are functionally important for the recruitment of monocytes in atherosclerosis,39 40 but differential roles in homing to the intestine have not been reported so far.
Envisioning the biological consequences of monocyte homing requires consideration of macrophage differentiation as an additional variable. A large body of research indicates that monocytes may develop into macrophages and monocyte-derived dendritic cells (DC) once homed to peripheral tissues.15 Moreover, depending on the presence of cytokines and microbial compounds, such differentiation can give rise to various types of macrophages. In general and although macrophage polarisation is now considered to be more complex18 and partly controversially discussed, an M1-like commitment is considered proinflammatory and leads to macrophages secreting cytokines like TNF-α, IL-12 or IL-23, which are important for the induction of pro-inflammatory T helper cell subsets. Conversely, an M2-like commitment is deemed anti-inflammatory due to production of cytokines like IL-10 or TGF-β.17 Although several subtypes of M2-like macrophages have been described and not all of them perfectly fit into that paradigm,16 an essential role of M2-like macrophages for the maintenance of mucosal homeostasis and tissue remodelling is undisputed.14 Particularly, M2-like macrophages have also been implicated in the process of intestinal wound healing.41 While the initial inflammatory phase after tissue injury is dominated by inflammatory macrophages, which fight potential infectious stimuli by secretion of proinflammatory cytokines, tissue degradation and recruitment of other immune cells, the proliferative phase of wound healing is orchestrated by alternatively activated wound healing macrophages that prevent further tissue damage, secrete growth factors and promote neoangiogenesis.17 26 Since our data show that inflammatory macrophages peak early during intestinal wound healing and wound healing macrophages appear later, this also seems to be the case in intestinal lesions.
Several publications support the view that classical monocytes are the monocyte subset homing to peripheral tissues to differentiate into macrophages, while non-classical monocytes predominantly patrol along vessel walls.14 42 However, already the above-mentioned cutting-edge findings from the study by Geissmann et al suggested that non-classical monocytes indeed home to tissues and play a role in homeostasis.21 This is further promoted by our findings, since we show that latex-bead labelled non-classical monocytes differentiate into macrophages in the tissue and present flow cytometric and microscopic evidence from an in vivo homing model supporting extravasation of non-classical monocytes. Recently, several other reports underscored the notion that non-classical monocytes are skewed towards M2-like differentiation in peripheral tissues following homing. In particular, it has been demonstrated that vessel-patrolling non-classical monocytes rapidly home into wounded tissues and initiate transcription of M2-related genes like mannose receptor (CD206), arginase1 and Fizz1 soon thereafter.25 Moreover, in a skin injury model, Olingy and colleagues showed that non-classical monocytes give rise to wound healing macrophages.24 Yet, it seems important to note that this must be understood as a preference but does not mean that classical monocytes are incapable of developing into wound healing macrophages or non-classical monocytes may not differentiate to inflammatory macrophages as demonstrated by in vitro evidence.43
However, the general perception that non-classical monocytes are biased towards wound healing macrophages fits well with our data indicating that the same preference exists in the intestine and that the inhibition of α4β7-dependent gut homing is associated with delayed wound healing as well as reduced presence of wound healing macrophages in the gut. This strongly argues for a direct axis from α4β7-expressing non-classical monocytes in the peripheral blood to intestinal wound healing macrophages, which can be interrupted by the blockade of α4β7. Although models and research goals were clearly different, this is also well in line with a recent study demonstrating the importance of β7 integrin expression in the innate immune compartment for the development of tolerogenic DCs and macrophage populations in the context of colitis.44
Finally, these aspects culminate in the question on the translational relevance of our findings in the context of IBD therapy with the anti-α4β7 integrin antibody vedolizumab. While the impact of vedolizumab on postoperative course and complications is still controversially discussed,45 46 several reports indicate an increased rate of postoperative infectious surgical site complications in vedolizumab-treated patients undergoing abdominal operation.47–50 Although our punch biopsy wound healing model obviously differs from surgical wound healing in several aspects, our data suggest that such infections might potentially result from slight delays in macrophage-mediated wound healing due to vedolizumab-mediated inhibition of non-classical monocyte homing, finally leading to increased pathogen translocation and subsequent infection of adjacent tissues. Although the observed effects of α4β7 deficiency or depletion on wound healing might be regarded as rather small, it is important to note that they are not substantially different from the effects in other reports addressing the role of key molecules such as epithelial STAT3 in wound healing.51 However, to evaluate the overall biological importance of the phenomenon and to definitely clarify the role of vedolizumab in postoperative wound healing, additional translational and clinical studies in humans in vivo will be required.
In conclusion, we present a previously unknown relevance of α4β7 integrin for non-classical monocyte homing and suggest that non-classical monocytes are biased precursors of wound healing macrophages. This might be clinically relevant in perioperative settings.
Acknowledgments
The research of RA, CN, IA, MFN and SZ was supported by the IZKF and the ELAN programme of the University of Erlangen-Nuremberg, the Fritz-Bender-Stiftung, the Else Kröner-Fresenius-Stiftung, the German Crohn’s and Colitis Foundation (DCCV), the Collaborative Research Unit TRR241 of the German Research Council (DFG), the DFG topic programme on Microbiota, the Emerging Field Initiative and the DFG Collaborative Research Centers 643, 796 and 1181. The present work was performed in fulfilment of the requirements for obtaining the degree ’Dr med' for LS. The authors thank Monique Slawik, Julia Derdau and Dorothee Dziony for excellent technical assistance. Moreover, the authors thank for excellent support by the FACS Core Unit of the University Hospital Erlangen. The 9D8 antibody was provided by F Hoffmann-La Roche.
References
Footnotes
LS and MW contributed equally.
MFN and SZ contributed equally.
Contributors LS, MW, KH, MTB, EB, ASK, AS, LLS and SZ performed the experiments. RA, TR, EB, SFM, KU, LB, MG, AJMW, CN, IA, MFN and SZ provided clinical samples, protocols, reagents or designed experiments. LS, MW, KH, MTB, EB, AS, LLS, CN, IA, MFN and SZ analysed and interpreted the data. LS, MW and SZ drafted the manuscript. All authors critically revised the manuscript for important intellectual content.
Funding Fritz-Bender-Stiftung, Research Grant of the German Research Foundation (ZU 377/3-1), Takeda, scholarship of the Interdisciplinary Center for Clinical Research (IZKF) to LS.
Competing interests MFN has served as an advisor for Pentax, Giuliani, MSD, AbbVie, Janssen, Takeda and Boehringer. MFN and SZ received research support from Takeda, Roche and Shire.
Provenance and peer review Not commissioned; externally peer reviewed.
Correction notice This article has been corrected since it published Online First. The affiliations have been amended.
Patient consent for publication Not required.