Article Text

Abstract
Objective Colonic ischaemia is frequently observed in clinical practice. This study provides a novel insight into the pathophysiology of colon ischaemia/reperfusion (IR) using a newly developed human and rat experimental model.
Design In 10 patients a small part of colon that had to be removed for surgical reasons was isolated and exposed to 60 min of ischaemia (60I) with/without different periods of reperfusion (30R and 60R). Tissue not exposed to IR served as control. In rats, colon was exposed to 60I, 60I/30R, 60I/120R or 60I/240R (n=7 per group). The tissue was snap-frozen or fixed in glutaraldehyde, formalin or methacarn fixative. Mucins were stained with Periodic Acid Schiff/Alcian Blue (PAS/AB) and MUC2/Dolichos biflorus agglutinin (DBA). Bacteria were studied using electron microscopy (EM) and fluorescent in situ hybridisation (FISH). Neutrophils were studied using myeloperoxidase staining. qPCR was performed for MUC2, interleukin (IL)-6, IL-1β and tumour necrosis factor α.
Results In rats, PAS/AB and MUC2/DBA staining revealed mucus layer detachment at ischaemia which was accompanied by bacterial penetration (in EM and FISH). Human and rat studies showed that, simultaneously, goblet cell secretory activity increased. This was associated with expulsion of bacteria from the crypts and restoration of the mucus layer at 240 min of reperfusion. Inflammation was limited to minor influx of neutrophils and increased expression of proinflammatory cytokines during reperfusion.
Conclusions Colonic ischaemia leads to disruption of the mucus layer facilitating bacterial penetration. This is rapidly counteracted by increased secretory activity of goblet cells, leading to expulsion of bacteria from the crypts as well as restoration of the mucus barrier.
- Colonic diseases
- mucosal injury
- intestinal barrier function
- ischaemia-reperfusion
- mucosal barrier
- immune response
- apoptosis
- gastrointestinal surgery
- inflammatory bowel disorders
- gut inflammation
- inflammatory mechanisms
- intestinal gene regulation
- nutrition
- abdominal surgery
- inflammation
- infant gut
- intestinal epithelium
- epithelial differentiation
- epithelial barrier
- epithelial cells
- small intestine
- intestinal development
- epithelial proliferation
- colorectal cancer
Statistics from Altmetric.com
- Colonic diseases
- mucosal injury
- intestinal barrier function
- ischaemia-reperfusion
- mucosal barrier
- immune response
- apoptosis
- gastrointestinal surgery
- inflammatory bowel disorders
- gut inflammation
- inflammatory mechanisms
- intestinal gene regulation
- nutrition
- abdominal surgery
- inflammation
- infant gut
- intestinal epithelium
- epithelial differentiation
- epithelial barrier
- epithelial cells
- small intestine
- intestinal development
- epithelial proliferation
- colorectal cancer
Significance of this study
What is already known on this subject?
-
The colonic mucus layer is critical for the spatial separation of luminal microbiota and the epithelium.
-
Breaches in the mucus layer are associated with colitis.
-
Colonic ischaemia/hypoxia can result in colitis.
What are the new findings?
-
This study is the first to provide insight into the pathophysiology of human colonic ischaemia/reperfusion.
-
Colonic ischaemia induces detachment of the mucus layer from the surface epithelium, facilitating penetration of bacteria into the otherwise sterile crypts.
-
Goblet cell compound exocytosis is associated with clearance of bacteria from the colonic crypts and rapid restoration of the mucus barrier.
-
These mechanisms limit exposure of colonic epithelium to potential intraluminal threats, thereby limiting inflammation.
How might it impact on clinical practice in the foreseeable future?
-
Improved understanding of the aetiology of ischaemic colitis is essential to develop preventive and/or therapeutic strategies.
-
Such knowledge might improve treatment modalities for inflammatory bowel disease (IBD), since epithelial hypoxia is a well-known pathophysiological phenomenon in the inflamed intestine of patients with IBD.
Introduction
The colonic intraluminal content is highly proinflammatory, harbouring an impressive density of 1012 bacteria per gram of luminal content in the distal colon.1 To prevent exposure to this potentially toxic luminal content and consequently a severe inflammatory response, the colon is equipped with several lines of defence. The mucus layer is the first anatomical site at which the host encounters the gut microbiota.2 ,3 This mucus layer serves as a physical and chemical barrier, preventing adherence of microbiota to the epithelium and translocation of potential pathogens and their toxins to the internal milieu.2–4
The properties of mucus are derived from the major gel-forming glycoprotein components called mucins.2 ,4 The secreted mucins are continuously produced by specialised goblet cells that are found in large numbers in the colonic epithelium. In the endoplasmic reticulum (ER) and Golgi, mucins are oligomerised and glycosylated to give them their viscous and protective properties.4 MUC2 is the most abundantly expressed secretory mucin in the colon and is stored in bulky apical granules of the goblet cells which form the characteristic goblet cell thecae.4 The mucin-containing granules are secreted from the apical surface both constitutively and in response to a variety of external stimuli.2 ,4 In addition, goblet cells can undergo so-called compound exocytosis, an accelerated secretory event resulting in the acute release of centrally-stored mucin granules.4 ,5
The importance of the mucus barrier is underlined by studies showing that mice deficient in MUC2, which is the structural component of the mucus layer, develop spontaneous chronic intestinal inflammation in the colon.6 In addition, dextran sulfate sodium (DSS)-induced chronic colonic inflammation in mice, a model for human inflammatory bowel disease (IBD), is preceded by breakdown of the mucus barrier and adherence of microbiota to the epithelium.7 Last, missense mutations in the MUC2 gene leading to aberrant MUC2 oligomerisation and glycosylation results in decreased barrier function and chronic colonic inflammation,8 which resembles the morphological and inflammatory changes observed in IBDs. These data show that both the quantity and quality of MUC2 is of utmost importance for the protective properties of the mucus layer and thereby for the mucus barrier capacity.
Colonic ischaemia/reperfusion (IR) is a frequently observed event that can potentially harm the colonic barrier function. Colonic ischaemia is the most common form of intestinal ischaemia and is observed in a number of situations including infections and vasculitis as well as in patients undergoing aortic surgery or cardiac bypass surgery.9–12 In addition, intestinal hypoxia is a well-known pathophysiological phenomenon in the (chronically) inflamed mucosa, such as mucosal inflammation observed in IBD.13 ,14
To study the pathophysiology of human colonic IR we developed a new human experimental colon IR model, analogous to a recently developed model for human small intestinal IR.15–17 The observed effects of IR were studied in more detail in a new rat colonic IR model in which we investigated the mucus barrier integrity, bacterial penetration and the response of mucus-producing goblet cells to increased bacterial threats. In addition, we investigated the colonic IR-associated development of intestinal inflammation.
Materials and methods
Patients and experimental procedures
Patients
For the experimental colonic IR protocol we took advantage of the fact that, during low anterior resections or abdominoperineal resections, part of the healthy colon (proximal to the tumour) has to be removed for surgical reasons. Ten patients were included. Patients with underlying intestinal disease were excluded. During surgery the segment of colon that had to be removed for surgical reasons was identified and the proximal 6 cm of this segment was carefully isolated using a cutting stapler (see supplementary figure 1). Next, the isolated segment was exposed to 60 min of ischaemia (60I) by placing two atraumatic vascular clamps across the mesentery. Meanwhile surgery continued as planned. After 60 min of ischaemia a small part of the isolated segment was stapled off and collected for research purposes, after which reperfusion of the remaining isolated segment was allowed by removal of the vascular clamps. Tissue was again collected at 30 and 60 min of reperfusion (30R and 60R, respectively). Owing to the duration of the surgical procedure it was not possible to investigate longer reperfusion periods. At the end of the experimental protocol a small part of healthy colon that was not exposed to IR was resected and served as internal control.
Tissue collection
Colonic tissue samples were immediately snap-frozen for quantitative PCR (qPCR), formalin-fixed for histology and immunohistochemistry or fixed in glutaraldehyde for electron microscopy (EM).
Animal studies
Male Sprague-Dawley rats weighing 300–350 g were purchased from Charles River Laboratories (Maastricht, The Netherlands) and housed under controlled conditions of temperature and humidity. Throughout the experiments, rats had unrestricted access to standard rodent chow and drinking water. The rats were randomly divided into groups of seven animals and exposed to the experimental protocols, as described below. Animals were anaesthetised with isoflurane (induction 4%, maintenance 1.5%) and buprenorfine (0.1 mg/kg) was used for analgesia.
Colonic IR
Following midline incision, a 10 cm colon segment directly distal to the ceacum was isolated and ligated at both ends. The mesocolon was carefully dissected and the segment was subjected to 60 min of ischaemia (60I) by placing atraumatic vascular clamps across the mesenteric branches. After ischaemia, the isolated segment was either directly resected (60I) or the clamp was removed to allow reperfusion for 30, 120 or 240 min (30R, 120R, 240R). Reperfusion was confirmed by regaining of normal colour and restoration of gut motility. Control colonic segments were obtained from rats exposed to similar surgical handling without exposing the colon segments to IR.
Tissue collection
Colonic tissue samples were immediately snap-frozen for quantitative PCR (qPCR) or fixed in methacarn solution to preserve the mucus layer (protocols for methacarn fixation were kindly provided by Dr G Hansson).3 All formaldehyde-fixed and methacarn-fixed tissue samples were embedded in paraffin and 4 μm sections were cut. Tissue of two animals per group was fixed in glutaraldehyde instead of methacarn for EM.
Histology and immunohistochemistry
For morphological analysis, sections were deparaffinised in xylene and rehydrated in graded ethanol to distilled water and stained with H&E and with Periodic Acid Schiff (PAS) and Alcian blue (AB).
For immunohistochemistry, sections were deparaffinised in xylene and rehydrated in graded ethanol to distilled water. Endogenous peroxidase activity was blocked using 0.6% hydrogen peroxide in methanol for 15 min. Non-specific antibody binding was blocked and sections were incubated with primary antibody rabbit anti-human myeloperoxidase (DakoCytomation (Glostrup, Denmark), cross-reacts with rat). Biotin-conjugated swine anti-rabbit IgG (DakoCytomation) was used followed by incubation with the streptavidin-biotin-HRP system (DakoCytomation). Binding of primary antibody was visualised with 3,3′-diaminobenzidine-tetrahydrochloride-dihydrate (Sigma, St Louis, Missouri, USA) and counterstained with haematoxylin. No staining was detected in slides incubated without primary antibody.
Details of quantification of goblet cell increased secretory activity are shown in the online supplement.
Immunofluorescence
Sections were deparaffinised in xylene and rehydrated in graded ethanol to distilled water. Antigen was retrieved by incubating the slides in 10 mM sodium citrate (pH 6) at 90–100°C. Slides were reduced using 10 mM dithiothreitol in 0.1 M Tris-HCl (pH 8) for 30 min at room temperature followed by alkylation using 25 mM iodoacetamide in 0.1 M Tris-HCl (pH 8) in the dark for 30 min at room temperature. After blocking non-specific binding sites, sections were incubated overnight at 4°C with polyclonal rabbit anti-rat MUC2 antiserum. Next, slides were washed and then incubated with FITC-conjugated Dolichos biflorus agglutinin (DBA, Vector (Burlingame, CA, USA)) and secondary antibody Cy3-labelled goat anti-rabbit IgG (Invitrogen, Carlsbad, California, USA). Slides were counterstained with 4',6-diamidino-2-phenylindole (DAPI) and mounted using the fluorescent mounting medium (DakoCytomation).
Fluorescent in situ hybridisation (FISH)
Sections were deparaffinised, incubated with ethanol and dried for 15 min. They were then incubated overnight at 37°C in the dark with the highly specific Cy3-conjugated bacterial probe in hybridisation solution (20 mM TrisHCl, 0.9 M NaCl, 0.1% SDS). The bacterial probe (EUB338, Eurogentec (Seraing, Belgium), The Netherlands) recognises ribosomal RNA (rRNA) of most bacteria. Next, sections were washed and counterstained with DAPI and mounted using the fluorescent mounting medium (DakoCytomation).
Details of quantification of bacteria in colonic crypt base epithelium and colonic surface epithelium are shown in the online supplement.
RNA isolation and quantitative PCR
RNA was isolated from snap-frozen tissue samples using AllPrep DNA/RNA/Protein kit (Qiagen, Hilden, Germany) according to the manufacturer's protocol. In short, colon samples were crushed with a pestle and mortar in liquid nitrogen. Disruption and homogenisation of the tissue was performed using an Ultra Turrax Homogeniser (IKA Labortechnik, Staufen, Germany) in lysis buffer containing β-mercaptoethanol (Promega, Madison, Wisconsin, USA). RNeasy spin columns were used to bind RNA. Columns were washed and RNA was eluted in RNase-free water. To analyse gene expression, qPCR was performed. RNA samples were treated with DNAse (Promega) to ensure removal of contaminating genomic DNA. RNA quantity was measured using the NanoDrop spectrophotometer (Thermo Scientific, Wilmington, Deleware, USA). Total cDNA was synthesised using the iScript cDNA synthesis kit (Bio-Rad, Hercules, California, USA). qPCR reactions were performed in a volume of 20 μl containing 10 ng cDNA, 1× IQ SYBR Green Supermix (Bio-Rad) and 300 nM of gene-specific forward primers and reverse primers. The sequence of primers is shown in table 1 in the online supplement. cDNA was amplified using a three-step cycling programme (40 cycles of 10 s at 95°C, 20 s at 60°C and 20 s at 70°C) with a MyiQ system (Bio-Rad). Specificity of amplification was verified by melt curve analysis. Gene expression levels were determined using iQ5 software using a ∆Ct relative quantification model. The geometric mean of internal control genes (see table 1 in online supplement) was calculated and used as a normalisation factor.
Electron microscopy (EM)
For EM, colonic tissue was immersed in 3% glutaraldehyde fixative buffered in 0.09 M KH2PO4 at pH 7.4. Samples were then washed in 0.09 M KH2PO4 buffer with 7.5% sucrose and transferred to a 1% OsO4 + 1.5% ferrocyanide solution and buffered to pH 7.4 with 0.1 M veronalacetate for subsequent post-fixation for 1 h at 4°C. After washing in phosphate veronalacetate buffer containing 7% sucrose at pH 7.4, dehydration was carried out rapidly in graded ethanol series followed by embedding in Epon. Tissue sections were cut and examined with a Philips CM 100 electron microscope.
Statistical analysis
Statistical analysis was performed using Prism 5.0 for Windows (GraphPad Software, San Diego, California, USA). In human studies, the Dunn multiple comparison test was used to compare values in time and the Wilcoxon signed rank test was used for between-group comparisons. In rat studies, the Mann–Whitney U test was used for between-group comparisons. All data are presented as mean±SE. A p value <0.05 was considered statistically significant.
Results
Human colonic IR is associated with limited tissue damage and increased goblet cell secretory activity
Directly after 60 min of ischaemia, subepithelial spaces were observed in the H&E staining at the surface epithelium of the colon (figure 1A, right upper panel, arrows), while these spaces were not present in control tissue (figure 1A, left upper panel). Early during reperfusion (30R), IR-damaged epithelial cells were shed into the lumen (figure 1A, left lower panel, arrowhead). After 60 min of reperfusion, the epithelial lining had an irregular appearance but extensive damage of the epithelial monolayer was not observed on H&E staining (figure 1A, right lower panel). Interestingly, it appeared that goblet cell numbers were reduced at 60 min of reperfusion. To assess goblet cells in more detail, goblet cell mucins were stained with PAS/AB. In control tissue, PAS/AB staining showed abundant presence of mucus within the goblet cell thecae (figure 1B). In line with this, EM of control colon showed normal distribution of secretory granules (figure 1C, left panel). In contrast, massive expulsion of goblet cell mucus was observed at 60 min of ischaemia in both the PAS/AB staining (figure 1B, arrowheads) and EM (figure 1C, middle and right panel, arrowheads, and supplementary figure 2A). During reperfusion, secretory activity of goblet cells continued, resulting in depleted goblet cell thecae at 60 min of reperfusion compared with control (figure 1B, arrow, and supplementary figure 2B). The high secretory activity of goblet cells was accompanied by a trend (p=0.07) towards increased mRNA expression of MUC2 at 30R (figure 1D).

Human colon ischaemia/reperfusion (IR) induces goblet cell compound exocytosis and increased secretory activity. (A) H&E staining shows normal colonic architecture in healthy colon (upper left panel). Colonic ischaemia was associated with the appearance of subepithelial spaces (upper right panel, arrows and asterisk in inset). At 30 min of reperfusion (30R), IR-damaged cells were shed into the lumen (lower left panel and inset, white arrowhead). At 60 min of reperfusion (60R), the epithelium appeared irregular but no significant gaps in the epithelial lining were observed (lower right panel). (B) Periodic Acid Schiff/Alcian Blue (PAS/AB) staining showed bulky presence of acidic mucins in goblet cells of healthy human colon (upper panel and inset). At 60 min of ischaemia (60I), goblet cells appeared to release their contents into the lumen (60I, white arrowhead and inset). This process continued at 30 min of reperfusion (60I 30R, see white arrowheads), which resulted in mucus-depleted goblet cells at 60 min of reperfusion (60I 60R, arrow). (C) Normal electron microscopic (EM) appearance of goblet cell granules in healthy human colon (left panel). In contrast, goblet cell compound exocytosis was observed at 60I in transversal sections (middle panel, red line demarcates the lumen (L)) and longitudinal sections (right panel, white arrowhead, note the expulsion of whole granules). (D) Next to the increased secretory activity, a trend towards increased mRNA expression of MUC2 was observed at 30R (p=0.07 vs control).
Mucus barrier loss results in bacterial adherence to the epithelium in rat colon exposed to 60 min of ischaemia
To obtain more insight into increased secretory activity of goblet cells following IR in the human colon, we set out to study the mucus layer and presence of microbiota in detail. For that purpose, we used a rat colon IR model in which we preserved the mucus layer using methacarn fixation.3 Of note, IR-induced epithelial damage in rat colon was comparable to epithelial damage observed in our human studies. PAS/AB staining as well as immunofluorescence for MUC2/DBA showed a thick highly-glycosylated mucus layer in control tissue (figure 2, A1 and B1). This mucus layer clearly separated intraluminal bacteria from the colonic epithelium at the surface and in the crypts, as shown by staining for bacterial rRNA (figure 2, C1 inset 1 and 2, respectively). In contrast, a disorganised damaged mucus layer was observed in rat colon exposed to 60I (figure 2, A2 and B2). This was accompanied by bacterial adherence to epithelial cells in both the surface epithelium (figure 2, C2 and inset 1) and in the base of the crypts (figure 2, C2 and inset 2). Quantification of the number of crypts colonised by bacteria showed bacterial presence in 80% of crypts at the surface epithelium while bacteria had penetrated into the crypt base in 55% of the crypts (figure 2D,E, respectively; p<0.01). In line with this, EM showed that whereas bacteria were not observed near the epithelium in control tissue (figure 2, F1), many bacteria were observed in close proximity to the epithelium in tissue exposed to 60 min of ischaemia (figure 2, F2 and F3, arrowheads, note bacteria in contact with microvilli). In addition, bacteria could be observed between epithelial cells (figure 2, F4).

Ischaemia-induced breaches in the mucus layer result in bacterial penetration into colonic crypts and adherence to the epithelium. (A1, B1) Periodic Acid Schiff/Alcian Blue (PAS/AB) and MUC2/Dolichos biflorus agglutinin (MUC2/DBA) staining show healthy highly-glycosylated mucins in control tissue. (C1) The mucus layer provides spatial separation (white arrows) of the colonic epithelium with intraluminal bacteria, stained in red with the specific probe for bacterial rRNA EUB338 (note in inset 1 and 2 of figure C1 that both surface epithelium and colonic crypts are devoid of bacteria in healthy tissue.) (A2, B2) In contrast, a disorganised breached mucus layer was observed in PAS/AB and MUC2/DBA staining of colon exposed to 60 min of ischaemia (60I). (C2) This facilitated bacterial adherence to the surface epithelium and penetration into the colonic crypts (white arrowheads and inset 1 and 2, respectively). (D) Increased bacterial colonisation of the crypt surface epithelium at 60I compared with control (from 4% to 80% of crypts, **p<0.01). (E) Colonisation of the crypt base increased from 0% in control tissue to 55% of crypts at 60I (**p<0.01). (F1) Electron microscopy showed that whereas bacteria were not observed in close proximity to the epithelium in healthy colon, (F2) many bacteria were observed near epithelial cells in colon exposed to 60I (red arrowheads) and bacteria appeared to make physical contact with the microvilli of epithelial cells (see inset in figure F2, white arrowhead). (F3) Moreover, bacteria could be observed in between epithelial cells following ischaemia (red arrowheads).
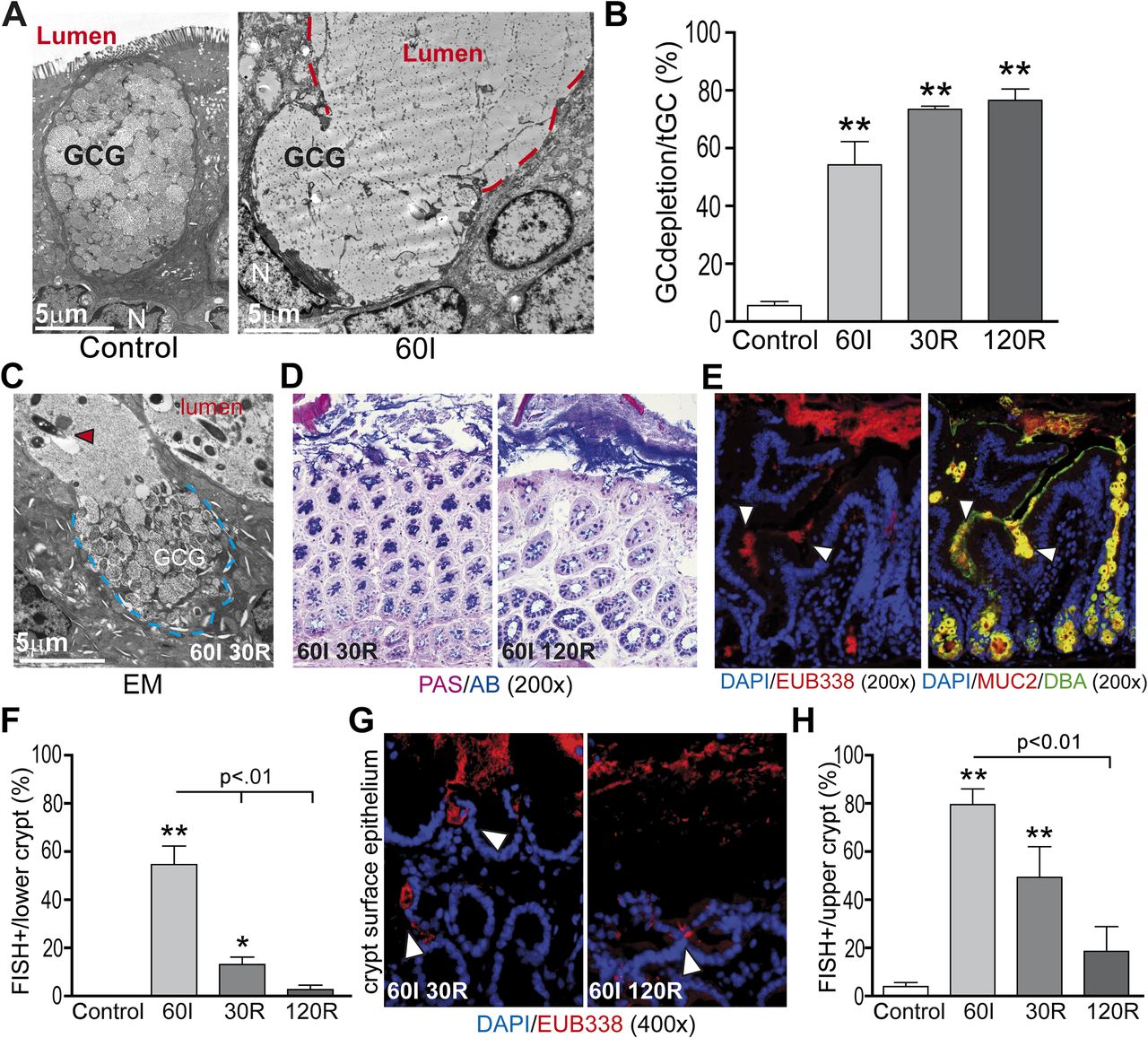
Increased goblet cell secretory activity is associated with expulsion of bacteria from the colonic crypts
In accordance with the human data, goblet cells in rat tissue exposed to ischaemia showed massive degranulation and expulsion of centrally stored mucins compared with control (figure 3A and B: 54% at ischaemia vs 6% in control tissue, p<0.01). Increased goblet cell secretory activity continued early during reperfusion as indicated by EM (figure 3C) and PAS/AB staining (figure 3D, left panel). Quantification of depleted goblet cell thecae showed that 73% and 77% of goblet cells exhibited signs of increased mucus secretion at 30R and 120R, respectively, compared with control (figure 3B, p<0.01). However, PAS/AB staining also revealed formation of a thin new mucus layer covering the surface epithelial cells at 120 min of reperfusion (figure 3D, right panel).
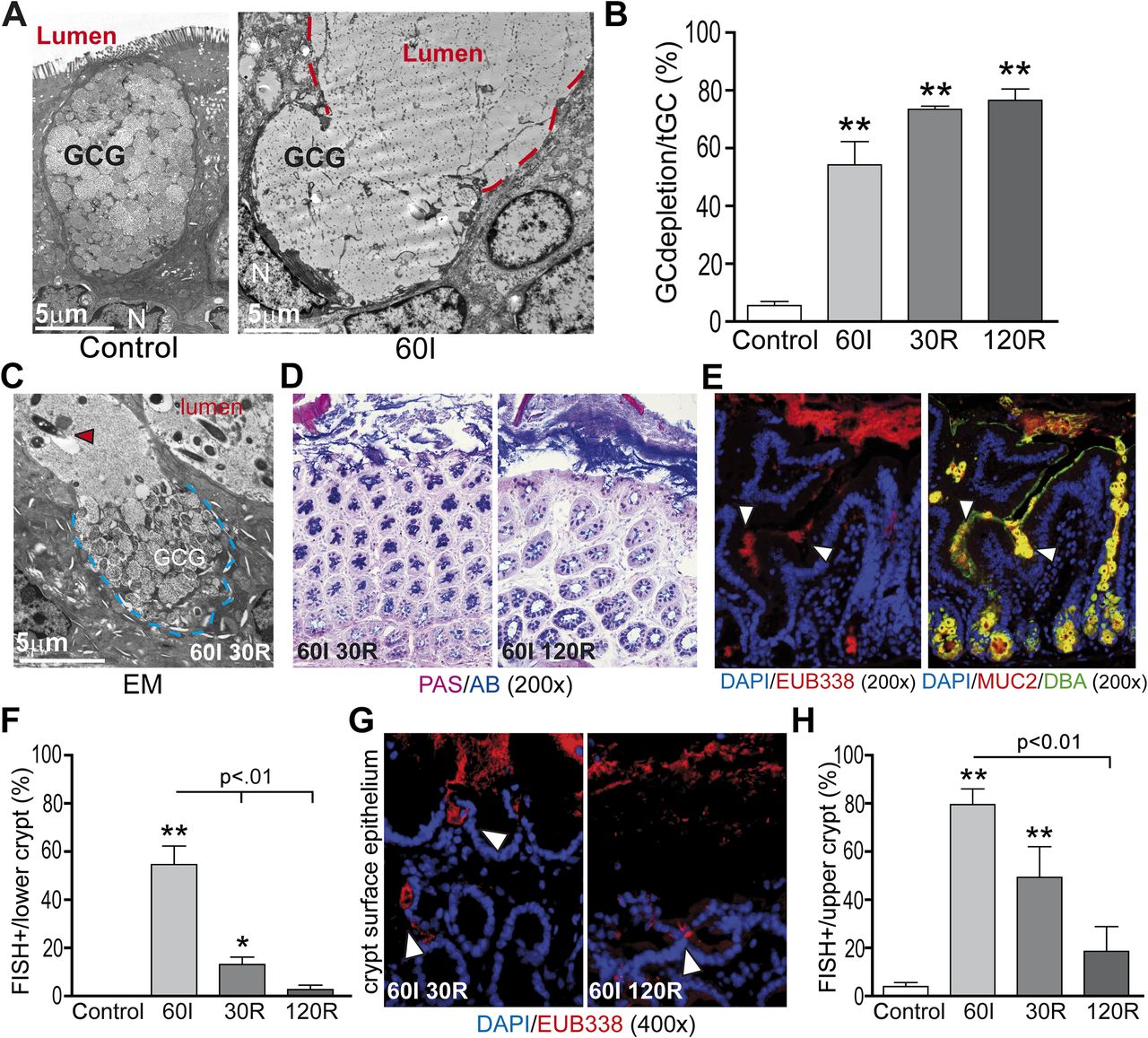
Increased goblet cell secretory activity is associated with expulsion of bacteria from the colonic crypts. (A) In line with human studies, electron microscopy showed that rat colon ischaemia was also accompanied by massive expulsion of goblet cell granules into the lumen (red line demarcates lumen; GCG, goblet cell granules; N, nucleus). (B) Simultaneously with bacterial crypt penetration at 60 min of ischaemia (60I), a significant increase in mucus-depleted goblet cells was observed as a sign of increased secretory activity (54% at 60I vs 6% in control tissue). During 30 and 120 min of reperfusion (30R and 120R, respectively), goblet cell mucus depletion further increased to 73% and 77%, respectively; **p<0.01. (C) Electron microscopy (EM) showed continued expulsion of mucus-containing granules at 30R (blue line demarcates goblet cell content, red arrowhead shows bacteria in close proximity to the epithelium). (D) Whereas a mucus layer was absent early during reperfusion (30R, left panel), a thin mucus layer had formed at 120R (right panel). (E) Staining for bacterial rRNA using probe EUB338 (left panel) and MUC2/Dolichos biflorus agglutinin (MUC2/DBA, right panel) in serial sections showed release of mucus at the sites where bacteria penetrate the colonic crypts (white arrowheads). (F) Simultaneous with increased mucus release, bacterial presence in the lower parts of the colonic crypts significantly fell from 55% at 60I to 13% and 3% at 30R and 120R, respectively (p<0.01). **p<0.01 vs control, *p<0.05 vs control. (G, H) In line with this, bacterial presence at the surface epithelium decreased over the course of reperfusion from 80% at 60I to 49% and 19% at 30R and 120R, respectively (p<0.01).
To investigate whether bacterial colonisation, as a result of mucus barrier loss, could be the trigger for the observed increased secretory activity of colonic goblet cells, bacterial penetration and mucus release were investigated in serial sections. Interestingly, at the sites where bacteria were observed near epithelial cells, massive expulsion of goblet cell contents was observed (figure 3E, serial sections, left panel: bacteria, right panel: mucus release). Further clues supporting the hypothesis that increased secretory activity is a response to bacterial colonisation were that increased goblet cell secretory activity preceded the reduction in bacterial crypt base colonisation during reperfusion (from 55% at ischaemia to 13% and 3% at 30R and 120R, respectively (figure 3F, p<0.01). In line with this, bacterial presence at the surface epithelium decreased significantly over the course of reperfusion (figure 3G and H, from 80% at ischaemia to 49% and 19% at 30R and 120R, respectively).
Increased goblet cell secretory activity is associated with restoration of the mucus layer during prolonged reperfusion
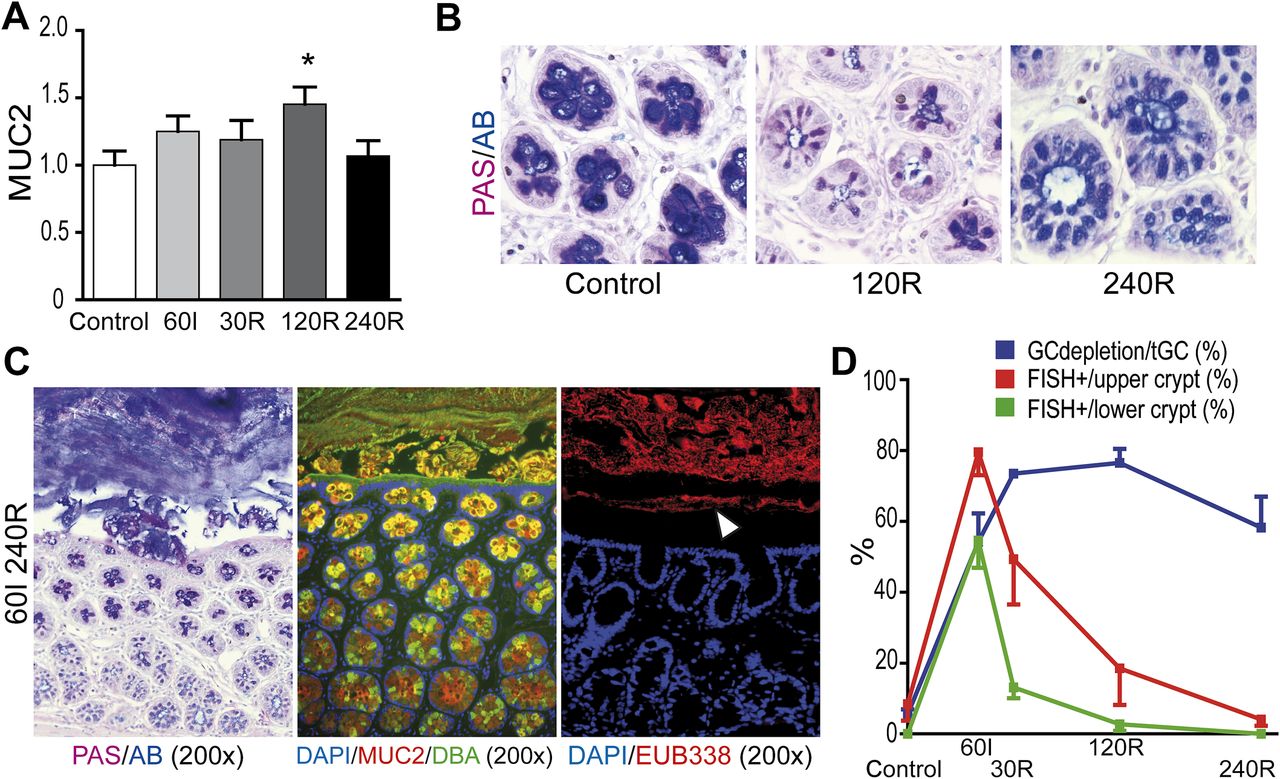
The increased goblet cell secretory activity was accompanied by increased mRNA expression of MUC2 at 120R compared with control (figure 4A, p<0.05).

Restoration of the mucus barrier separates intraluminal bacteria from the colonic epithelium after prolonged reperfusion. (A) Increased MUC2 mRNA expression at 120 min of reperfusion (120R; *p<0.05 vs control) indicates a restorative response following increased mucus secretion. (B) In healthy colon, bulky presence of acidic AB-positive mucins was observed in goblet cell thecae, whereas goblet cells appeared depleted of mucus at 120R. In line with MUC2 mRNA expression data, partial restoration of goblet cell contents was observed at 120 min of reperfusion (240R). (C) Periodic Acid Schiff/Alcian Blue (PAS/AB, left panel), MUC2/Dolichos biflorus agglutinin (MUC2/DBA, middle panel) and fluorescent in situ hybridisation (FISH) for bacterial rRNA (right panel) showed renewed spatial separation of bacteria (white arrowhead) from the colonic epithelium at 240R. (D) Plotting of both the percentage of depleted goblet cells and bacterial colonisation of the crypts shows that goblet cell mucus release occurs simultaneously with bacterial infiltration in the crypts. Increased goblet cell secretion during IR is associated with clearance of bacteria from the colonic crypts. GC, goblet cell; tGC, total goblet cell numbers.
Increased MUC2 expression led to partial recovery of the mucin content in goblet cell granules at 240R. In control colon, PAS+/AB+ mucus was abundantly present in goblet cell granules whereas depleted goblet cell thecae were observed at 120R. However, PAS/AB staining revealed recovery of mucin quantity at 240R (figure 4B).
In addition, a new mucus barrier was observed at 240R in PAS/AB staining and in immunofluorescent staining for MUC2/DBA (figure 4C, left and middle panel). This newly formed mucus layer efficiently separated luminal microbiota from colonic epithelial cells (figure 4C, right panel) since the presence of bacteria was no longer observed in either the lower or upper parts of the colon crypts at 240R (figure 4D). In conclusion, this set of experiments showed that IR-induced bacterial translocation is accompanied by increased goblet cell mucus secretion. The increased release of mucus is associated with expulsion of bacteria from the lower and upper parts of the crypts (figure 4D) and restoration of the mucus barrier.
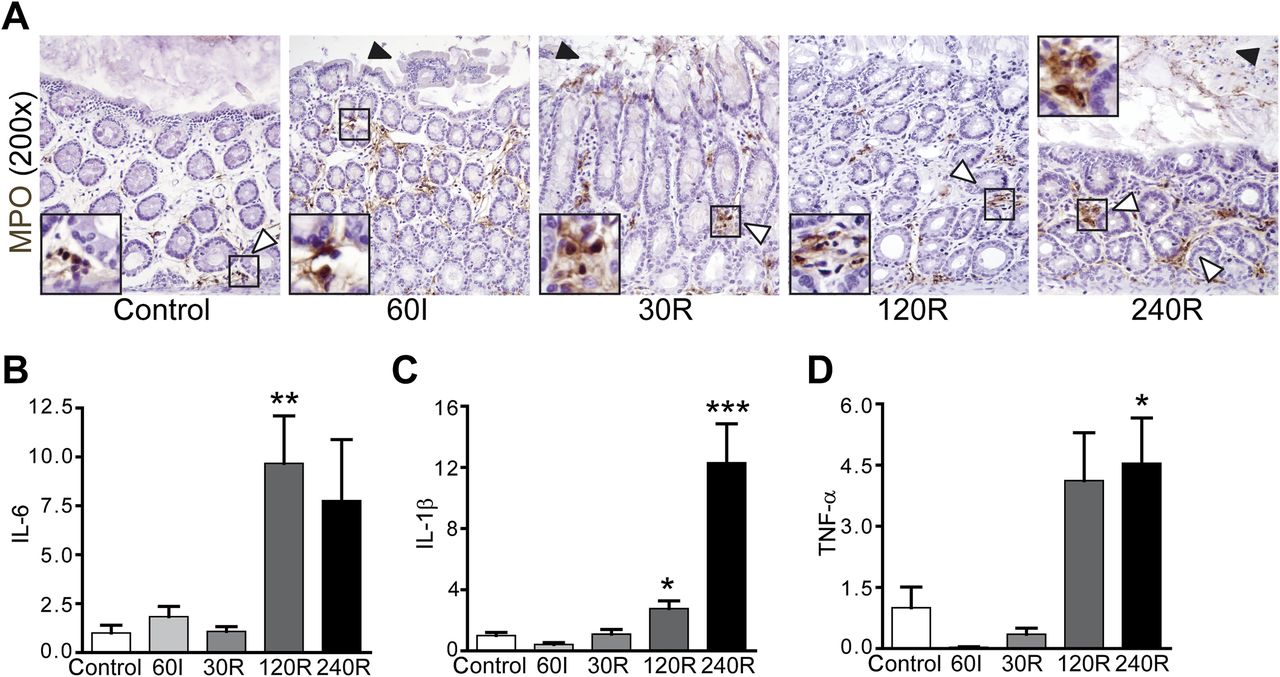
Rat colon IR results in influx of neutrophils and increased expression of TNFα, IL-6 and IL-1β
In human tissue, increased neutrophil influx was not observed at ischaemia or during reperfusion. In line with this, the expression of proinflammatory cytokines did not change over the course of IR (data not shown). This was expected since we were only able to investigate inflammatory responses over a short period of time in the human setting. We therefore investigated the inflammatory response in rat colon exposed to IR. A minor increase in myeloperoxidase-positive neutrophils was observed in rat tissue exposed to ischaemia with reperfusion, particularly at the prolonged stages of reperfusion (figure 5A, right panel). A significant increase in interleukin (IL)-6 (figure 5B, p<0.01) and IL-1β (figure 5C, p<0.05) mRNA expression was observed at 120R while, at 240R, mRNA expression of IL-1β (figure 5C, p<0.001) and tumour necrosis factor (TNF)α (figure 5D, p<0.05) was significantly increased.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Rat colonic ischaemia/reperfusion (IR) is associated with limited influx of neutrophils into colonic lamina propria and increased expression of proinflammatory cytokines. (A) Neutrophils were observed especially near the crypts of healthy control colon (white arrowhead). A marginal increase in neutrophils was observed in the lamina propria at 60 min of ischaemia (60I), 30 and 120 min of reperfusion (30R and 120R) (white arrows). IR-damaged cells were pinched off into the lumen during IR (black arrowheads). At 240R, increased neutrophil staining was observed in the lamina propria (white arrowheads). IR-damaged shed enterocytes were observed in the intestinal lumen (black arrowhead). (B–D) In line with myeloperoxidase (MPO) staining, an inflammatory response became apparent with longer duration of reperfusion, as indicated by increased expression of interleukin (IL)-6 and IL-1β at 120R and IL-1β and tumour necrosis factor (TNF)α at 240R. *p<0.05; **p<0.01; ***p<0.001.
Discussion
This study is the first to show that colonic ischaemia leads to loss of the mucus layer, an important first line of defence against intraluminal microbiota. Loss of the mucus layer allows bacteria to penetrate into colonic crypts and to adhere to epithelial cells. Intriguingly, goblet cells react rapidly in response to these bacterial threats with expulsion of their mucus-containing granules, probably in order to flush bacteria from the normally sterile crypts. Thus, we show that the colon has an efficient mechanism to prevent excessive exposure of the epithelium to bacteria following ischaemia-induced mucus barrier loss.
The effectiveness of the mucus layer as a barrier is emphasised by the spatial separation of the colonic microbiota from the epithelium in the healthy intestine.3 Loss of the mucus layer in MUC2-deficient mice has been shown to cause spontaneous colitis6 and to increase susceptibility to lethal infectious colitis induced by pathogenic and commensal flora.18 In addition, DSS-induced chronic colonic inflammation in mice, a model for human IBD, is preceded by mucus barrier breakdown and adherence of microbiota to the epithelium.7 In line with these studies, we report that ischaemia-induced breaches in the mucus layer result in bacterial adherence to epithelial cells and translocation of bacteria into the colonic crypts.
The mechanisms underlying the appearance of breaches in the mucus barrier are not fully understood. In the setting of ischaemia, shedding of ischemically damaged epithelial cells into the intestinal lumen might contribute to the detachment and/or breakdown of the mucus layer. In addition, detachment of mucus from epithelial cells might be due to changes in cellular homeostasis during hypoxia, altering the conditions for mucins to remain connected to the epithelium. For example, it has been demonstrated that proper expansion and dispersion of mucus is bicarbonate-dependent due to the cationic nature of mucins.19 ,20 In that respect, mucus detachment of the epithelial lining might be due to the development of an acidic milieu in epithelial cells during hypoxia leading to bicarbonate depletion and disaggregation of the mucus layer.
Interestingly, goblet cells responded directly to bacterial threats by so-called compound exocytosis, a process by which whole goblet cell content is rapidly expulsed into the lumen.2 ,4 ,5 This has been described as a mechanism to reinforce the barrier and exclude pathogens from the normally sterile crypts. Compound exocytosis results in a 100–1000-fold expansion in mucin volume upon hydration and contributes to the rapid replacement of a degraded mucus barrier.4 In this study we found compound exocytosis to be a successful mechanism to clear bacteria, since bacteria were flushed from the colonic crypt base and surface epithelium during reperfusion of ischaemic colon. In addition, massive secretion of mucus allowed rapid recovery of the mucus layer, providing renewed spatial separation of luminal bacteria and the epithelium at 240R in rats.
It is interesting to note that, although the colonic luminal content, with the high density of microbiota, is potentially much more toxic than the intraluminal content of the small intestine, colon ischaemia generally tends to have a much milder course. Inflammatory complications including sepsis and multiple organ failure rarely occur after an episode of colonic ischaemia, and up to 90% of all cases of colonic IR have a favourable outcome following conservative treatment.12 ,21–23 This is in sharp contrast with the outcome of small intestinal ischaemia,9 indicating that the colon has efficient mechanisms to prevent inflammatory complications following colonic IR. The protective mechanisms described here could explain in part the discrepancy between complications observed after small and large intestinal IR.
In addition to mucin quantity, the quality of mucins is also of major importance. Mucus quality is associated with correct glycosylation, sialylation and sulphation of mucins.4 Loss of O-glycosylation of mucins has been shown to induce spontaneous colitis in mice,24 and reduced glycosylation is also observed in ulcerative colitis in humans.8 ,25 In addition, impaired glycosylation has recently been linked to endoplasmic reticulum stress in mice and humans.8 Although we did not study alterations in mucin quality in this acute model of intestinal ischaemia, we observed significant endoplasmic reticulum stress and activation of the unfolded protein response in the human and rat colon exposed to IR (data not shown). Future studies should be aimed at investigating whether this could be associated with decreased mucus quality in a more chronic form of intestinal IR.
Interestingly, reduced goblet cell function and decreased mucus quality are pathophysiological phenomena in the inflamed intestine of patients with IBD.8 ,25 In addition, epithelial hypoxia is a well-known event in the colonic epithelium of patients with IBD.14 However, these phenomena have never been linked to each other. The data from this study make it temping to speculate on hypoxia as a potential aetiological factor in IBD, involved in decreased mucus quantity and quality, bacterial adherence and hence chronic inflammation. Improved understanding of hypoxia as a potential phenomenon in the aetiology or perpetuation of IBD might lead to novel preventive and/or therapeutic strategies.
In conclusion, we have shown that colonic ischaemia results in breaches in the mucus barrier and detachment of the mucus layer from the epithelium. This exposed the colonic epithelium directly to the intraluminal content which was associated with bacterial penetration. Simultaneously, goblet cell compound exocytosis was observed which was successful in clearing bacteria from the colonic crypts. In addition, increased goblet cell secretory activity was successful in restoring the mucus barrier during reperfusion, limiting the exposure of proinflammatory luminal content to the colon epithelium. We therefore propose that goblet cell compound exocytosis is an important protective mechanism involved in the prevention of bacterial translocation-induced intestinal inflammation in the human and rat colon exposed to ischaemia and reperfusion.
Acknowledgments
The authors thank Dr G C Hansson for providing the methacarn-fixation protocols, Dr T H Florin for providing the MUC2/DBA staining protocol, Dr J Bloemen and Dr J P M Derikx for their assistance with the surgical procedure, and R Janssen and H Duimel for excellent technical support.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Data supplement 1 - Online supplementary
- Data supplement 2 - Online figure legends
- Data supplement 3 - Online table 1
- Data supplement 4 - Online figure 1
- Data supplement 5 - Online figure 2
Footnotes
-
Funding This work was financially supported by the Dutch Digestive Foundation (MLDS grant WO10-57 to CHD).
-
Competing interests None.
-
Ethics approval This study was approved by the Medical Ethical Committee of Maastricht University Medical Centre and was conducted according to the revised version of the Declaration of Helsinki (October 2008, Seoul). Written informed consent was obtained from all patients. Experimental animal protocols were approved by the Animal Ethics Committee of Maastricht University Medical Centre.
-
Provenance and peer review Not commissioned; externally peer reviewed.