Article Text

Abstract
Background Iron replacement therapy is a common treatment in patients with anaemia and Crohn's disease, but oral iron supplements are less tolerated. The pathogenesis of Crohn's disease is attributed to intestinal bacteria and environmental factors that trigger disease in a genetically predisposed host. The aim of this study was to characterise the interrelationship between luminal iron sulfate, systemic iron, the gut microbiota and the development of chronic ileitis in a murine model of Crohn's disease.
Methods Wild type (WT) and heterozygous TNFΔARE/WT mice were fed with an iron sulfate containing or iron sulfate free diet in combination with intraperitoneal control injections or iron injections for 11 weeks.
Results TNFΔARE/WT mice develop severe inflammation of the distal ileum but remained completely healthy when transferred to an iron sulfate free diet, even if iron was systemically repleted. Absence of luminal iron sulfate reduced cellular markers of endoplasmic reticulum (ER) stress responses and pro-apoptotic mechanisms in the ileal epithelium. Phenotype or reactivity of major effector intraepithelial CD8αβ+ T cells were not altered in the absence of luminal iron. Interestingly, ER stress mechanisms sensitised the small intestinal epithelial cell (IEC) line Mode-K to cytotoxic function of effector T cells from TNF∆ARE/WT mice. Pyrosequencing of 16S rRNA tags of the caecal microbiota revealed that depletion of luminal iron sulfate induced significant compositional alterations, while total microbial diversity (Shannon's diversity index) and number of total operational taxonomic units were not affected.
Conclusion This study showed that an iron sulfate free diet in combination with systemic iron repletion prevents the development of chronic ileitis in a murine model of Crohn's disease. Luminal iron may directly affect IEC function or generate a pathological milieu in the intestine that triggers epithelial cell stress-associated apoptosis through changes in microbial homeostasis. These results suggest that oral replacement therapy with iron sulfate may trigger inflammatory processes associated with progression of Crohn's disease-like ileitis.
- Iron
- Crohn's disease
- microbiota
- endoplasmic reticulum stress
- inflammatory bowel disease
- intestinal epithelium
- iron nutrition
- stress
Statistics from Altmetric.com
- Iron
- Crohn's disease
- microbiota
- endoplasmic reticulum stress
- inflammatory bowel disease
- intestinal epithelium
- iron nutrition
- stress
Significance of this study
What is already known about this subject?
In animal models, a high-iron diet enhances intestinal inflammation, and iron induces ROS production and therefore DNA damage.
In human trials, it is suggested that oral iron enhances the inflammation index. Patients with IBD often need iron supplementation.
Endoplasmic reticulum (ER) stress is a possible pathological factor in IBD.
What are the new findings?
Oral iron deprivation prevents chronic ileitis in TNFΔARE/WT mice and systemic iron repletion maintained the protective effect of luminal iron deprivation.
Iron triggers ER stress in intestinal epithelial cells and ER stress sensitises the epithelium towards cytotoxic T cell induced apoptosis.
The comprehensive analysis of the caecal microbiota by pyrosequencing of 16S rRNA tags in WT and TNF∆ARE/WT mice revealed that the depletion of luminal iron sulfate and not inflammation per se induced significant compositional alterations of the caecal microbiota.
How might it impact on clinical practice in the foreseeable future?
These data imply that systemic iron repletion should be preferred for the treatment of anaemic patients with Crohn's disease as orally administered iron sulfate led to intestinal inflammation in susceptible TNFΔARE/WT mice.
Introduction
Present aetiological theories of inflammatory bowel diseases (IBDs) imply a role of environmental factors like smoking, luminal enteric bacteria and trace elements like iron for the pathogenesis in a genetically susceptible host but mechanistic data are lacking.1 One third of patients with IBD develop iron deficiency anaemia due to an inappropriate intake or loss of iron affecting the quality of life. Oral supplementations are common alternatives for the treatment of iron deficiency and contain iron in the form of Fe(II) salts, such as Fe(II) sulfate, Fe(III) polymaltose complexes or haem iron polypeptides. However, oral iron replacement therapy is poorly tolerated and may even contribute to the inflammatory processes and tissue pathology in patients with IBD.2 3
The intestinal epithelium represents a highly selective barrier between the gut lumen and underlying cells of the immune system, and intestinal epithelial cells (IECs) must adapt to constant changes in their environment by processing both bacterial and host-derived immune signals.4 An emerging paradigm suggests that stress responses in the endoplasmic reticulum (ER) contribute to the development of chronic intestinal inflammation through mechanisms that promote the loss of tissue homeostasis including goblet5 and Paneth cell dysfunctions.6 We showed that immune-mediated control mechanisms modulate ER stress-associated activating transcription factor (ATF) 6 signalling at the IEC level. The lack of immune homeostasis triggered ER stress-dependent mechanisms for the development of chronic intestinal inflammation in animal models and human IBD.7 Consistent with these findings, Brandl et al identified an N-ethyl-N-nitrosourea (ENU)-induced mutation that disrupts ATF6-driven ER stress responses leading to increased susceptibility for dextran sodium sulfate (DSS)-induced colitis in mice.8 Most importantly, gene variants of the transcription factor X-box protein-1 (XBP-1) were linked to an increased risk for IBD.6
Heterozygous TNFΔARE/WT mice show impaired regulation of tumour necrosis factor (TNF) synthesis and develop a severe CD8+ T cell-dependent ileitis closely resembling the immune- and tissue-related phenotype of human Crohn's disease with ileal involvement.9 Functional proteome analysis of primary ileal IECs from wild-type (WT) and TNF∆ARE/WT mice fed an iron sulfate containing diet or iron sulfate free diet revealed significant differences in the regulation of proteins participating in energy metabolism and stress responses when comparing inflamed with non-inflamed conditions.10 In this study, we used heterozygous TNF∆ARE/WT mice to investigate the mechanistic role of both luminal iron sulfate and systemic iron in the development of chronic ileitis in a model of Crohn's disease focussing on the effect of iron status on ER stress regulation and changes in gut microbial ecology.
Material and methods
Animals
Conventionally raised WT and heterozygous TNFΔARE/WT mice on a C57BL/6 background received an iron sulfate containing (Altromin C1000, 180 mg Fe/kg as iron sulfate) or iron sulfate free diet (Altromin C1038, <10 mg Fe/kg; N=10/group) for 11 weeks after weaning period (age of 7 weeks). Mice were housed in groups according to gender and treatment. At the age of 18 weeks, mice were killed by cervical dislocation.
An initial dose finding study was performed to evaluate the concentrations for systemic iron replacement with Fe(NO3)3 complexed with nitrilotriacetic acid (NTA) in a 1:2 ratio [FeNTA(1:2)]. The iron sulfate diet contained 180 mg/kg (180 μg/g) iron sulfate and, based on an average daily intake of 720 μg of iron sulfate, the mice were administered an average of 28 μg iron sulfate/g mouse per day. The bioavailability (BV) for iron sulfate is estimated to be about 10% (2.8 μg Fe/g mouse), resulting in an equivalent of 50 nmol [FeNTA(1:2)]/g mouse/day for the systemic iron injection. By applying one injection per week, 10% bioavailability corresponds to 360 μmol (350 μmol+10 μmol as safety value) iron per week. We tested four concentrations of FeNTA for the systemic iron repletion including 360 μmol (10% BV), 180 μmol (5% BV), 90 μmol (2.5% BV) and 45 μmol (1.25% BV). Due to the toxic effects of 360 μmol (10% BV) and 180 μmol (5% BV) FeNTA concentrations, the mice received weekly intraperitoneal injections of 90 μmol FeNTA per g body weight to restore the hepatic non-haem iron concentrations. Control mice on the iron sulfate containing and iron sulfate free diet received NaCl control (Ctrl) injections (0.9% NaCl per g body weight).
For the analysis of tissue pathology, tissue sections of the distal ileum were fixed in 10% neutral buffered formalin. Histopathology was measured by blindly assessing the degree of lamina propria mononuclear cell infiltration, crypt hyperplasia, goblet cell depletion and architectural distortion as previously described.11
Isolation and phenotyping of intraepithelial lymphocytes
See supplemental material
Isolation of primary mouse IECs
Primary IECs from ileal epithelium of WT as well TNF∆ARE/WT mice were purified as previously described.12
Western blot analysis
Purified primary IECs (pooled for each group; equal amount of protein used from each single animal) or Mode-K cells were lysed in 1× Laemmli buffer and 20 μg of protein was subjected to electrophoresis (10% SDS-PAGE gels). Anti-p-RelA (Cell Signaling, Danvers, USA), Anti-grp78 (Sigma Aldrich, Munich, Germany), anti-p-eiF2α (Cell Signaling), anti-cleaved caspase 3 (Cell Signaling), TNF (Cell Signaling), ferritin (FTH1; Cell Signaling) and anti-β-actin (MP Biomedicals, Illkirch, France) were used to detect immunoreactive p-RelA, grp78, p-eiF2α, cleaved caspase 3, ferritin and β-actin respectively, using an enhanced chemiluminescence light-detecting kit (Amersham, Arlington Heights, Illinois, USA).
Non-haem measurement
Livers and ileum were collected and 30 mg tissue was dissolved in acid mix (6 M HCl, 20% v/v TCA). After incubation (65°C, 20 h) and centrifugation (3000 rpm, 3 min), non-haem iron content was measured photometrically using a commercial kit following the manufacturer's instructions (Feren-B, Bioanalytic, Umrich/Freiburg, Germany).
Haematocrit and haemoglobin determination
Blood was drawn from each animal from the vena cava inferior at the killing time point of 18 weeks. Haematocrit and haemoglobin concentrations were measured using the microhaematocrit method (No. 749311; Brand, Wertheim, Germany; Haematocrit-centrifuge 2104; Hettlich, Tuttlingen, Germany) and the cyanmethaemoglobin method (reagent: Bioanalytic 4001; Umrich, Freiburg, Germany; photometer: UV-DK-20; Beckmann, Munich, Germany), respectively. Mean corpuscular haemoglobin concentration (MCHC) values were calculated by using the formula (haemoglobin/haematocrit) × 100.
Isolation and stimulation of CD4+ T cells from spleen
See supplemental material.
Ileal explant cultivation
See supplemental material.
Isolation of mesenteric CD8αβ T cells
See supplemental material.
Epithelial cell and/or intraepithelial lymphocytes (IEL) co-culture experiments
See supplemental material.
Cell-mediated cytotoxicity assay
For quantification of cell-mediated cytotoxicity, the Cellular DNA Fragmentation ELISA (Roche, Mannheim, Germany) was used. The assay was performed according to the manufacturer's standard protocol.
ELISA analysis
TNF, interferon γ (INFγ) or Granzyme B protein concentrations were determined in supernatant of ileal explant cultivation, blood samples or medium from co-culture experiments at different time points, by mouse-specific ELISA kits according to the manufacturer's instructions (R&D Systems, Wiesbaden-Nordenstadt, Germany).
Microbial analysis using 454 pyrosequencing
See supplemental material.
Statistical analysis
Statistical tests were performed using unpaired t-test or one-way ANOVA followed by the Holm–Sidak test. Differences were considered significant if values were <0.05(*) or <0.01(**), <0.001(***).
Results
Chronic ileitis is associated with induction of ER stress and apoptosis in primary ileal IEC
Histological examination (score 0–12) of TNFΔARE/WT mice at the age of 18 weeks revealed severe ileitis in TNFΔARE/WT mice (8.30±0.91) in contrast to healthy WT mice (1.20±0.29) (figure 1A). H&E stained distal ileal segments from severely inflamed TNFΔARE/WT mice show massive leucocyte infiltration, villus atrophy and crypt hyperplasia (figure 1B).

TNFΔARE/WT mice as model for human Crohn's disease-like ileits. Histopathological analysis of WT and TNFΔARE/WT mice (A) with representative H&E staining of ileal segments (B). Alterations in CD8αα+/CD8αβ+ IEL T cell numbers between WT and TNFΔARE/WT mice (C). ER stress-, inflammation and apoptosis-associated protein expression in purified primary ileal epithelial cells from inflamed TNFΔARE/WT mice compared to healthy WT mice (D). ER, endoplasmic reticulum; IEL, intraepithelial lymphocytes; TNF, tumour necrosis factor; WT, wild type.
TNFΔARE/WT mice represent a CD8+ T cell-driven model where disease progression is associated with a loss of CD8αα+ and an increase in CD8αβ+ IEL as major T effector cell phenotype.9 13 We confirmed loss of CD8αα+ and an increase of CD8αβ+ IEL in TNFΔARE/WT compared to WT mice (figure 1C).
Since ER stress in IEC has been discussed in the pathogenesis of IBD, we isolated primary ileal IEC to evaluate ER stress responses in TNFΔARE/WT mice. Five animals from each of the two groups were pooled and 25 μg of total protein was used for western blot analysis. As shown in figure 1D, we detected increased expression levels of the major ER chaperone grp-78 and phosphorylated eukaryotic initiation factor 2α (p-eiF2α) in TNF∆ARE/WT mice compared to WT mice. In addition to ER stress response mechanisms, we detected phosphorylated/activated nuclear factor kappa B (NF-κB) subunit RelA (p-RelA) as well as enhanced protein expression of apoptosis-related cleaved caspase 3 (cc3) in severely inflamed TNF∆ARE/WT mice.
Systemic iron repletion does not extenuate the protective effect of luminal iron sulfate deprivation
We next sought to evaluate the role of luminal iron sulfate on the development of experimental ileitis in TNF∆ARE/WT mice independent from the depletion of systemic iron stores (liver and spleen). Therefore, we fed WT (N=5) or TNF∆ARE/WT (N=8) mice an iron sulfate free diet. In parallel we treated these mice with weekly intraperitoneal injections of FeNTA (iron-repleted) or NaCl (Ctrl injection). The third group of WT and TNFΔARE/WT mice received the iron sulfate containing diet in combination with weekly applied NaCl Ctrl injections. Histological examination of the distal ileum revealed absence of inflammation in TNF∆ARE/WT mice receiving an iron sulfate free diet combined with NaCl Ctrl injections as well as FeNTA-injections (figure 2A). In contrast, TNF∆ARE/WT mice receiving the iron sulfate diet combined with NaCl-Ctrl-injections were severely inflamed. WT groups showed no signs of inflammation. H&E staining of representative paraffin-embedded distal ileal segments from TNFΔARE/WT mice receiving the iron sulfate free diet combined with NaCl Ctrl injections as well as FeNTA injections confirmed the markedly reduced tissue pathology compared to severely inflamed TNFΔARE/WT mice on the iron sulfate containing diet (figure 2C). As shown in figure 2B, iron repletion of WT and TNFΔARE/WT mice on the iron sulfate free diet restored at least 50% of hepatic non-haem iron content compared to level of WT and TNFΔARE/WT mice fed with the iron sulfate containing diet.

Systemic iron repletion did not adversely affect the protective effect of the iron sulfate free diet. Histological analysis of WT and TNFΔARE/WT mice fed with an iron sulfate containing diet or iron sulfate free diet in combination with weekly applied NaCl Ctrl injections or FeNTA injections (A). Non-heme iron content of liver (B). Representative H&E staining of ileal segments (C). Ctrl, control; FeNTA, Fe(NO3)3 complexed with nitrilotriacetic acid; TNF, tumour necrosis factor; WT, wild type.
Haematocrit (Hc) levels of iron-repleted TNFΔARE/WT mice showed no significant differences to control mice (supplementary figure 1A). In contrast, WT and TNFΔARE/WT mice on an iron sulfate free diet showed significantly reduced hepatic non-haem iron content associated with significantly reduced Hc levels compared to all other groups. There were no significant changes in haemoglobin levels between the treatment groups (supplementary figure 1B). Table 1 shows the calculated MCHC values of all groups, demonstrating no significant differences between the different treatments. These results suggested that the iron sulfate free diet almost completely depleted hepatic iron stores without shifting the mice to an anaemic state.
Values of the mean corpuscular haemoglobin concentration
Luminal iron deprivation reduced ER stress and apoptosis in IEC
Inflamed TNFΔARE/WT mice showed enhanced ER stress responses and apoptosis in primary IEC when compared to healthy WT mice (figure 1D). To further elucidate the role of luminal versus systemic iron to trigger ER stress responses and pro-apoptotic signals in the intestinal epithelium, we performed an additional experiment with TNF∆ARE/WT mice receiving an iron sulfate free diet. Consistent with the experiment described above, systemic iron repletion of TNFΔARE/WT mice (1.67±0.20) maintained the protective effect as observed in TNFΔARE/WT mice (1.54±0.25) receiving the iron sulfate free diet and NaCl Ctrl injections in contrast to severely inflamed TNFΔARE/WT mice (7.00±0.63) on the iron sulfate containing diet and NaCl Ctrl injections. Iron repletion of iron sulfate free fed mice (158.61±27.36 μg Fe/g liver) was completely effective to restore hepatic non-haem iron content compared to level of the iron sulfate containing diet (115.55±28.04 μg Fe/g liver). Western blot analysis revealed a slight inhibition of RelA phosphorylation in primary IEC from non-inflamed TNF∆ARE/WT mice receiving an iron sulfate free diet as well as iron-repleted TNF∆ARE/WT compared to inflamed TNF∆ARE/WT mice (figure 3A) receiving the iron sulfate containing diet and NaCl Ctrl injections. However, western blot analysis showed absence of ER stress and apoptosis in primary ileal epithelial cells under luminal iron deprivation (figure 3A, middle row). Interestingly, systemic iron repletion did not affect ER stress levels in IEC, but revealed slightly enhanced protein expression levels for cc3 compared to iron sulfate free fed TNF∆ARE/WT mice with NaCl Crtl injections (figure 3A, right row). To further evaluate IEC iron levels, we performed a ferritin western blot. Consistent with the non-haem iron measurement in total ileal tissue (supplementary figure 2), we detected enhanced levels of ferritin in TNF∆ARE/WT mice fed with the iron sulfate containing diet compared to mice on the iron sulfate free diet. Although the systemic iron repletion led to enhanced expression levels of ferritin compared to NaCl Crtl injected mice, the strongest expression of ferritin in IEC was shown for TNF∆ARE/WT mice receiving an iron sulfate containing diet (figure 3A).
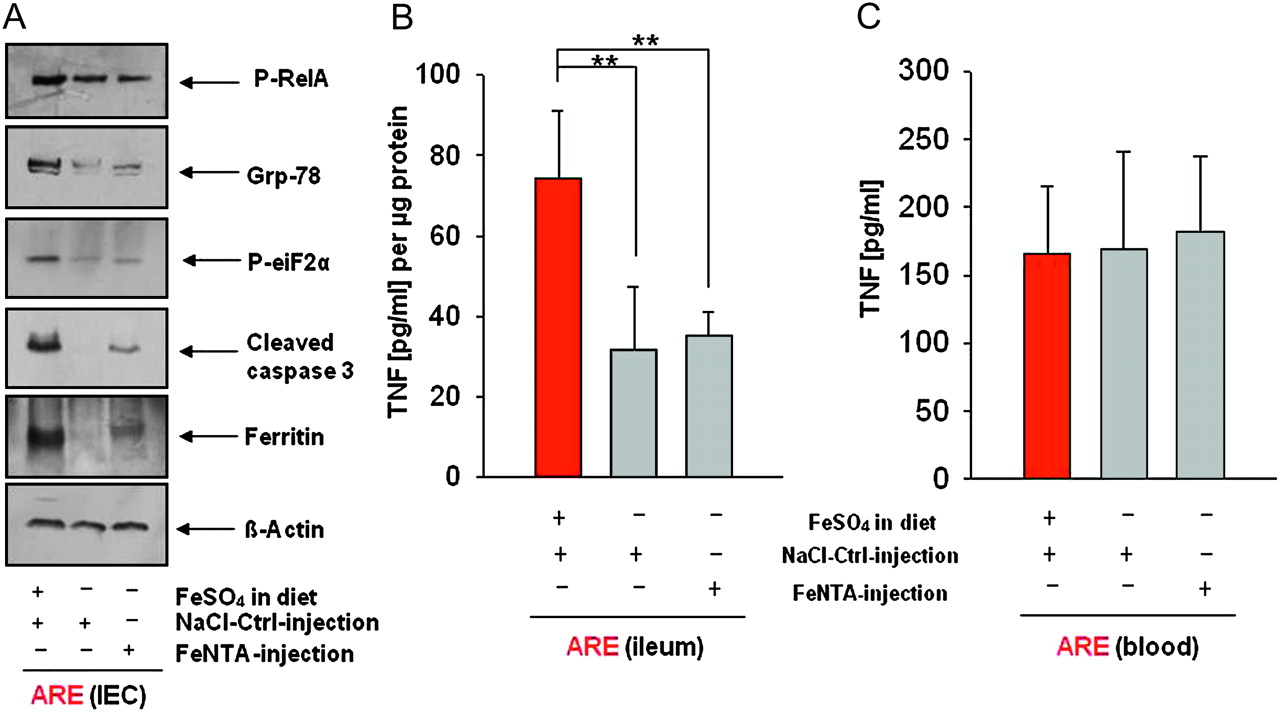
Luminal iron sulfate deprivation as well as systemic iron repletion downregulated ER stress-associated protein expression and apoptosis in primary ileal IECs. Reduced ER stress- and apoptosis-associated protein expression in purified primary ileal epithelial cells from iron sulfate free diet fed (middle row) and iron-repleted (right row) TNFΔARE/WT mice compared to TNFΔARE/WT mice fed with the iron sulfate containing diet (left row) (A). TNF protein expression of cultivated ileal explants (B). TNF protein expression in blood samples at killing time point (C). ER, endoplasmic reticulum; IECs, intestinal epithelial cells; TNF, tumour necrosis factor.
We next cultivated distal ileal segments from TNF∆ARE/WT mice of the second injection experiment for 24 h to determine TNF expression. Consistent with the degree of tissue pathology, ELISA analysis revealed significantly elevated levels of TNF in the inflamed group receiving the iron sulfate containing diet and NaCl Crtl injections (74.10±16.91 pg/ml) compared to the group receiving the iron sulfate free diet and NaCl Ctrl injections (31.58±15.74 pg/ml) as well as FeNTA injections (35.16±6.01 pg/ml) (figure 3B). Blood samples were taken to measure TNF serum concentrations at 18 weeks of age showing no significant differences between the three groups (figure 3C), suggesting that absence of luminal iron sulfate mediates protective effects on the ileal tissue rather than affecting systemic inflammatory responses.
Luminal iron sulfate deprivation and systemic iron repletion did not influence phenotype and activity of IEL
To evaluate the role of luminal iron sulfate deprivation and systemic iron repletion on IEL phenotype, we performed five colour surface staining and FACS analysis. Iron-repleted TNF∆ARE/WT mice showed a marginal, but significant, loss of CD4+ T cells when compared to TNF∆ARE/WT mice on the iron sulfate containing or iron sulfate free diet (figure 4A) with NaCl Ctrl injections. Feeding of an iron sulfate free diet as well as iron repletion in TNFΔARE/WT mice had no significant effect on CD8αα+ or CD8αβ+ T cell numbers when compared to the iron sulfate containing diet, indicating that absence of tissue pathology under luminal iron sulfate deprivation is not due to the loss of effector T cells.

Luminal iron sulfate deprivation as well as systemic iron repletion did not affect IEL phenotype or function. No changes in major effector T cells CD8αα+/CD8αβ+ IEL numbers when comparing inflamed, iron sulfate containing diet fed TNFΔARE/WT mice to iron sulfate free diet fed as well as iron-repleted TNFΔARE/WT mice (A). CD25 and CD69 surface staining in IEL subsets (B and C). No differences in TNF and INF-γ expression after stimulation of CD4+ T cells from spleen of TNFΔARE/WT mice receiving the three different iron protocols (D and E). IEL, intraepithelial lymphocytes; TNF, tumour necrosis factor.
Whereas total T cell numbers in TNF∆ARE/WT mice were not affected, we further elucidated the role of luminal iron sulfate deprivation and systemic iron repletion on T cell activation status by determining CD25 and CD69 expression levels in CD4+, CD8αα+ and CD8αβ+ IEL. As shown in figure 4B, we detected a significant increase in CD25 positive CD8αα+ IEL in TNF∆ARE/WT mice on the iron sulfate containing diet compared to the iron sulfate free diet and iron-repleted TNF∆ARE/WT group. A significant increase in CD25 positive CD4+ IEL was detectable only in the iron sulfate free fed group. In iron-repleted mice, we detected a higher percentage of CD25 positive CD8αβ+ IEL compared to the iron sulfate containing diet fed group. Compared to the iron sulfate containing diet fed group, an increased percentage of CD69 positive CD4+, CD8αα+ and CD8αβ+ IEL was found for the iron sulfate free fed mice as well as iron-repleted TNF∆ARE/WT mice (figure 4C). Interestingly, these results suggest that the T cell activation level was not affected by luminal iron sulfate deprivation.
Finally, T cell reactivity was tested by stimulating isolated CD4+ T cells from the spleen of TNF∆ARE/WT mice receiving the three different iron protocols. As shown in figure 4D,E, no significant differences were detectable for secretion of TNF and INFγ after stimulation (72 h) of CD4+ T cells isolated from each of the TNFΔARE/WT mice fed with the iron sulfate containing diet and the iron sulfate free diet as well as iron-repleted TNFΔARE/WT mice.
Iron triggers ER stress and apoptosis in the small IEC line Mode-K
To further investigate the effect of iron stimulation on IEC cultures, the small IEC line Mode-K was stimulated with 50 μM or 500 μM FeNTA. Western blot analysis showed a concentration-dependent increase for protein expression levels of grp-78 and p-eiF2α in Mode-K cells at early time points (figure 5A). However, induction of pro-apoptotic cc3 appeared for the high concentration at the later time point (3 h), suggesting that sustained ER stress triggers cell death in IEC.

Iron stimulation of IECs led to induction of ER stress and apoptosis and ER stress sensitised IECs towards cytotoxic T cell-induced apoptosis. ER stress- and apoptosis-associated protein expression after iron stimulation in Mode-K cells (A). Higher levels of DNA fragments (lower graph) in ER-stressed Mode-K cells after co-culturing with cytotoxic T cells (Tm + aTC) than in un-stressed Mode-K cells (aTC). Induction of apoptosis is accompanied by cytolytic granzyme B expression (upper graph) (B). aTC, activated T cells; ER, endoplasmic reticulum; IECs, intestinal epithelial cells; Tm, tunicamycin.
ER stress sensitises the epithelium to cytotoxic T cell-induced apoptosis
Since reduction of luminal iron did not affect cytotoxic CD8αβ+ T cell numbers in TNFΔARE/WT mice, we next wanted to evaluate the effect of activated cytotoxic T cells on ER stressed and unstressed IEC. Therefore, we performed co-culture experiments with Mode-K cells and CD8αβ+ T cell isolated from MLN of TNFΔARE/WT mice. Cytotoxic T cells were activated by CD3+CD28+ MicroBeads. Before co-culturing epithelial and T cells, Mode-K cells were pre-incubated for 24 h with BrdU which incorporates into the DNA. After induction of apoptosis, BrdU-labelled DNA fragments can be measured photometrically in co-culture supernatants. In addition, Mode-K cell were treated with the ER stress inducer tunicamycin (Tm; 0.5 μM; 6 h) where indicated.
As shown in figure 5B (lower panel), pre-treatment of Mode-K cells with Tm before co-culturing with activated T cells (Tm+aTC) led to a significant increase in DNA fragmentation compared to the control treatment, suggesting that induction of ER stress in the intestinal epithelium may lead to a higher susceptibility towards cytotoxic T cell-induced apoptosis. Activation of CD8αβ+ T cell was confirmed by measuring the level of cytolytic Granzyme B (GrzmB) (figure 5B upper panel).
Iron sulfate free diet significantly alters gut bacterial composition
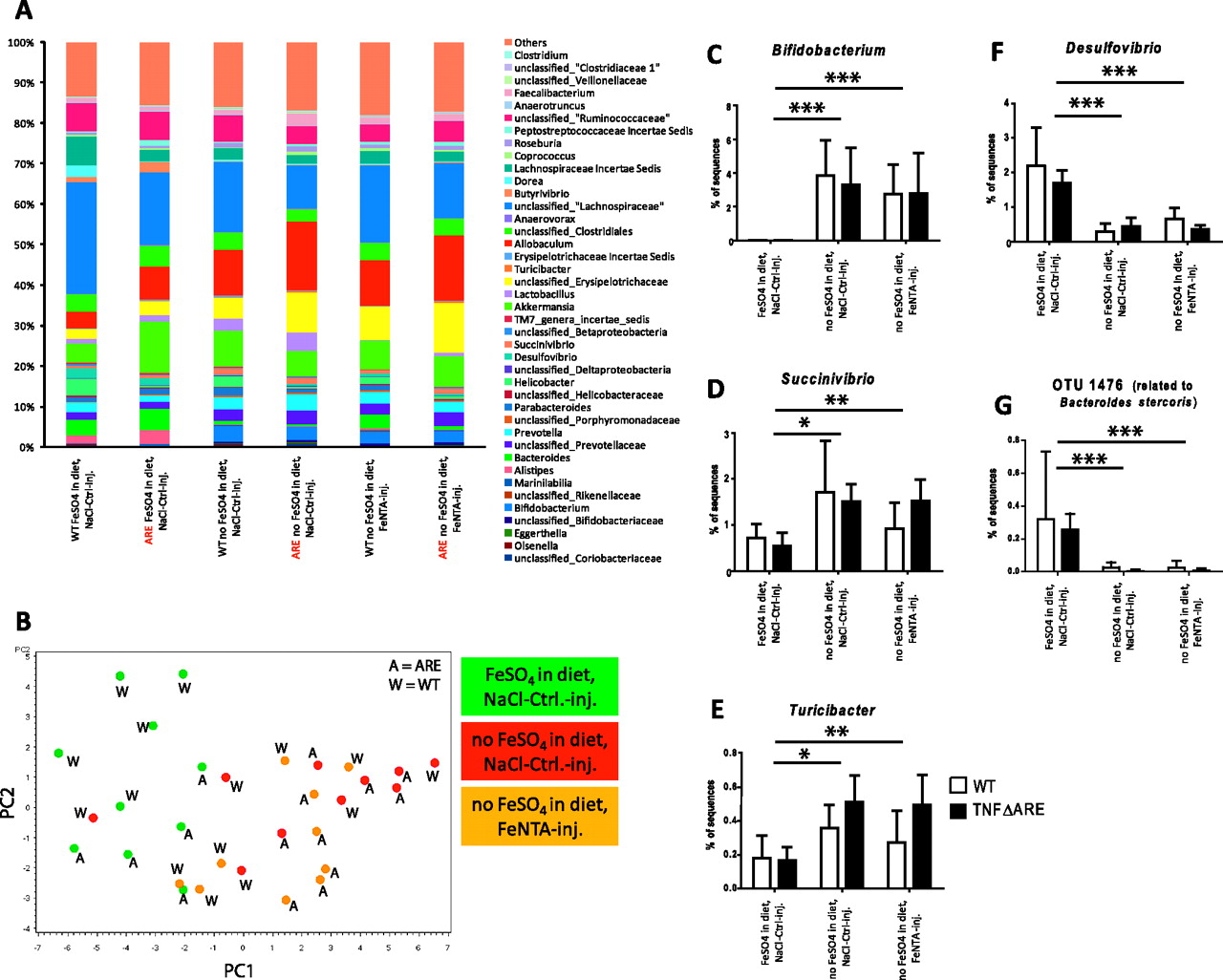
Bacteria likely contribute to the pathogenesis of human IBD and they have been suggested to be one factor that drives ER stress in the intestinal epithelium.14 Therefore, we performed a comprehensive analysis of the caecal microbiota by pyrosequencing of 16S rRNA tags in WT and TNF∆ARE/WT mice from the first injection experiment (N=5 per group). We obtained a total of 132 938 sequences after quality control with an average of 4431 sequences per animal. Analysis of sequence pools revealed significant differences in microbial community composition between groups of animals, while total microbial diversity (Shannon's diversity index) and number of total operational taxonomic units (OTUs) were not significantly affected by the treatments (figure 6A). Figure 6B shows the score plot of the principal component analysis (PCA) analysis based on normalised abundance of dominant bacterial genera with colour code according to iron treatment using principal components PC1 (explained 33% of the variation) and PC2 (explained 11% of the variation). The score plot illustrates major clustering of samples according to iron treatment and not to host genotype, indicating that the iron sulfate free diet led to most of the alterations in microbial population regardless of systemic iron repletion.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Luminal iron sulfate deprivation rather than host inflammatory response induces differences within the gut microbiota. Characterisation of the caecal microbiota at genus level of WT and TNFΔARE/WT mice receiving the three different iron protocols (A). Score plot of the PCA analysis based on proportions of dominant bacterial groups at genus taxonomic level as identified by the RDP classifier, coloured by treatment (B). Proportions of TNFΔARE/WT mice of the genus Bifidobacterium (C), genus Succinivibrio (D), genus Turicibacter (E), genus Desulfovibrio (F) and OTU 1478 (related to Bacteroides stercoris) (G), which all show statistically significant alterations in proportion by luminal iron deprivation (*<0.05; **<0.01; ***<0.001). PCA, principal components analysis; RDP, Ribosomal Database Project; TNF, tumour necrosis factor; WT, wild type.
The analysis revealed that eight bacterial families, nine bacterial genera (supplementary table 1), and 46 OTUs (supplementary table 2) were significantly (p<0.01) affected by luminal iron sulfate deprivation. For most taxa, intraperitoneal injection of iron had no detectable effect. Among others, the genera Bifidobacterium (p=0.0018) (figure 6C), Succinivibrio (p=0.0027) (figure 6D), Turicibacter (p=0.0020) (figure 6E) and Clostridium (p=0.0017) were significantly increased in mice fed with the iron sulfate free diet, while the genera Desulfovibrio (p<0.0001) (figure 6F) and Bacteroides related OTU 1476 were signifcantly decreased. The indicated changes in bacterial composition significantly correlated with disease activity (data not shown).
Only one phylum, two families, two genera (supplementary table 3), and 29 OTUs (supplementary table 4) showed significant differences (p<0.01) in abundance in WT and TNF∆ARE/WT mice. Among others, TNF∆ARE/WT mice had an overall decrease in the phylum Proteobacteria (p=0.0024) and the genus Helicobacter (p=0.0016) and a significant increase in Peptostreptococcaceae Insertae Sedis (p=0.0025).
One genus, Dorea, (supplementary table 5), and 13 OTUs (supplementary table 6) were affected by an interaction between treatment and host-mediated inflammation.
Discussion
In the present study, we investigated the role of iron sulfate as an environmental factor in the pathogenesis of chronic ileitis in the TNF∆ARE/WT mouse model. We clearly demonstrated that a diet free of iron sulfate prevents the onset of severe ileitis. Most importantly, iron repletion by weekly injections maintained the protective effect of the iron sulfate free diet, suggesting a critical role for luminal but not systemic iron in the pathogenesis of chronic ileitis in the genetically susceptible host. The iron sulfate free diet as well as the iron repletion had no impact on the percentage and activation level of disease-relevant IEL effector T cells (CD8αβ+), but interestingly, luminal iron depletion led to a significant reduction of ER stress in primary ileal epithelial cells. Consistently, iron exposure to cultured epithelial cells triggered ER stress responses, and as a consequence, co-culture experiments revealed a higher susceptibility of ER-stressed IEC towards cytotoxic T cell-mediated apoptosis. These results clearly suggest that reduction of ER stress could be one of the main mechanisms by which a decrease in luminal iron exposure reduces inflammation. Recent studies indicate that ER stress responses at the epithelial cell level play an important role in the pathogenesis of IBD.(reviewed by Kaser and Blumberg15) In the present study, the iron sulfate free diet as well as iron repletion inhibited activation of ER stress responses and pro-apoptotic processes, suggesting that reduction of luminal iron sulfate may prevent tissue pathology in TNF∆ARE/WT mice through mechanisms that involve ER stress-related apoptosis in the epithelium.
Iron supplementation may become ultimately necessary for the treatment of anaemia as one of the most frequent complications in IBD. We speculate that an oral iron supplementation strategy may lead to an exacerbation of intestinal inflammation. There is accumulating evidence in patients with chronic inflammation that oral iron supplementation may contribute to inflammatory processes and tissue pathology because of the pro-oxidative capacity of iron. Inflamed mucosa from patients with IBD already show an imbalance in ROS production and antioxidative defence mechanisms.16 Increased reactive oxygen intermediates (ROIs), DNA oxidation products as well as elevated iron levels in combination with decreased copper and zinc superoxide dismutase (Cu–Zn SOD) activity were found in mucosal biopsies from patients with Crohn's disease. Elevated mucosal iron levels were also detected in patients with ulcerative colitis.17 Patients with IBD suffer from an increase in mucosal ROS production, measured by luminal-amplified chemiluminescence. For ulcerative colitis, this increase is in proportion to disease activity.18 Application of an iron chelator on mucosal ulcerative colitis biopsies significantly decreased luminal-amplified chemiluminescence pointing to a possible therapeutic use.19 Erichsen et al showed that oral supplementation with ferrous fumarate for patients with Crohn's disease led to an increasing trend of the disease activity index accompanied by diarrhoea, abdominal pain and nausea. Additionally, patients had significantly decreased plasma-reduced cysteine as well as plasma-reduced glutathione as possible markers of oxidative stress.20
It has also been shown that the gut microbiota differs between healthy humans and patients with IBD,21 and bacterial translocation and increased microbial colonisation of the mucosal surface is implemented in the pathogenesis of Crohn's disease.22 23 Bacteria might drive oxidative and ER stress as well as apoptosis in the epithelium, and iron deprivation might induce both compositional and functional changes within the gut microbiota. Accordingly, our study revealed major alterations in microbial composition by the iron sulfate free diet, while host genotype, inflammatory response, and systemic iron application had a smaller effect. Interestingly, Desulfovibrio spp., which decreased through the iron sulfate free diet, produce toxic sulfides, which have been suggested to contribute to pathogenesis in IBD, and which have been shown to be increased in patients with ulcerative colitis.24 Bacteroides, Bifidobacterium, Prevotella and Clostidium are also often discussed in context of IBD.25 An iron sulfate free diet induced a significant increase in the abundance of bifidobacteria, which are often used as probiotics and have been shown to prevent inflammation in murine models of IBD.26
In conclusion, this study clearly revealed that the absence of luminal iron sulfate inhibits the development of Crohn's disease-like ileitis in TNF∆ARE/WT mice. We clearly demonstrated that deprivation of luminal iron sulfate led to a significant change in microbial composition independently of systemic iron repletion. In parallel, luminal iron sulfate deprivation was accompanied by markedly reduced ER stress and apoptosis in primary epithelial cells. At the level of T cell activation, luminal iron sulfate deprivation as well as systemic iron repletion did not influence percentage of IEL subsets or T cell activity in TNF∆ARE/WT mice, supporting the hypothesis that the luminal milieu including presence of iron sulfate and/or changes in the bacterial ecosystem may shape susceptibility of the epithelial cell interface towards ER stress-related and T cell-mediated apoptosis. However, and most importantly, these data imply that systemic iron repletion in contrast to oral iron supplementation with iron sulfate should be preferred in the treatment of patients with anaemia and Crohn's disease. Since non-haem iron sulfate has profound pro-inflammatory effects in an animal model for Crohn's disease-like ileitis, the role of dietary iron including haem iron as a disease-conditioning environmental factor should be evaluated in further experiments.
Acknowledgments
The authors are particularly grateful to Dr Georg Kollias at the Biomedical Sciences Research Center “Al. Fleming” (Vari, Greece) for providing the TNFΔARE/WT mice. We thank the staff of the Core for Applied Genomics and Ecology (CAGE) at the University of Nebraska for excellent support in the sequencing of 16S rRNA tags.
References
Supplementary materials
Online only appendix
Files in this Data Supplement:
Footnotes
See Commentary, p 287
Linked articles 229047.
Funding German Research Foundation (DFG) GRK 1482.
Competing interests None.
Ethics approval The animal-use protocols were approved by the Bavarian Animal Care and Use Committee (AZ 55.2-1-54-2531-74-06).
Provenance and peer review Not commissioned; externally peer reviewed.