Article Text

Abstract
Objective Acinar cells display plasticity in vitro and in vivo and can activate a variety of differentiation programmes that may contribute to pancreatic diseases. The aims were to determine: (1) the differentiation potential of acinar cells under conditions which favour stem cell survival, and (2) its relationship to the phenotypes acquired by pancreatic epithelial cells in chronic pancreatitis.
Design Murine acinar cells were cultured in suspension and their molecular phenotype was characterised by qRT-PCR, chromatin immunoprecipitation, immunocytochemistry and global transcriptome analysis. These findings were compared to the changes occurring in experimental chronic pancreatitis induced by pancreatic duct ligation and chronic caerulein administration.
Results Acinar cells in suspension culture acquired a dedifferentiated phenotype characteristic of pancreatic embryonic progenitors, consisting of the co-expression of Ptf1a and Pdx1, presence of an embryonic-type PTF1 transcriptional complex, activation of the Notch pathway, and expression of additional pancreatic progenitor cell markers such as CpA1, Sox9 and Hnf1b. A senescence programme, associated with activation of Ras and ERK signalling, limited the proliferative capacity of the cells. A similar progenitor-like phenotype with activation of a senescence programme was observed in experimental chronic pancreatitis induced by pancreatic duct ligation or repeated caerulein administration, with the concomitant and differential activation of proliferation and senescence in distinct cell populations.
Conclusions Acinar cells dedifferentiate into an embryonic progenitor-like phenotype upon suspension culture. This is associated with the activation of a senescence programme. Both processes take place in experimental chronic pancreatitis where senescence may contribute to limit tumour progression.
- Acinar cells
- pancreatic progenitors
- dedifferentiation
- senescence
- chronic pancreatitis
- pancreatic ductal adenocarcinoma
- pancreatic cancer
- pancreatic damage
- pancreatic enzymes
- pancreatitis
- stem cells
Statistics from Altmetric.com
- Acinar cells
- pancreatic progenitors
- dedifferentiation
- senescence
- chronic pancreatitis
- pancreatic ductal adenocarcinoma
- pancreatic cancer
- pancreatic damage
- pancreatic enzymes
- pancreatitis
- stem cells
Significance of this study
What is already known about this subject?
Acinar cells can differentiate into ductal, endocrine, adipocytic and hepatic cells. They have been identified as major contributors to the pathogenesis of chronic pancreatitis (CP) and may be the origin of pancreatic ductal adenocarcinoma (PDAC).
A history of CP is associated with an increased risk of PDAC in humans; CP enhances PDAC development in genetic mouse models.
What are the new findings?
Murine acinar cells can dedifferentiate and acquire a pancreatic progenitor-like phenotype.
Adhesion cues suppress the acquisition of the progenitor cell phenotype.
Dedifferentiated cells activate a senescence programme with concomitant activation of wild-type Ras and ERK, thus limiting their proliferative potential.
The same mechanism is activated in experimental CP caused by pancreatic duct ligation or repeated caerulein administration, where it can protect against tumour development.
How might it impact on clinical practice in the foreseeable future?
Our study has no direct impact on clinical practice. However, we show that the activation of an embryonic phenotype and of a senescence response may participate in the physiopathology of CP and PDAC. This knowledge may help to design strategies to prevent the development or progression of these conditions.
Introduction
The exocrine pancreas comprises >90% of acinar cells producing digestive enzymes and ductal cells secreting electrolytes and mucins. Understanding the molecular mechanisms involved in the regulation of exocrine pancreatic differentiation is crucial to unravel the pathogenesis of diseases such as chronic pancreatitis (CP) and pancreatic ductal adenocarcinoma (PDAC), to identify the mechanisms through which CP is associated with an increased risk of PDAC, and to develop novel therapeutic strategies.1 2
PDAC is one of the most lethal human tumours, with a 5-year survival rate of <5%.1 PDAC has long been thought to originate from ductal cells based upon tumour histology and the presence of ductal preneoplastic lesions (pancreatic intraepithelial neoplasia (PanIN)). However, several recent studies using lineage tracing in mice have shown that both PanINs and PDAC can arise from acinar cells3–6 upon introduction of a mutant Kras2 knock in allele. Indeed, so far it has not been demonstrated that targeting mutant Kras2 to ducts leads to PDAC. Mutant Kras is a much more effective oncogene when activated in the embryo than in adult mice, where chronic administration of caerulein leads to CP and enhances PDAC development.3 It has been proposed also that transgene-mediated induction of levels of Ras signalling above a certain threshold may suffice for CP and PDAC.5 Upon Ras activation, acinar cells have been reported to undergo ductal differentiation, a process referred as acinar–ductal metaplasia that may resemble an embryonic phenotype.7 These findings suggest that CP-associated ductal metaplasia constitutes an intermediate step towards PDAC.
A detailed understanding of the molecular mechanisms involved in acinar–ductal metaplasia is, however, currently lacking.8 The precise mechanisms through which deregulation of acinar differentiation takes place upon development of PDAC are not known. The pancreatic transcription factors Ptf1a and Pdx1 are differentially expressed in acinar and endocrine cells in the adult. In the embryo, they are required for organ formation and are uniquely co-expressed in pancreatic progenitors.8 9 During early development, Ptf1a binds Rbpj to form the embryonic PTF1 complex (ePTF1). ePTF1 binds to the promoters of Ptf1a and Rbpj-like (Rbpjl), leading to a super-induction of Ptf1a and to the expression of Rbpjl.10 11 Subsequently, Rbpjl replaces Rbpj as the Ptf1a partner and the adult-type PTF1 (aPTF1) complex drives high levels of expression of transcripts coding for digestive enzymes, supports Ptf1a and Rbpjl expression through an autoregulatory loop, and maintains acinar homeostasis.11 12 The co-activator Pcaf associates with Ptf1a and ensures high transcriptional activity and nuclear localisation.13
Cell proliferation can be regulated through mechanisms linked to induction of differentiation; for example, Ptf1a couples acinar cell differentiation and G1–S progression.14 In addition, various types of stresses including aberrant oncogenic signalling, DNA damage, telomere dysfunction, and oxidative stress can lead to senescence, characterised by permanent growth arrest.15 Oncogene-induced senescence can occur in early stages of tumour development both in mouse cancer models and in patient tissues and is thought to play an important role as a tumour suppressor.15
Previous work revealed that human and rodent acinar cells display potential for plasticity16–19 and that, upon adherence, they acquire a ductal phenotype. Here, we conduct a detailed quantitative analysis of the molecular changes associated with loss of murine acinar differentiation. Because prevention from adherence has been established as a useful method to isolate stem cell/progenitor populations, acinar cells were cultured in suspension. We show that, under these conditions, acinar cells do not attain a ductal phenotype but rather dedifferentiate, acquiring features consistent with pancreatic progenitor cells. In these cells a senescence programme is activated, possibly initiated by oncogenic stress due to activated Ras signalling. Using two models of experimental CP induced by pancreatic duct ligation (PDL) or repeated caerulein administration,20 21 we show that the embryonic progenitor cell phenotype is present in these lesions with the concomitant activation of senescence associated with the restricted proliferation of a subpopulation of cells.
Materials and methods
For a detailed description, see Supplementary Materials and Methods.
Cell isolation and culture
Acinar cell fractions from CD1 mice (age 8–12 weeks) were obtained as described.22 Cells were cultured in suspension or in monolayer in RPMI 1640 glutamax medium (Gibco BRL, Paisley, Scotland) supplemented with 10% fetal bovine serum, penicillin, streptomycin, geneticin sulfate, and soybean trypsin inhibitor.
Pancreatic duct ligation
A laparatomy was performed in 8–10-week-old CD1 mice through a midline abdominal incision. The pancreatic ducts of the gastric and the splenic segments were ligated as described.20 A double ligature was performed around the pancreas, just next to the blood vessels, in a region close to the great curvature of the stomach marked by the cranial mesenteric lymph nodes. The ligation totally obstructs exocrine flow from the gastrosplenic portion of the pancreas, which accounts for 50–60% of the entire pancreas.
Caerulein-induced chronic pancreatitis (CaeCP)
Chronic damage was induced in 8-week-old C57BL/6 mice by administering 5 h intraperitoneal injections of caerulein (50 μg/kg; Sigma, Bornem, Belgium) 3 days/week over 4 weeks.21 Control mice received saline.
Results
Acinar cells dedifferentiate when cultured in suspension
A preparation of 92% acinar cells, determined by amylase immunostaining, was obtained (referred to as ‘isolated acini’) (supplementary figure 1). Cells could be maintained as free-floating aggregates (suspension cultures) in serum-supplemented medium (supplementary figure 2B) for up to 10 days.
Gene expression analysis was performed after 24 h and 5 days (D5) of culture (figure 1). Within 24 h, acinar cells underwent dedifferentiation based on qRT-PCR and immunocytochemistry analyses. This is reflected by a dramatic downregulation of digestive enzymes and the transcriptional regulators involved in acinar cell maturation. Regarding the PTF1 complex, after 24 h, Ptf1a and Rbpjl mRNA levels were reduced to 11±3% and 1±1% of their initial expression levels. Pcaf and Mist1 transcript levels were also markedly reduced (18±2% and 1±1% of isolated cells, respectively) (figure 1A). By contrast, there was only a modest reduction in Rbpj transcript levels (36±4%). Accordingly, with the loss of acinar transcription factors, there was a dramatic drop in the levels of transcripts encoding for digestive enzymes with carboxypeptidase A1 (Cpa1), chymotrypsinogen B1 (Ctrb1), amylase 2 (Amy2), and elastase 1 (Ela1) all being <10% of the original levels (figure 1B). In comparison with 24 h of culture, a significant upregulation in Ptf1a transcript levels was observed at D5 (34±7% of isolated cells, p=0.001) whereas the levels of transcripts coding for Rbpjl, Rbpj, Pcaf, and Mist1 remained unchanged (figure 1A). At D5, Cpa1 and Ctrb1 transcript levels remained very low (1.6±0.3% and 1.3±0.3%) but significantly higher than those of Amy2 and Ela1 that were essentially undetectable (0.1%) (figure 1B). Interestingly, Cpa1 and Ctrb1 are markers of pancreatic multipotent progenitors and early acinar differentiation, respectively.23 24 Protein expression was assessed by immunocytochemistry: Cpa and Ctrb were detected in most cells while Amy expression was essentially undetectable at D5 (figure 1D).
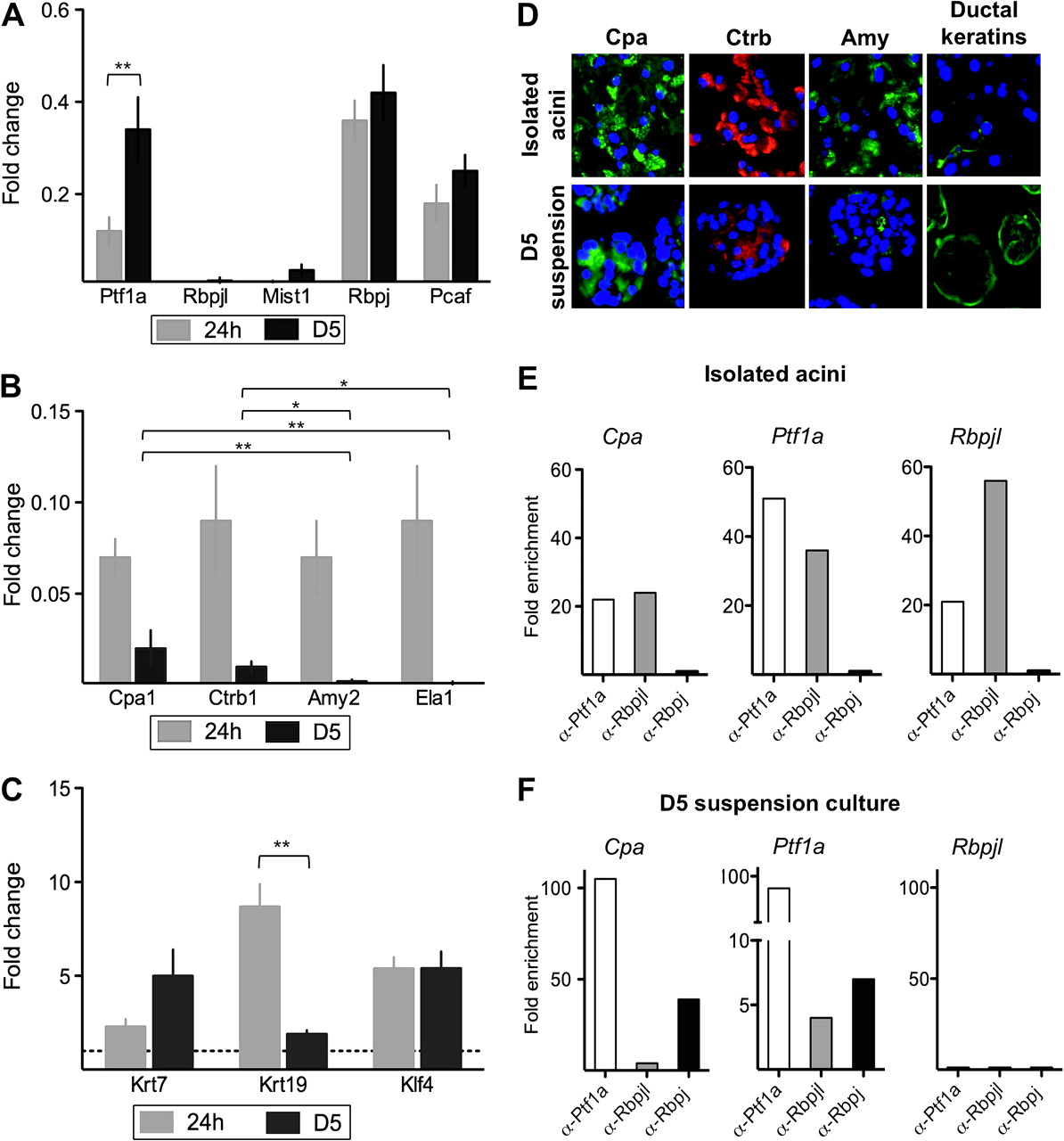
Expression analyses of acinar and ductal markers, and ChIP of promoter occupancy by PTF1 complex components, in freshly isolated acinar cells and in D5 suspension cultures. (A–C) qRT-PCR analysis at 24 h (grey) or 5 days (black), relative to isolated acini, referred to Hprt (mean±SEM; n=4–12). *p<0.05; **p<0.01. (D) Immunofluorescence analysis of digestive enzymes and ductal keratins. Nuclei are stained with DAPI. *p<0.05; **p<0.01. (E,F) ChIP-qPCR analysis of Cpa1 (left), Ptf1a (middle), and Rbpjl (right) promoter occupancy with antibodies specific for Ptf1a, Rbpjl, or Rbpj. Results are expressed as fold enrichment over controls. One representative experiment is shown (n=3–4).
Concomitant to the extinction of the acinar programme, an induction of markers shared by embryonic and adult ducts was observed. This was characterised by the upregulation of Krt7 (2.3±0.4-fold at 24 h and 5.0±1.4-fold at D5) and Krt19 (8.7±1.2-fold at 24 h and 1.9±0.2-fold at D5). mRNA levels of Klf4, a transcriptional activator of Krt19,25 were induced both at 24 h and at D5 (5.4±0.6-fold at 24 h, 5.4±0.9-fold, respectively) (figure 1C,D).
To trace the origin of cells expressing ductal markers (ie, Krt19), ElaCreERT2;R26R-LSL-LacZ mice were pretreated with tamoxifen. Only acinar cells were labelled under these conditions. Isolated acini showed a minimal contamination by Krt19-expressing cells and a high level of Cre-mediated recombination. A clear-cut increase in LacZ-positive Krt19-expressing cells was observed upon suspension culture, indicating that keratin-positive cells originate from acinar cells (supplementary figure 2).
Dedifferentiated suspension cultures display an embryonic-type PTF1 complex
The low levels of Ptf1a – together with the sustained expression of Rbpj, Cpa1, and Ctrb1 – and the lack of Rbpjl suggest that cells in suspension cultures display features of embryonic progenitors. To assess the presence of an ePTF1 complex, we analysed promoter occupancy using ChIP with antibodies specific for either Rbpjl or Rbpj (supplementary figure 3).
In agreement with the reported composition of the aPTF1 complex, Ptf1a and Rbpjl – but not Rbpj – were strongly enriched at the Cpa1 promoter in isolated acini from adult pancreas (figure 1E, left). By contrast, Ptf1a and Rbpj were strongly enriched at the Cpa1 promoter in suspension cultures (figure 1F, left). As expected, Rbpjl was not found at the Cpa1 promoter in suspension cultures since it is not expressed in these cells (figure 1A,F). Ptf1a was modestly enriched at the Ela1 promoter in D5 cultures, where neither Rpbj nor Rbpjl were detected (data not shown), in agreement with the almost undetectable expression of Ela1 mRNA in suspension cultures (figure 1B).
The ePTF1 complex, containing Rbpj, activates expression of Ptf1a and Rbpjl. Accordingly, Ptf1a and Rbpjl were strongly enriched at the Ptf1a and Rbpjl promoters in freshly isolated acini (figure 1E, middle and right panel). By contrast, Ptf1a and Rbpj were strongly enriched at the Ptf1a promoter – but not at the Rbpjl promoter – in suspension cultures (figure 1F, middle and right panel). These results are consistent with the transcript levels of Ptf1a and Rbpjl (figure 1A) and with the notion that suspension cultures display an embryonic-like transcriptional programme.8 23 26
Acinar suspension cultures display features of pancreatic embryonic progenitors
We analysed the expression of two categories of stem cell markers.
Established pancreatic progenitor markers
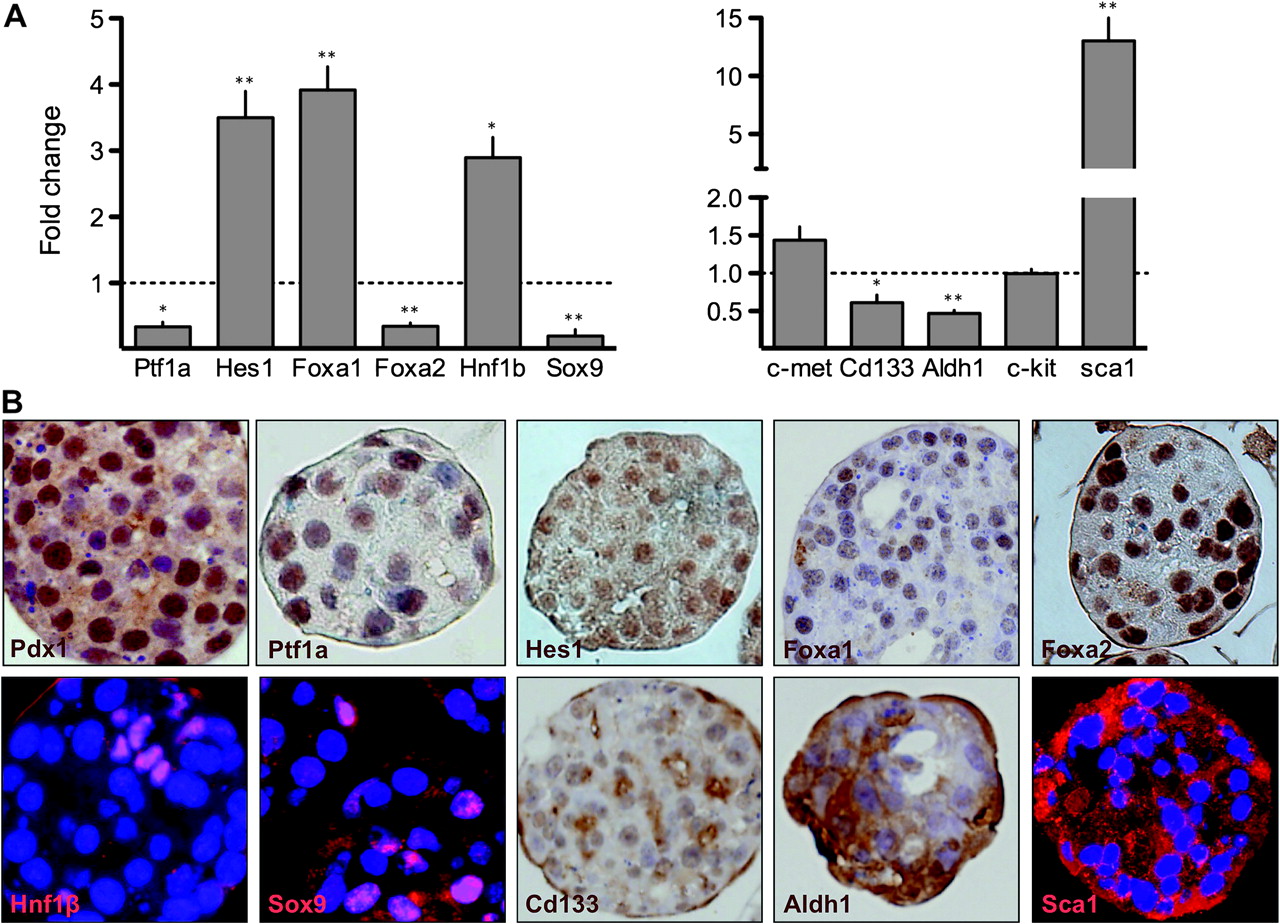
Well established pancreatic progenitor markers that are essential for embryonic organ development, include Pdx1, Ptf1a, and the Notch target gene Hes1. All three proteins were detected in the majority of cells in D5 cultures (figure 2B). Ptf1a mRNA is expressed in D5 suspension cells. Hes1 mRNA levels were 3.5±0.4-fold higher in D5 suspension cultures (figure 2A).

D5 suspension cultures display characteristics of embryonic pancreatic progenitors. (A) qRT-PCR analysis of pancreatic progenitor and stem cell marker transcripts, relative to isolated acini, referred to Hprt. Mean±SEM (n=8–12) (*p<0.05;**p<0.01). (B) Immunocytochemical and immunofluorescence analysis of pancreatic progenitor and stem cell markers in D5 suspension cultures. Nuclei are counterstained with haematoxylin or DAPI.
Regarding other progenitor markers, Foxa1 mRNA levels increased 3.9±0.4-fold during culture whereas Foxa2 mRNA was downregulated (0.4±0.1-fold) (figure 2A); both proteins were detected in suspension cultures (figure 2B). Nkx6.1 mRNA and protein were both undetectable (data not shown). Hnf1b and Sox9 are expressed early on during pancreas development.8 Hnf1b mRNA levels were 2.9±0.3-fold higher at D5 (figure 2A); Hnf1b was detected only in approximately 10% of cells. (figure 2B). As of Sox9, we observed a decreased mRNA expression during suspension culture (0.2±0.1) (figure 2A) and the protein was detected in the nucleus of approximately 15% cells (figure 2B), predominantly co-localising with Hnf1b (data not shown).
Markers reported to label stem cell populations
Markers reported to label a stem cell population in the normal adult pancreas, include c-Met, Cd133, c-Kit, Aldh1, and Sca127–29: c-Met and c-kit mRNA expression was unchanged whereas Aldh1 and Cd133 mRNAs were decreased (0.6±0.1 and 0.5±0.1, respectively) (figure 2A). Cd133 was detected by immunohistochemistry in 40–50% of cells (figure 2B) and Aldh1a was expressed in >90% cells (figure 2B) in D5 cultures. Sca1 mRNA was upregulated 13.0±3.4-fold (figure 2A) and the protein was detected using immunofluorescence (figure 2B).
These results further support the notion that acinar cells cultured in suspension acquire features of embryonic pancreatic progenitors (supplementary figure 4).
Adherence suppresses the embryonic-like expression profile of the suspension-cultured acinar cells
Freshly isolated acini showed poor adherence to culture-treated dishes. However, a fraction of cells readily adhered after 3–4 days of culture in suspension. The proportion of acinar-derived cells populating the monolayers was comparable to their contribution to the suspension cultures, assessed using ElaCreERT2;R26R-LSL-LacZ mice (supplementray figure 2). In the monolayers, Ptf1a and Pcaf mRNA expression was highly reduced compared to the suspension cultures (0.34±0.07 vs 0.04±0.03-fold and 0.25±0.04 vs 0.03±0.01-fold, respectively). Rbpjl mRNA was undetectable in both conditions (figure 3A). By contrast, Rbpj mRNA levels were unchanged (figure 3A). In agreement with the lack of expression of the pancreas-specific components of PTF1, Cpa1 and Ctrb1 mRNAs were essentially undetectable under these conditions (60- and 13-fold lower than in suspension cultures, respectively), as were Amy2 and Ela1 transcripts (figure 3B). Krt19 and Krt7 mRNA expression was similar in monolayer and suspension despite reduced Klf4 expression (figure 3C).
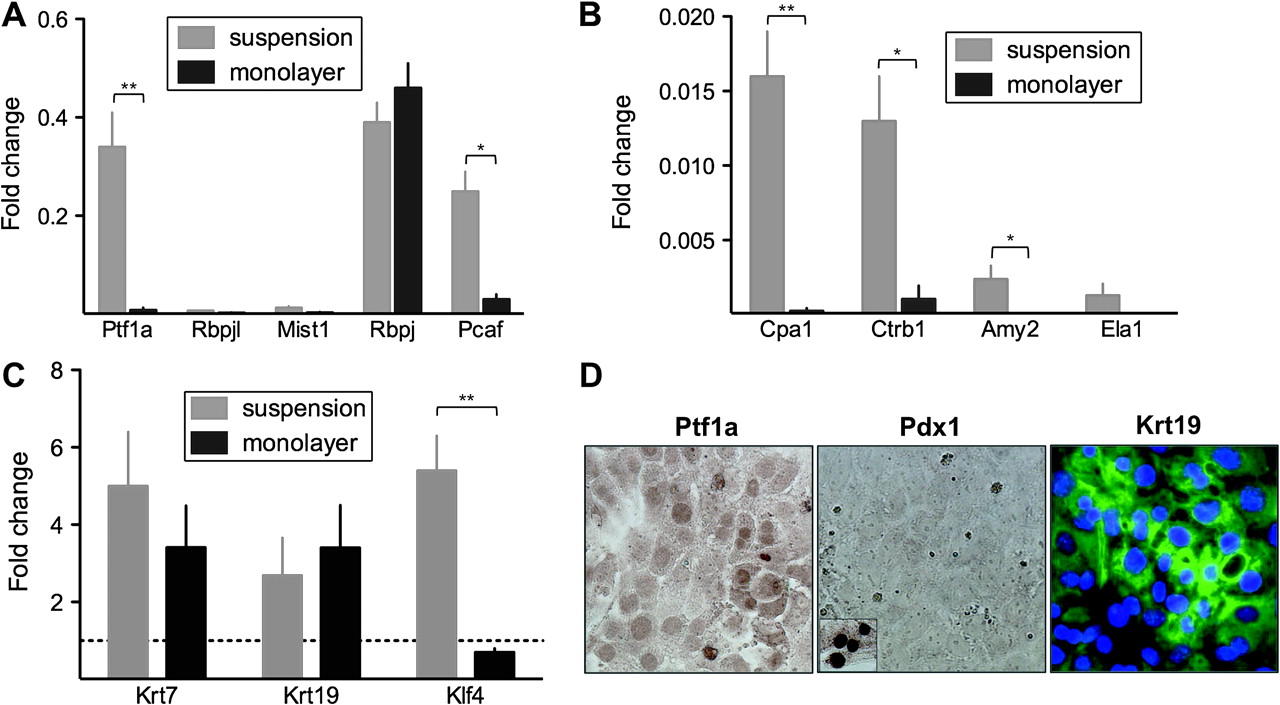
Expression analyses of acinar and ductal markers in acinar suspension cultures and monolayer culture. (A–C) qRT-PCR analysis of transcript levels in suspension cultured cells (grey bars) versus monolayer cultures (black bars), relative to isolated acini and referred to Hprt. Mean±SEM (n=3–4). *p<0.05; **p<0.01 (D) Immunocytochemical analysis of Ptf1a, Pdx1 and Krt19 in adherent cultured cells.
The loss of Ptf1a expression in Krt19-expressing cells was confirmed using immunocytochemistry and western blotting; Pdx1 was also undetectable in adherent cultures (figure 3D and supplementary figure 5). The lack of Ptf1a, Pdx1 and other early pancreatic markers indicates that the embryonic progenitor phenotype is suppressed upon adherence.
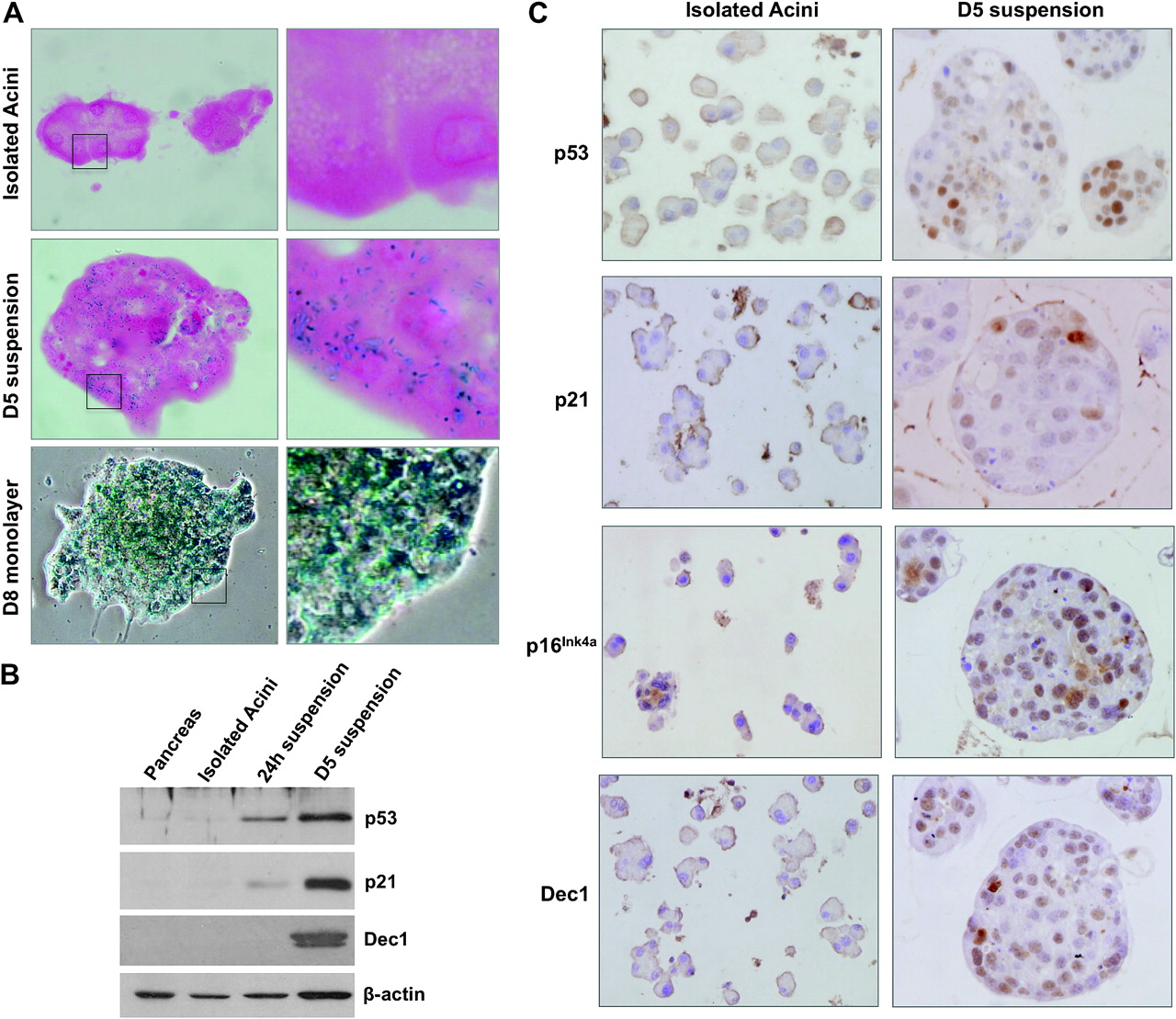
Dedifferentiated acini undergo a senescent-like permanent growth arrest
Despite the acquisition of characteristics of pancreatic progenitors, suspension cultures lack the ability to proliferate (supplementary figure 6A). To assess apoptosis, activated caspase 3 was analysed and only a low proportion of cells were positive; using western blotting, full-length caspase 3 was the predominant species (supplementary figure 7). Thus, we hypothesised that a senescence-like programme might be activated. First, we analysed pH 6 senescence-associated β-galactosidase activity (SA β-gal)30: while isolated acini were negative, the suspension cultures were SA β-gal positive from D5 on (figure 4A). Expression of the senescence markers p53, p21, and Dec131 was markedly induced during culture as shown using both western blotting (figure 4B) and immunocytochemistry. These proteins and p16 showed heterogeneous staining (figure 4C). Double labelling confirmed that the exceptional cells undergoing proliferation failed to express p53 and Dec1 (supplementary figure 6). Similarly, SA β-gal was uniformly detected in monolayer cultures, in agreement with the very low proportion of Ki67+ cells (figure 4A and supplementary figure 6A).

Suspension and monolayer cultures activate a senescence-like growth arrest phenotype. (A) SA β-gal staining of isolated acini, D5 suspension, and D8 monolayer cultures. (B) Western blotting analysis of senescence marker expression in total pancreas, isolated acini, and suspension cultures at 24 h and D5. (C) Immunohistochemical analysis of senescence markers in isolated acini and D5 cultures.
Two important activators of senescence are DNA damage and oncogenic stress.15 We did not find evidence of phospho-γ-H2AX or p53BP foci, two markers of the DNA damage response (DDR) (supplementary figure 8). By contrast, we found increased levels of both total Ras and active Ras at D5, assayed by Raf1-GST pull-down (figure 5A). This was accompanied by ERK activation at 24 h and D5 of culture. By contrast, AKT was activated at a lower level (figure 5B).
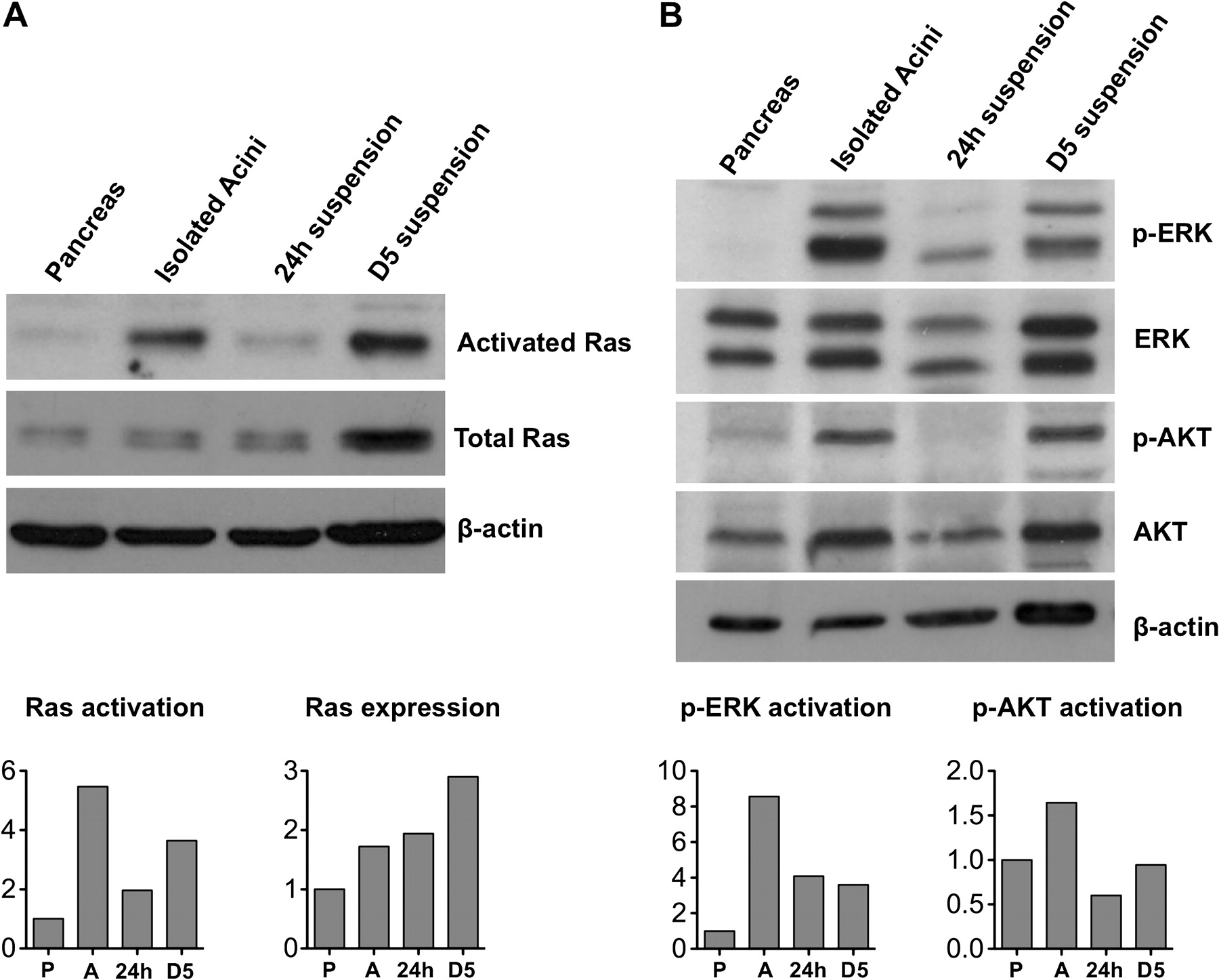
Activation of Ras and signaling in cultured acini. (A) Ras pull-down followed by western blotting to detect active Ras in extracts of pancreas, isolated acini, and suspension cultures at 24 h and D5 (first row). Western blotting analysis of total Ras and beta-actin in extracts of pancreas (P), isolated acini (A), and suspension cultures at 24 h, and D5 (second and third row). Quantification was performed by densitometric analysis using β-actin as reference. (B) Western blotting analysis of Ras downstream targets ERK and AKT, in total pancreas (P), isolated acini (A), and suspension cultures at 24 h and D5. Quantification was performed by densitometric analysis using β-actin as reference.
Transcriptome analysis of acinar suspension cultures reveals dedifferentiation, a specific gene expression pattern, and activation of a p53 response
To further characterise the suspension cultures, we analysed the transcriptome using expression arrays. Comparison of D5 cultures versus isolated acini revealed that approximately 1500 genes were differentially expressed (figure 6A upper). To globally assess the status of the adult differentiated acinar transcriptome, we examined changes in genes that have been shown to be differentially expressed in Rbpjl null mice12: 21/25 transcripts encoding digestive enzymes and secretory proteins, 7/8 transcripts encoding zymogen granule, packaging, or secretory machinery proteins, and 3/5 transcripts encoding proteins involved in mitochondrial metabolism were downregulated with p values <0.05 (supplementary table 1). We also compared their transcriptome with that of isolated mouse pancreatic ductal cells: approximately 7300 genes were differentially expressed (figure 6A upper) and several of the bona fide ductal markers were significantly downregulated in D5 acinar cultures (supplementary table 2), indicating that suspension cultures do not display a ductal phenotype. The majority of genes (ca. 95%) upregulated in ducts versus isolated acini were absent from the D5 versus isolated acini differentially regulated genes, further supporting their distinct phenotype (figure 6A lower), as also shown by principal component analysis (figure 6B).
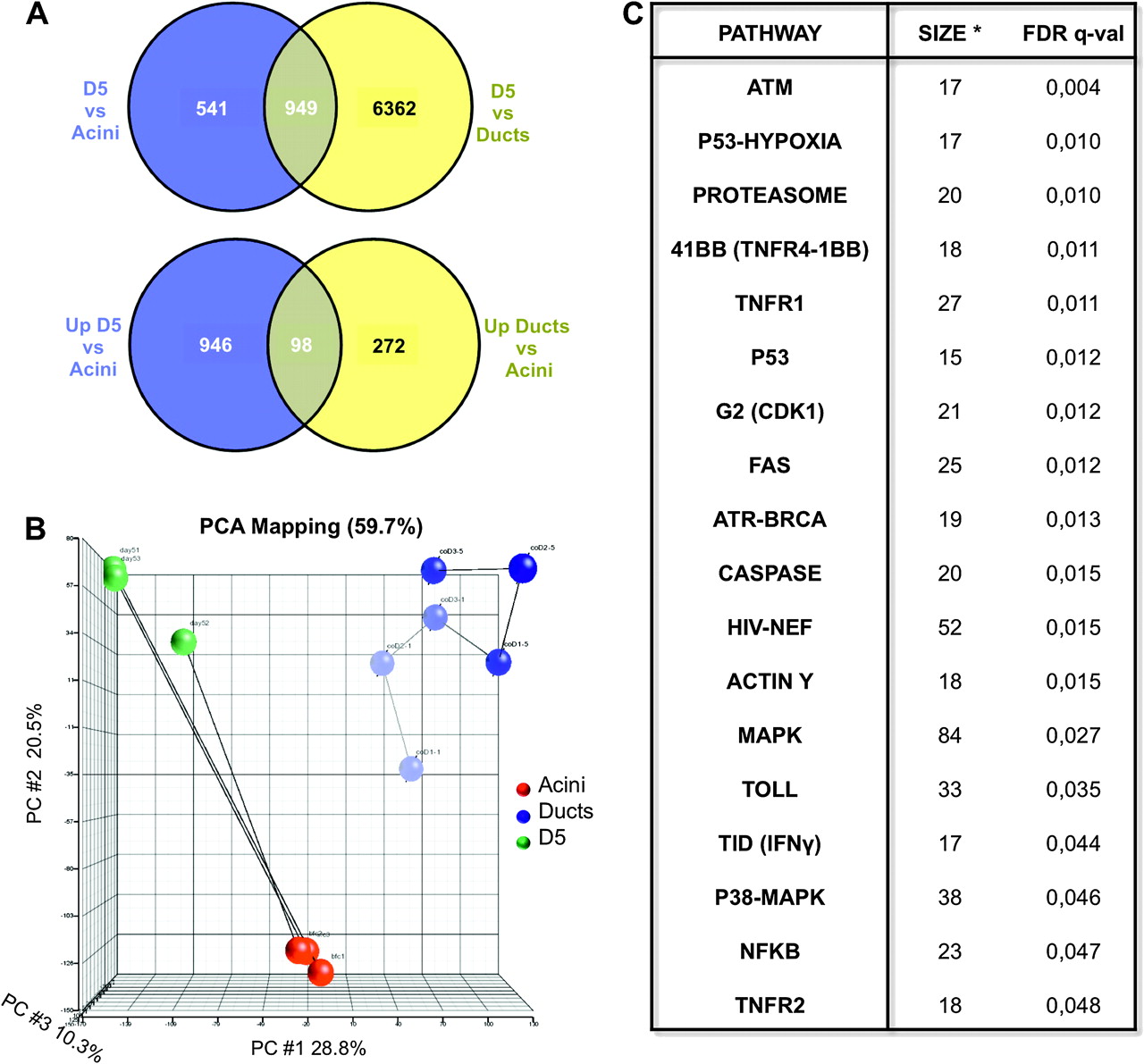
Transcriptome analyses of D5 suspension cultures. (A) Upper panel: Venn diagram showing the overlap of differentially expressed genes in D5 cultures compared with isolated acini and isolated ductal cells. Lower panel: Venn diagram showing the overlap of differentially upregulated genes in D5 cultures compared with isolated acini and differentially upregulated genes in isolated ducts compared with isolated acini. Genes were considered significantly regulated when presenting a fold-change >2 and after adjusting for multiple testing (FDR<0.01). (B) Principal component analysis of isolated acini, isolated ductal cells, and D5 cultures. (C) Pathway analysis of genes differentially regulated genes in D5 cultures compared to isolated acini. ‘Size’ (*) refers to the number of genes contained in the gene set whose expression was significantly enriched. Differences were considered significant after adjusting for multiple testing (FDR<0.05).
Pathway analysis of genes differentially expressed in D5 cultures versus isolated acini using Biocarta annotation showed a significant upregulation of several pathways downstream of p53, including genes involved in cell cycle arrest, checkpoint activation, p53-hypoxia, and apoptosis. In agreement with these data, the modules corresponding to MAPK and p38-MAPK pathways were also upregulated in D5 cultures. The main differentially regulated pathways are shown in figure 6C.
Activation of both a progenitor phenotype and a senescence programme in experimental CP
To assess whether the above findings are relevant to disease, we used two models of experimental CP: PDL and CaeCP. In PDL, the distal ligated tissue is mainly occupied by glands with a wide lumen that are lined by flat cells and by tubular structures composed of small cells showing branching and anastomoses. Residual acini are very rare but structures reminiscent of acini with a widened lumen are observed. The majority of cells in the ligated region expressed Ptf1a, Pdx1, and the embryonic markers Hes1, Sox9, Hnf1b, and Foxa2, and Krt19, supporting a progenitor phenotype (supplementary figure 9B). These findings are in agreement with, and extend, prior observations. A similar, but distinct, cellular composition was observed in CaeCP where most acinar glands showed a widened lumen and expression of Ptf1a, Pdx1, Sox9, with partial expression of cytoplasmic Hes1 and absence of Hnf1b; intermingled among these glands were tubular complexes (supplementary figure 9B). qRT-PCR analysis confirmed these changes in both models (supplementary figure 9A).
Regarding senescence, SA β-gal activity was detected in metaplastic ducts in the ligated pancreas, but not in the unligated pancreas, in the PDL model. Using immunohistochemistry, p53, p19, and Dec1 were exclusively detected in the ligated region with variable degrees of heterogeneity, as in cultured cells (figure 7A). In this model, 8.2±1.8% (n=6) of epithelial cells were Ki67+; however, p53 was absent from the latter cell population (figure 7B).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Expression of senescence markers in experimental chronic pancreatitis (CP). (A,B) Pancreatic duct ligation model. (A) SA β-gal staining and expression of p53, p19, and Dec1 using immunoperoxidase. (B) Double immunofluorescence analysis of the proliferation marker Ki67 and the senescence marker p53. Nuclei were counterstained with DAPI. (C) CaeCP model. Expression of p53, p21 and Dec1 using immunoperoxidase. Original magnification, ×200. DAPI, 4′-diamidine-2′-phenylindol dihydrochloride.
In the CaeCP model, p53 and p21 were upregulated in damaged areas, especially in those retaining morphological appearance of damaged acini; Dec1 was over-expressed with a broader pattern (figure 7C). However, we did not detect SA β-gal staining. In this model, a higher proliferation rate was observed (12.4±1.1% of Ki67+ cells among epithelial cells). This is in accordance with a more restricted activation of expression of senescence markers. Altogether, the results indicate that in experimental CP increased proliferation and activation of a senescence programme occur concomitantly in distinct cell populations.
Discussion
Adult pancreatic acinar cells from humans and rodents display cellular plasticity and can acquire ductal,16–18 endocrine,32–34 and hepatic35 fates. Embryonic acinar cells are permissive for transformation by Kras2 oncogenes whereas their adult counterparts are not.3 Based on these observations, it is conceivable that adult acinar cells can acquire a less differentiated phenotype rendering them susceptible to develop into PanINs or PDAC.
We explored the ability of acinar cells to dedifferentiate, rather than transdifferentiate, in suspension cultures because stem cells are more efficiently propagated forming ‘spheres’36 and provide novel evidence indicating that adult acinar cells dedifferentiate and acquire features of embryonic pancreatic progenitors. We reach this conclusion using stringent molecular criteria: (1) Pdx1 and Ptf1a are differentially expressed in the endocrine and exocrine adult compartments yet are uniquely co-expressed in embryonic pancreatic progenitors8 and upon culture in suspension, most cells co-express both proteins; and (2) in these cells the aPTF1 complex is replaced by the ePTF1 complex, which is found on bona fide promoters on which it is active in embryonic pancreas.10 11 This is accompanied by the expression of low levels of ‘early’ digestive enzyme transcripts – such as Cpa1 and Ctrb123 24 – and the virtual absence of ‘late’ digestive enzyme transcripts (ie, Amy2 or Ela1). While the detection of transcript or protein might not represent newly synthesised molecular species, the demonstration by ChIP of the occupancy of the Cpa1 promoter by proteins of the ePTF1 complex, but not of the aPTF1 complex, strongly supports our conclusions. This phenotype, together with the transcriptomic analyses, is reminiscent of the findings in Rbpjl null mice which display a failure in complete acinar differentiation.12
The majority of cells in the suspension cultures express Ptf1a, Pdx1, Hes1, Cpa1, Ctrb1, Sca1, Aldh1a, Cd133, FoxA1/2 and ductal keratins; Sox9 and Hnf1b have a more restricted expression, pointing at a certain heterogeneity which may, in part, be explained by the contribution of centro-acinar cells. Other markers such as Cck2r and Bmi1 were not considered here but have been identified by us in a comparable culture model and they are also expressed in embryonic pancreatic precursors.37 38
Several studies have reported that acinar cells can acquire ductal features in vitro.16–19 The lack of expression of late acinar markers together with the upregulation of Krt19, Krt7, and Klf4 – features that are generally used to assess ductal cell differentiation – might suggest an acinar–ductal phenotypic switch. However, our analyses indicate that the phenotype of suspension cultures corresponds to that of embryonic pancreatic progenitors, indicating that cells undergo dedifferentiation rather than transdifferentiation. One important finding that needs further analysis is that adherence cues prevent the acquisition of this phenotype and favour a ductal-like differentiation: cells cultured as a monolayer turned down the expression of Pdx1, Ptf1a and all digestive enzymes analysed to undetectable levels, unlike in suspension cultures. Mammospheres deposit an extracellular matrix that is similar to that of the developing mammary gland and helps cell survival in suspension.39 Recent work strongly supports the notion that integrin-mediated mechano-sensing can guide the stem cell versus differentiated phenotype of epidermal cells.40 Similar mechanisms participate in the control of cell fate decisions in the pancreas through control of microlumen formation during development.41
The observation that an embryonic phenotype is associated with lack of proliferation led us to analyse which mechanisms limit cell growth in this system. Apoptosis, demonstrated by the detection of activated caspase 3, was found to be only a minor contributor, leading us to consider the activation of a senescence programme.15 Indeed, we found an upregulation of classical senescence markers during in vitro culture, in marked contrast with their absence in normal pancreas tissue. Senescence can be activated by oncogenic stress, oxidative damage, DNA damage, and telomere shortening.15 The latter is unlikely to contribute since once cells are placed in culture, there is no proliferation. However, we found strong activation of Ras and ERK signalling upon acinar cell isolation and culture, likely as a result of cell detachment of matrix, growth factor stimulation and/or oxidative stress. We have used several experimental approaches to prevent KRas activation in our model, in order to causally establish its role in the induction of senescence. However, these attempts have been unsuccessful. During acute pancreatitis acinar cells acquire a transient ductal phenotype with complete recovery of function. However, mutant KRas can subvert the recovery of the acinar phenotype by interfering with the effects of β-catenin.42 Our results underscore that activation of endogenous wild-type KRas may suffice, in the absence of oncogenic mutations, to induce acinar dedifferentiation and are in accordance with previous studies showing that increased Ras activity levels can contribute to disease, including senescence induction.5 Of note, we have found that patients with wild-type KRAS PDAC are more likely to have a history of CP.43 p53 and p16 have been shown to play critical roles in senescence and they can limit the proliferation potential of progenitor and stem cells, including iPS.44 45 Preliminary experiments indicate that acinar suspension cultures from Tp53 knock out mice display increased proliferation and can be subcultured (unpublished data).
The activation of senescence suggests that tumour suppressive mechanisms may be activated in dedifferentiated acinar cells upon stress and can be relevant to the development of PDAC among patients with CP. To determine whether our in vitro observations are relevant to CP, we analysed two experimental CP models and found that both in PDL and in CaeCP, two processes take place: a fraction of cells undergoes increased proliferation while other cells activate a senescence-like programme and display progenitor marker expression. These two phenotypes are mutually exclusive as shown by Ki67 labelling and p53 expression. Thus, activation of a senescence programme in CP can provide a tumour-protective mechanism to normal adult cells. Yet, a proportion of these cells appear to not activate such programme and therefore may more effectively contribute to tumour initiation. A detailed analysis of the acinar cell contribution and the extent of dedifferentiation in CP is warranted.
In summary, a detailed molecular–functional analysis of cultured acinar cells has unveiled their ability to acquire an embryonic progenitor-like phenotype in conditions of stress-induced Ras signalling. Experimental CP models have further highlighted the complex concomitant activation of proliferative and senescence programmes in the pancreas in chronic inflammatory conditions. Our findings are important to better understand the differentiation potential of acinar cells, the physiopathology of CP, and the mechanisms involved in PDAC development.
Acknowledgments
We are grateful to N. del Pozo, D. Pastor, Y. Cecilia, M. Moreno, E. De Blay, M. Baekelandt, I. Houbracken and W. Rabiot for technical assistance, to J. Earl and G. Gómez for help with the analysis of the microarray data, and A. Skoudy for valuable comments to the manuscript. We are also grateful to the investigators who provided materials and reagents used in the study.
References
Supplementary materials
online only appendix
Files in this Data Supplement:
Footnotes
↵* AVP and IR made equivalent contributions and share first authorship.
Funding This work was supported, in part, by grant SAF2007-60860 and ONCOBIO Consolider from Ministerio de Ciencia e Innovación (Madrid, Spain) to FXR; grant EU-Marie Curie-ERG 204582 to IR; grant FWO G 0480 06 N to LB. AVP was supported by grant SFRH/BD/15902/2005 from Foundation for Science and Technology (FCT), Portugal and is a fellow of the Graduate Program in Areas of Basic and Applied Biology (GABBA), University of Porto, Portugal. IR was supported, in part, by the Fund for Scientific Research Flanders (FWO) and by the Francqui Foundation. There was also support from NIH RO1 DK060694 (AKR, MR) and National Pancreas Foundation (MR).
Competing interests None.
Ethics approval All mouse experiments were performed in accordance with institutional ethics committees and national guidelines and regulations.
Provenance and peer review Not commissioned; externally peer reviewed.